Resveratrol Inhibits Porcine Intestinal Glucose and Alanine Transport: Potential Roles of Na+/K+-ATPase Activity, Protein Kinase A, AMP-Activated Protein Kinase and the Association of Selected Nutrient Transport Proteins with Detergent Resistant Membranes

Abstract
:1. Introduction
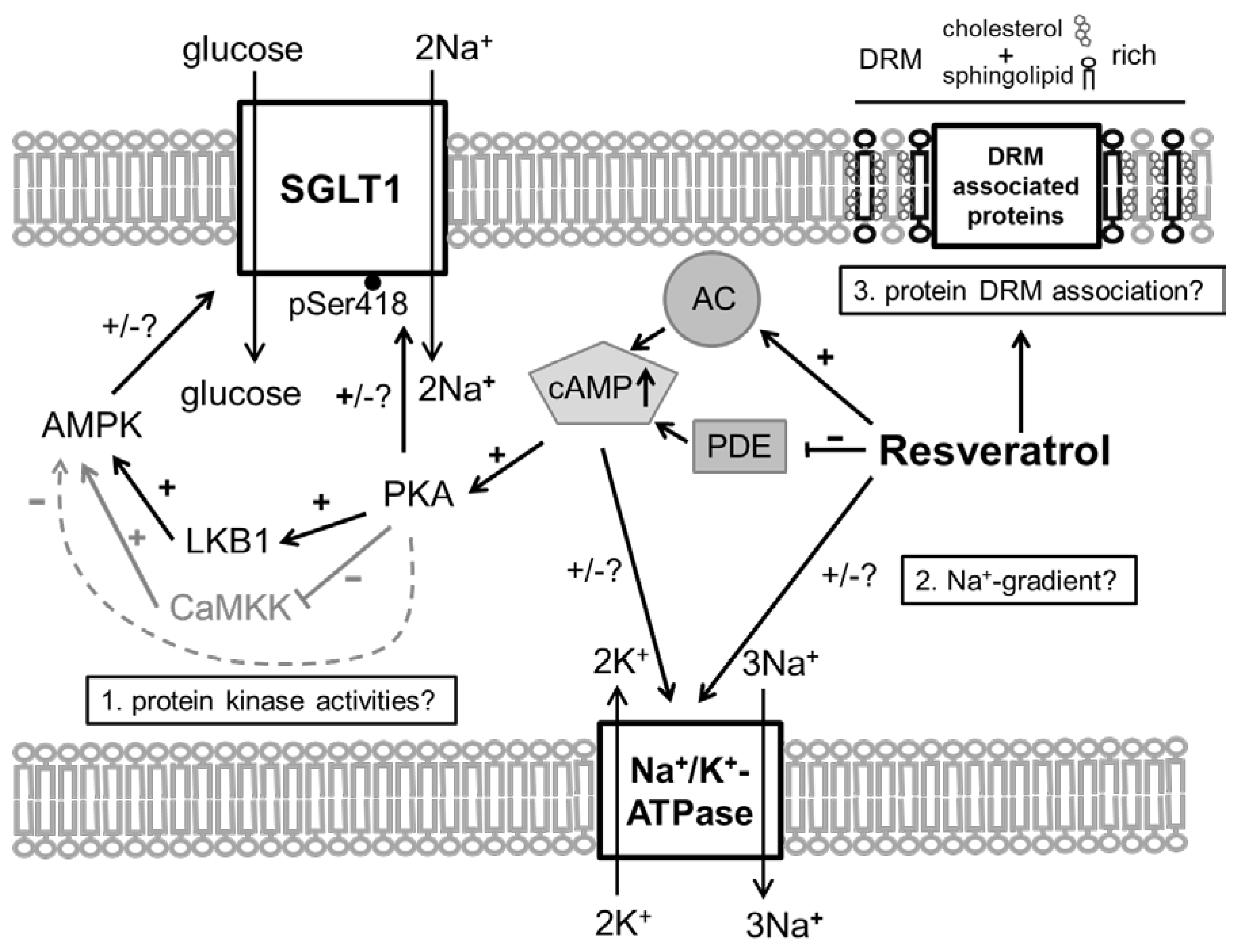
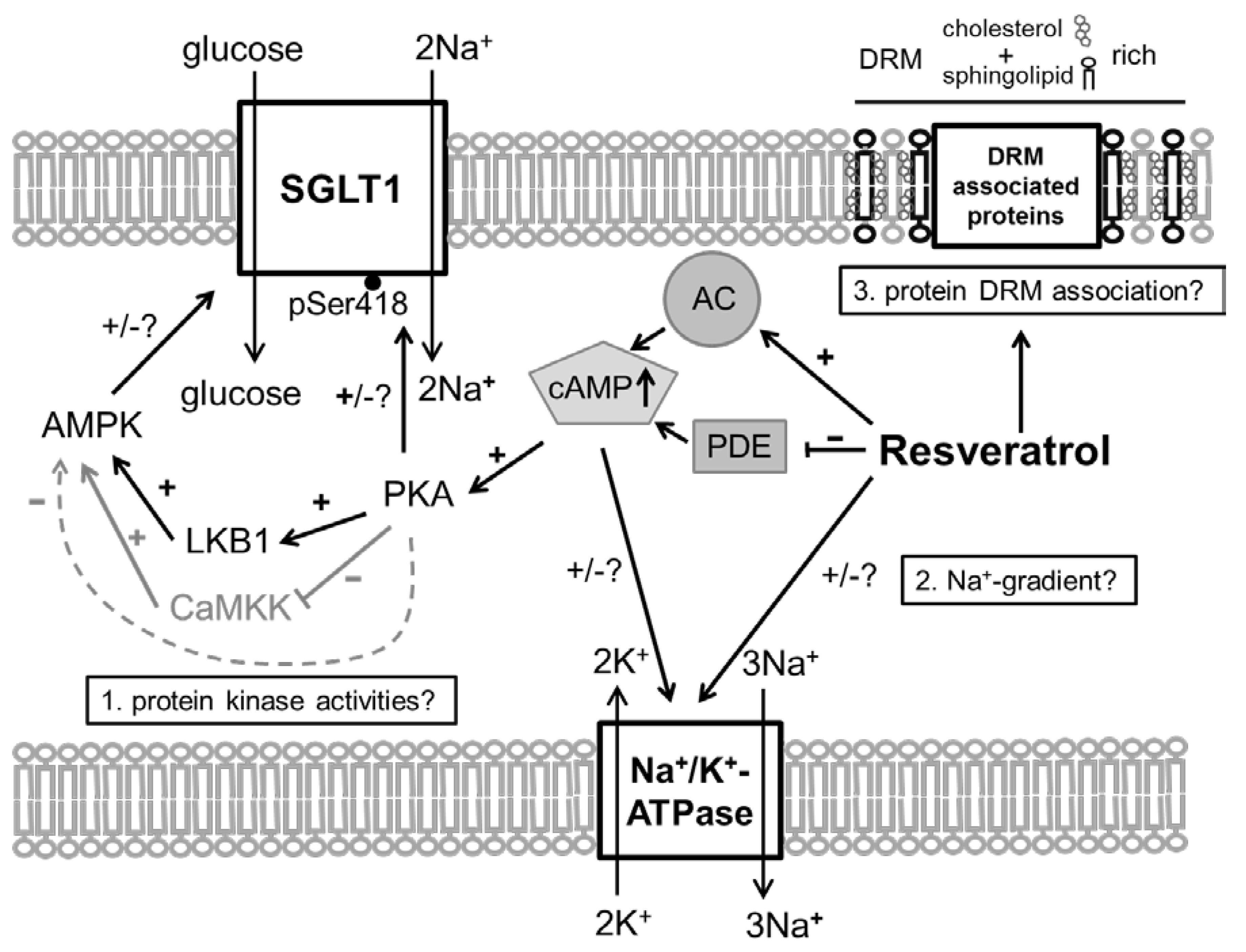
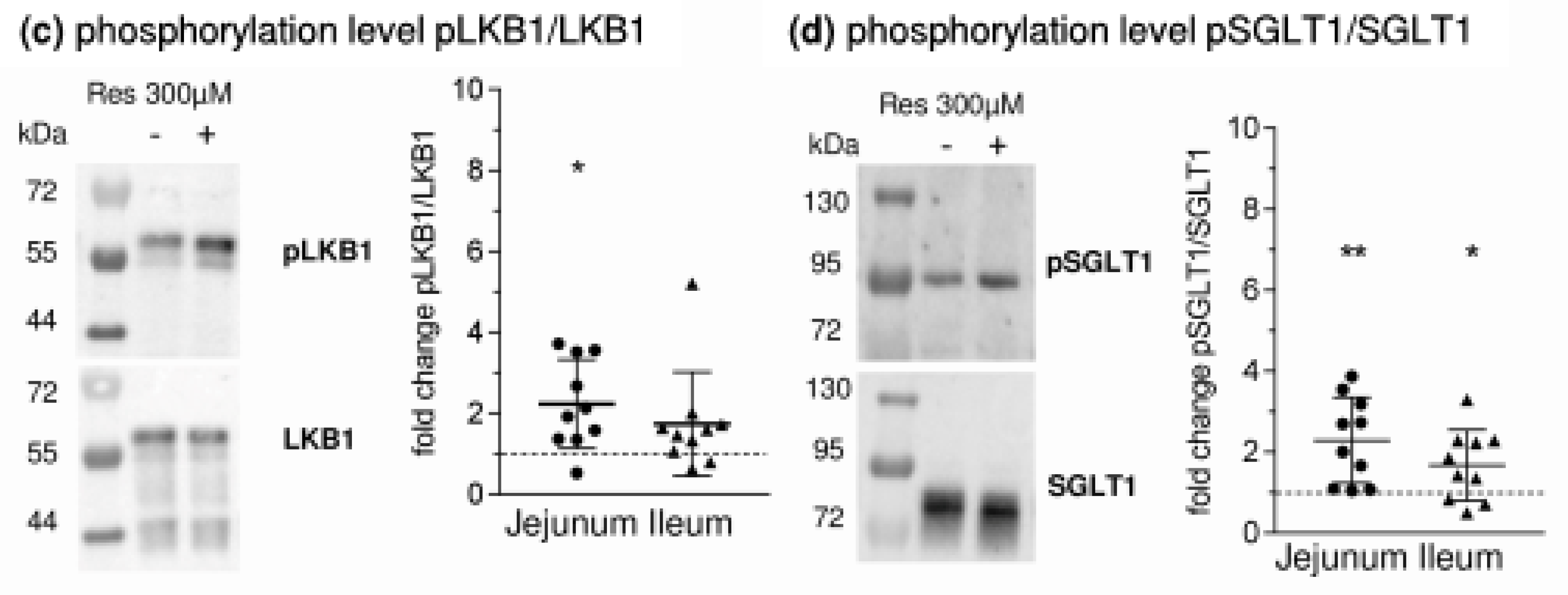
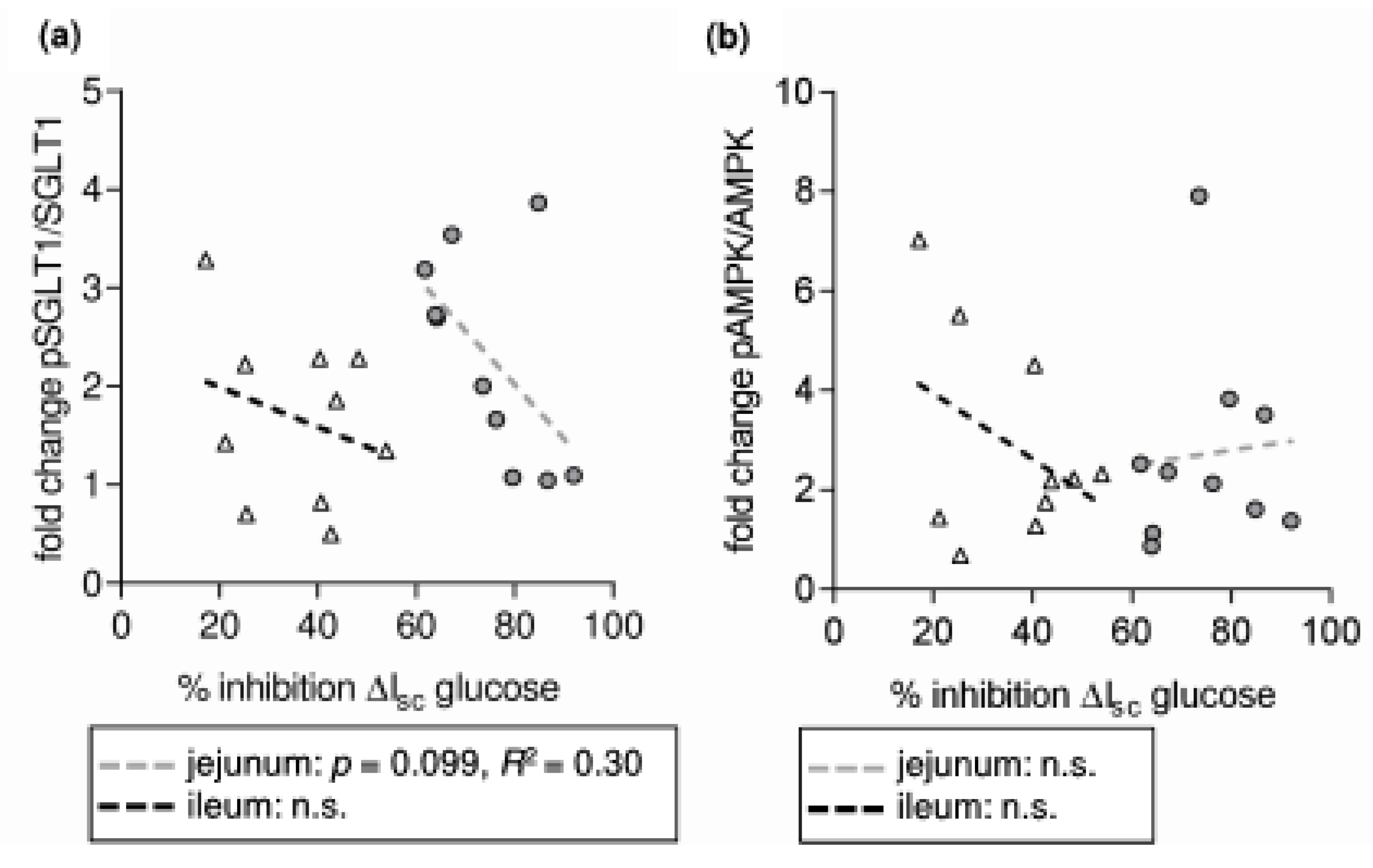
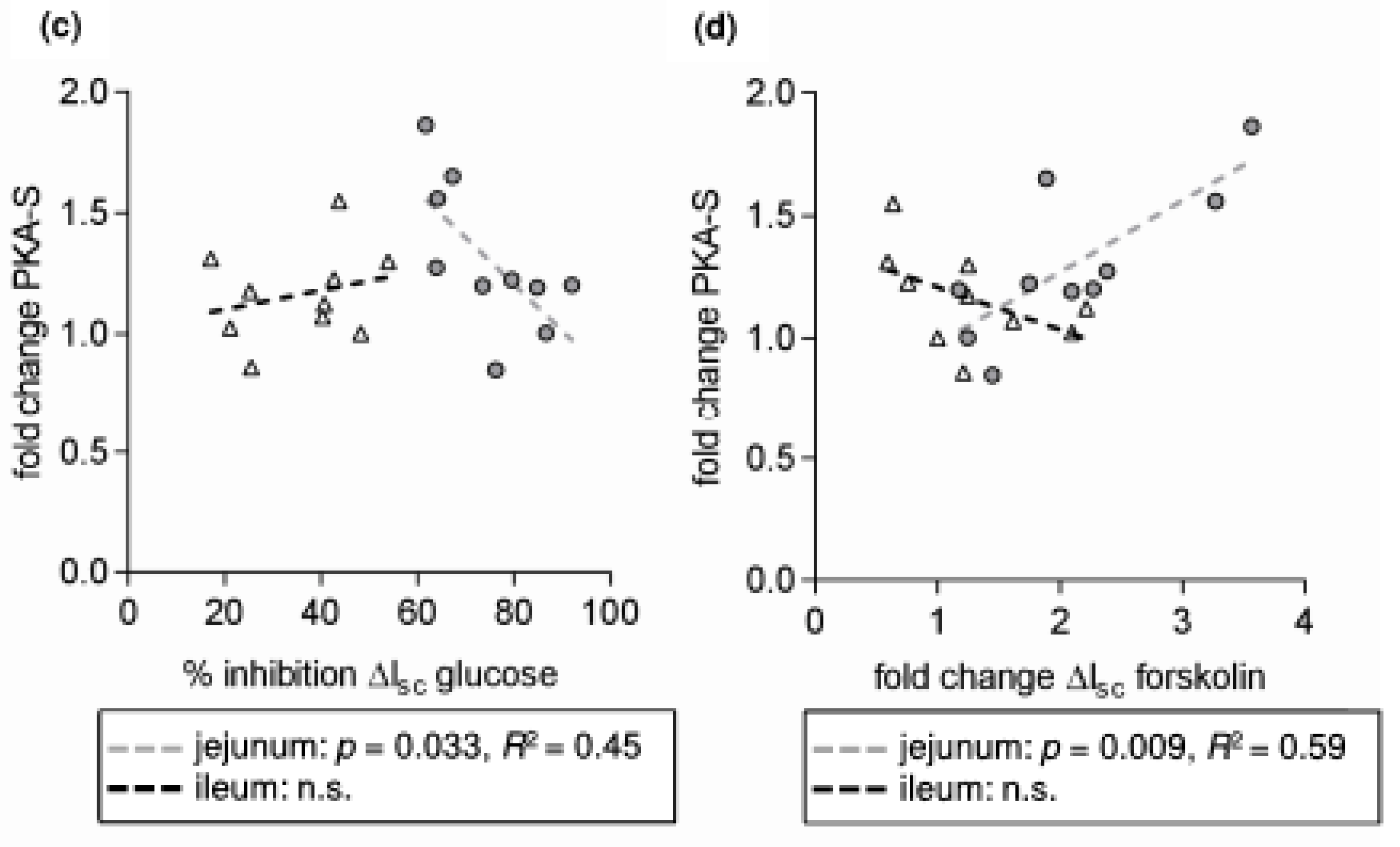
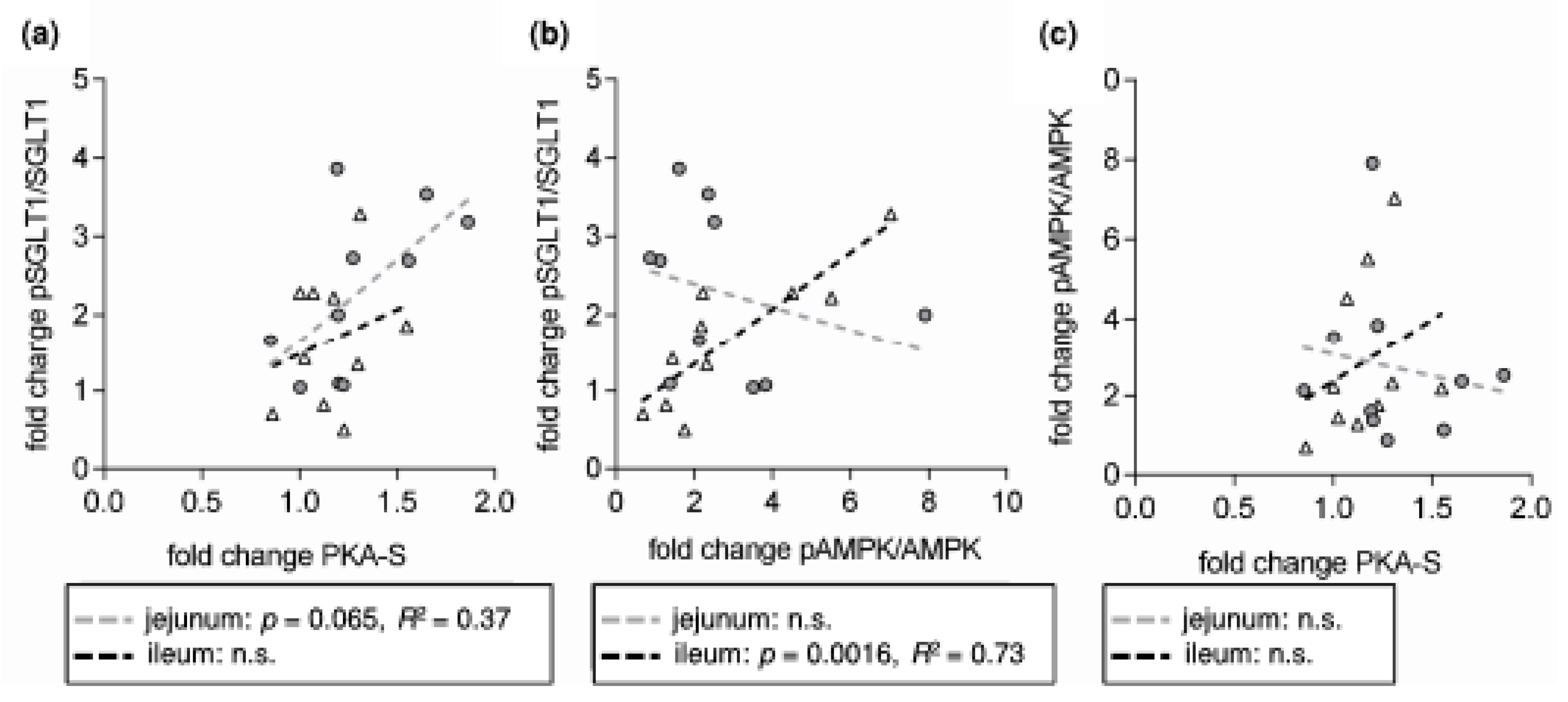
- SGLT1 activity is modulated by different intracellular pathways responsible for the functionality of enterocytes. SGLT1 is phosphorylated by protein kinase A (PKA) at Serin 418 (Ser418), which is likely to activate SGLT1, although, depending on the species, inhibitory effects have also been described [9,10]. AMP-activated protein kinase (AMPK) also influences SGLT1. Inhibitory effects [11] were observed as activating responses as well [12,13].
- The secondary active transport of glucose across the apical membrane depends on the inwardly directed Na+ gradient that is maintained by the basolateral Na+/K+-ATPase. Additionally, membrane potential affects Na+-coupled sugar transport and the activity of SGLT1 [14,15]. The local Na+-evoked, SGLT1-inhibiting depolarization at the apical membrane is compensated by potassium (K+) channels localized in the basolateral or apical enterocyte membrane, depending on paracellular tightness [16,17].
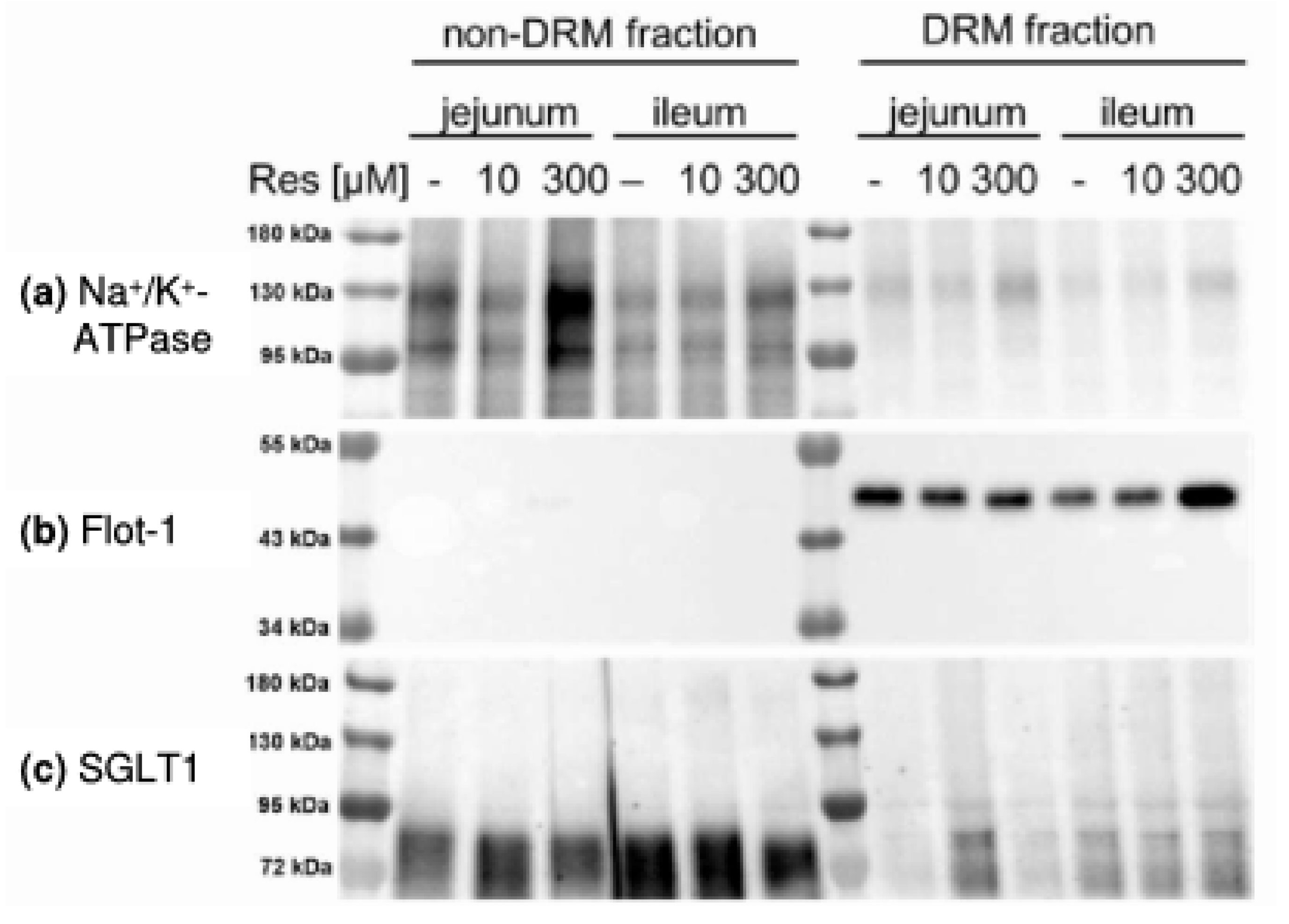
- The activity of several transmembrane proteins depends on their lipid environment including the membrane fluidity and their association with detergent-resistant membrane (DRM) fractions (or lipid rafts) that represent membrane fractions enriched in cholesterol and sphingolipids. This is also true for the activity of SGLT1 that increases with decreasing membrane fluidity [18] and decreases with decreasing membrane cholesterol content of the membrane [18,19,20].
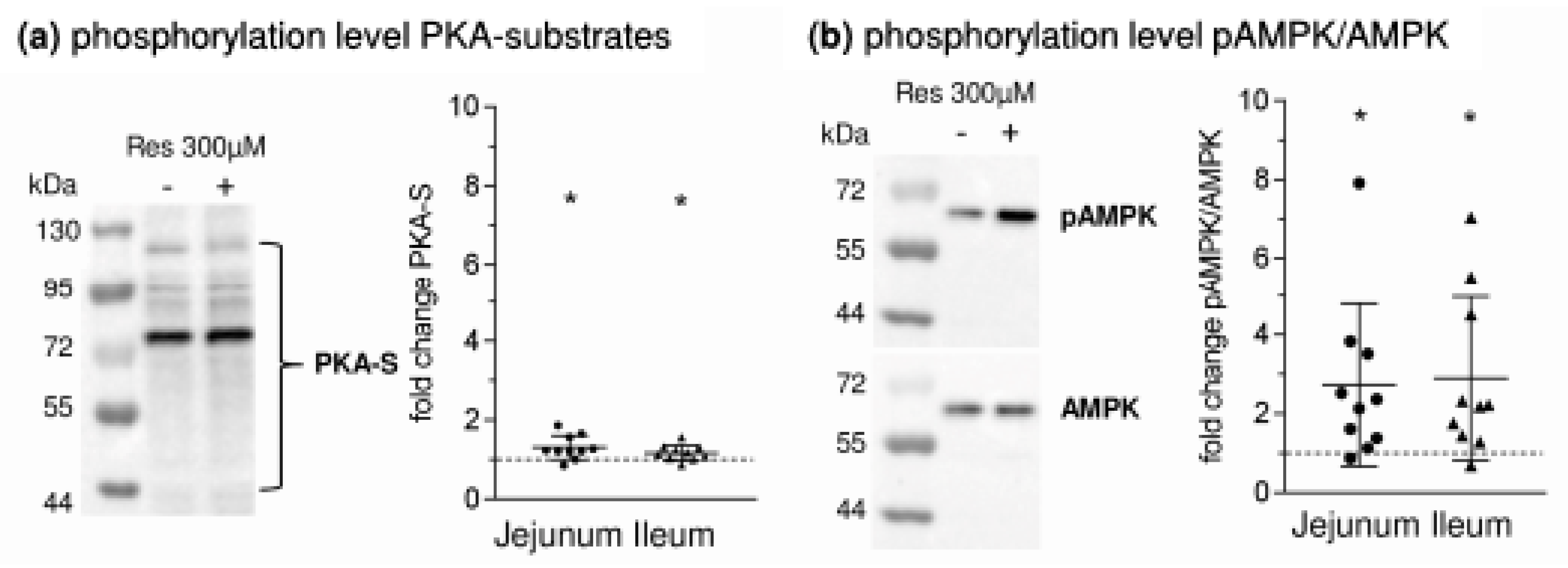
- RSV acts on intracellular pathways. In this context, the inhibition of phosphodiesterases (PDE) [21], or also adenylate cyclase activation [22,23], leading to increased levels of cyclic adenosine monophosphate (cAMP), is of interest as the main stimulus for PKA activation. Additionally, RSV is known to activate AMPK [21]. Besides mechanisms related to cellular energy status, AMPK is activated by phosphorylation at threonine 172 (Thr172) by upstream kinases including liver kinase B1 (LKB1) [24]. In turn, it has been shown that activating effects of RSV on AMPK require LKB1 and that RSV activates LKB1 [25,26]. The role of PKA in regulating AMPK is complex since the possibility of activating LKB1 and therefore promoting AMPK activity has been described [27] as well as the potential to inhibit AMPK by phosphorylation or by inhibiting the activating AMPK upstream kinase calcium/calmodulin-dependent protein kinase kinase (CaMKK) [28,29].
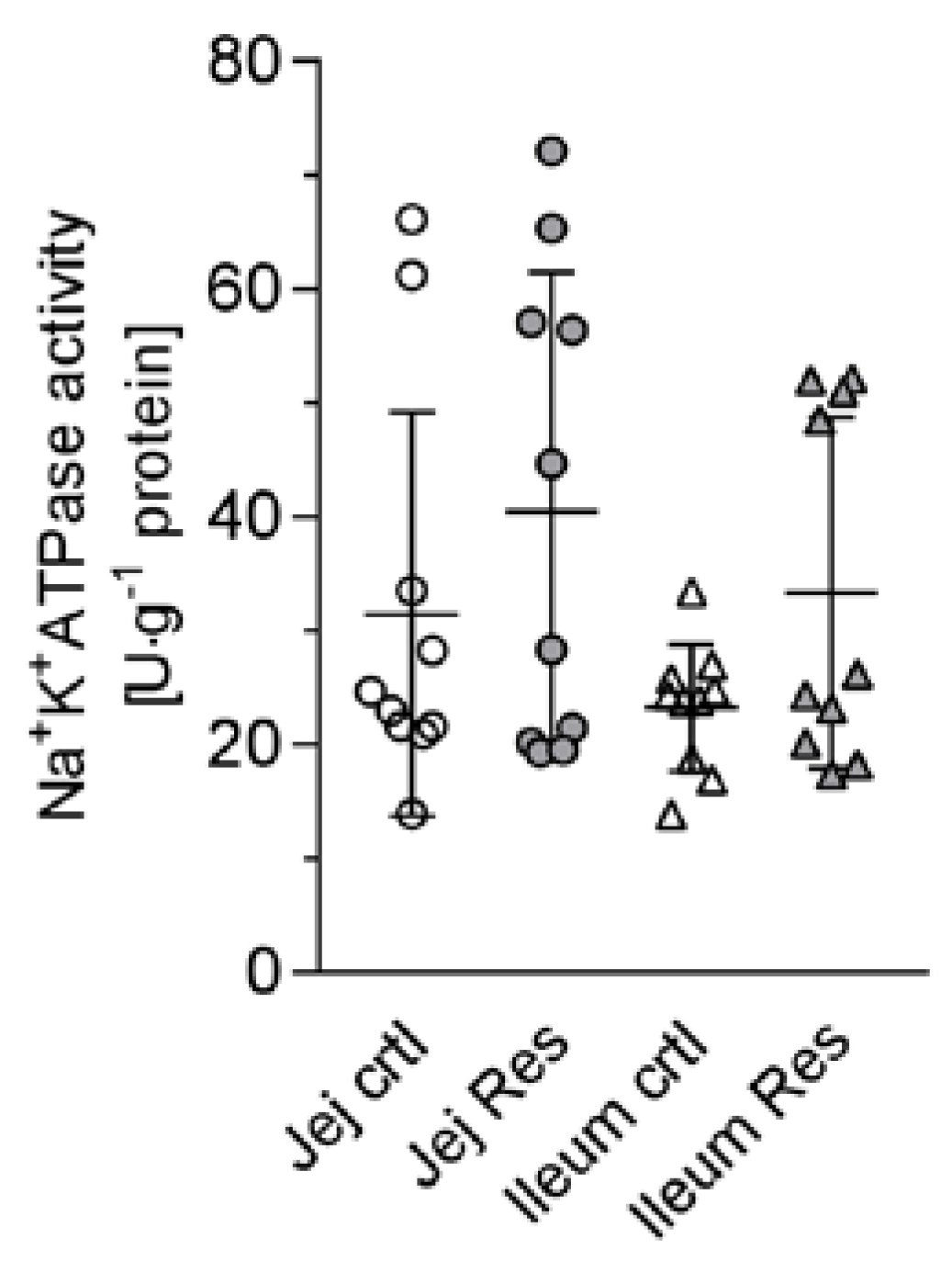
- In addition to potential effects of resveratrol on the membrane potential by influencing the K+ conductance, RSV may also exert direct effects on the activity of the Na+/K+-ATPase. In erythrocytes, RSV activates the pump [30], but assuming a cAMP-mediated effect it has to be taken into account that the effects of cAMP on the activity of Na+/K+-ATPase are tissue selective [31].
- RSV is known to affect membrane properties by accumulating in the outer leaflet of the lipid bilayer [32] or in DRMs and one way to enter the cell is via raft-dependent endocytosis [33,34]. There is also evidence that RSV influences the formation of lipid rafts [35], which might be associated with the variety of effects that are exhibited by RSV.
- may also lead to an inhibition of other intestinal Na+-dependent transport processes such as Na+-dependent alanine transport and whether the activity of the Na+/K+-ATPase is altered;
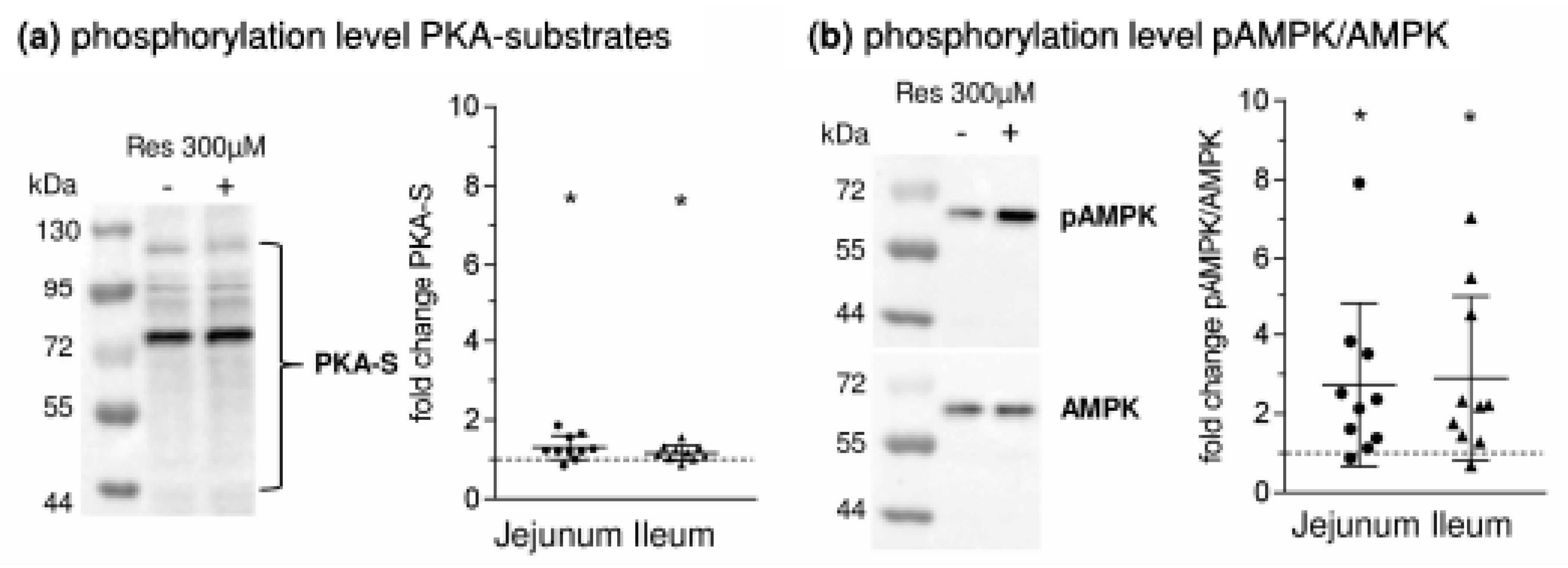
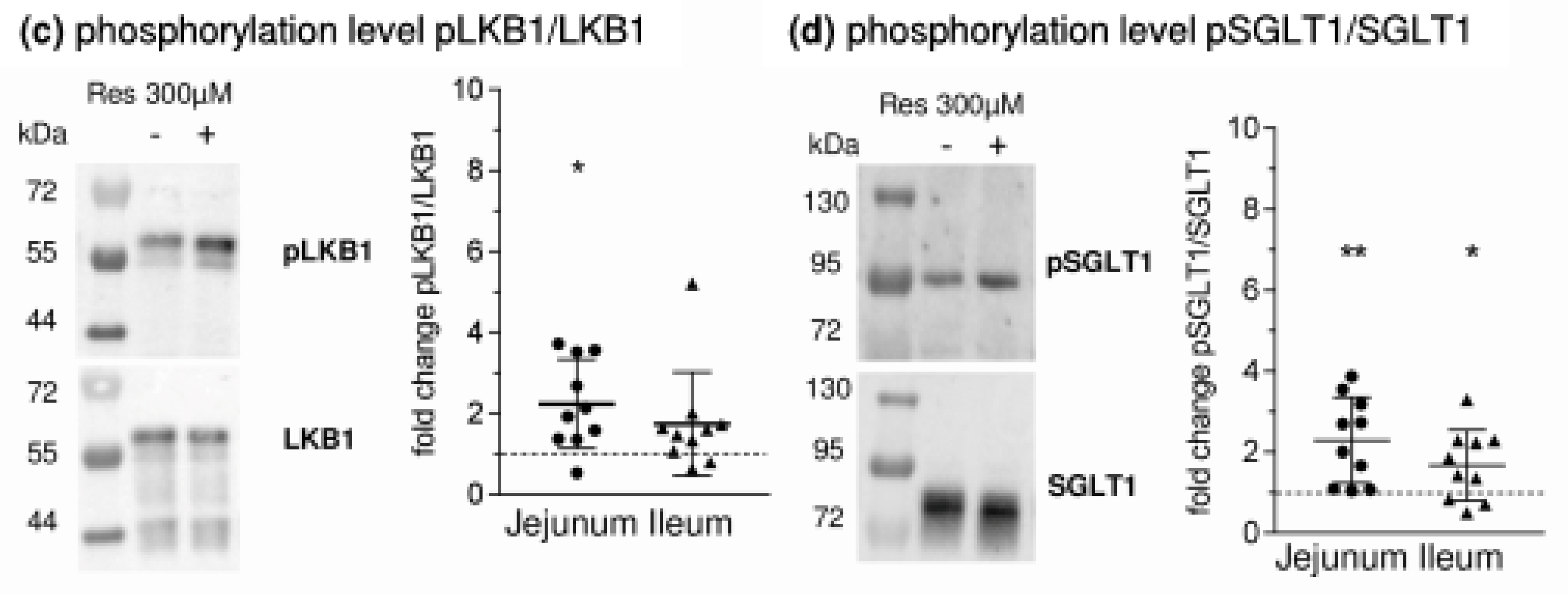
- leads to an activation of AMPK, PKA and LKB1 and to changes in the apical expression and phosphorylation of SGLT1 at the PKA phosphorylation site Ser418;
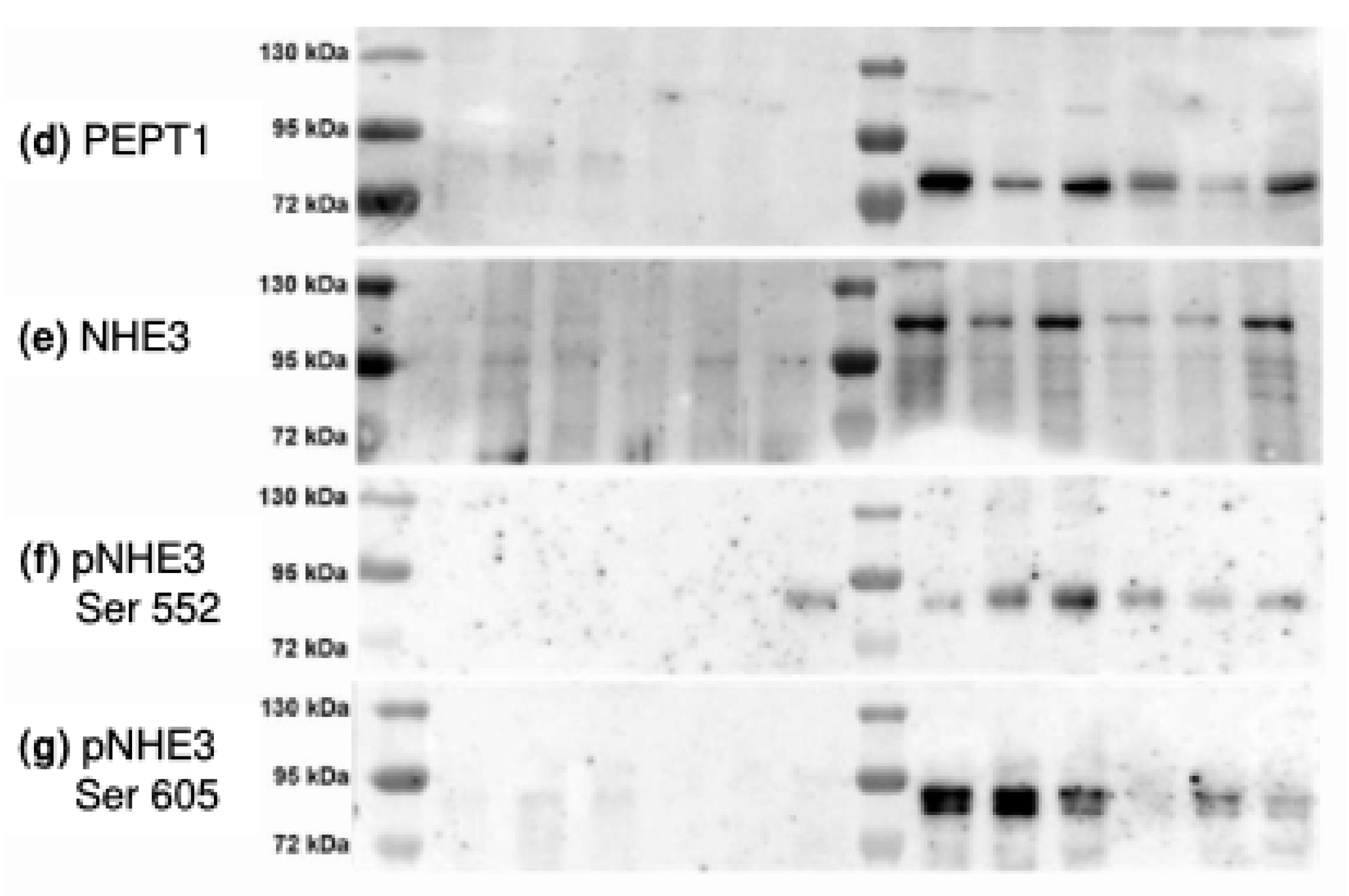
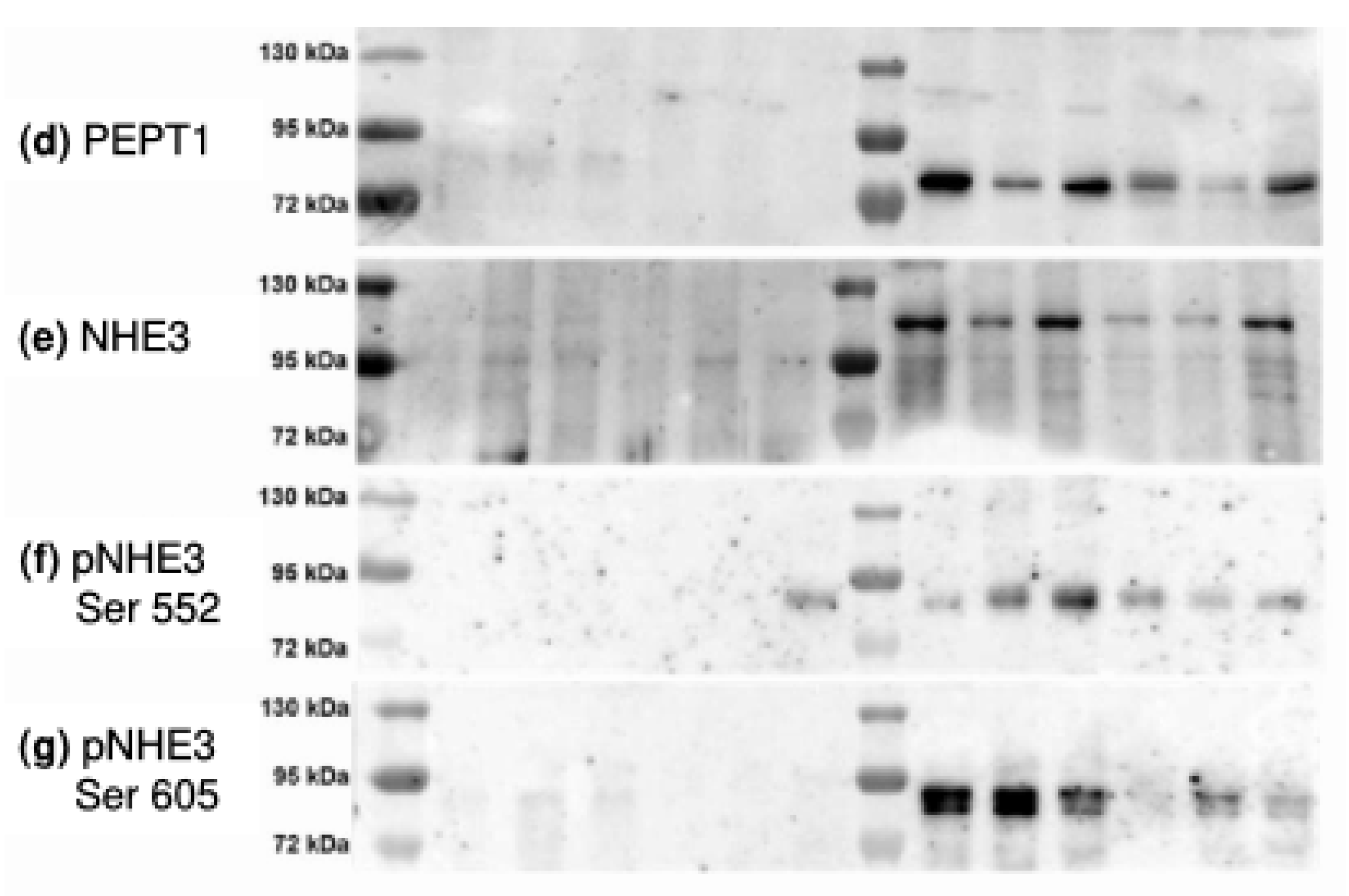
- changes the distribution of SGLT1 and other putatively DRM-associated transport proteins (Na+/H+-exchanger 3 (NHE3) and its phosphorylated forms and PEPT1) between DRM and non-DRM fractions in the apical membrane of enterocytes.
2. Materials and Methods
2.1. Animals and Tissue Removal
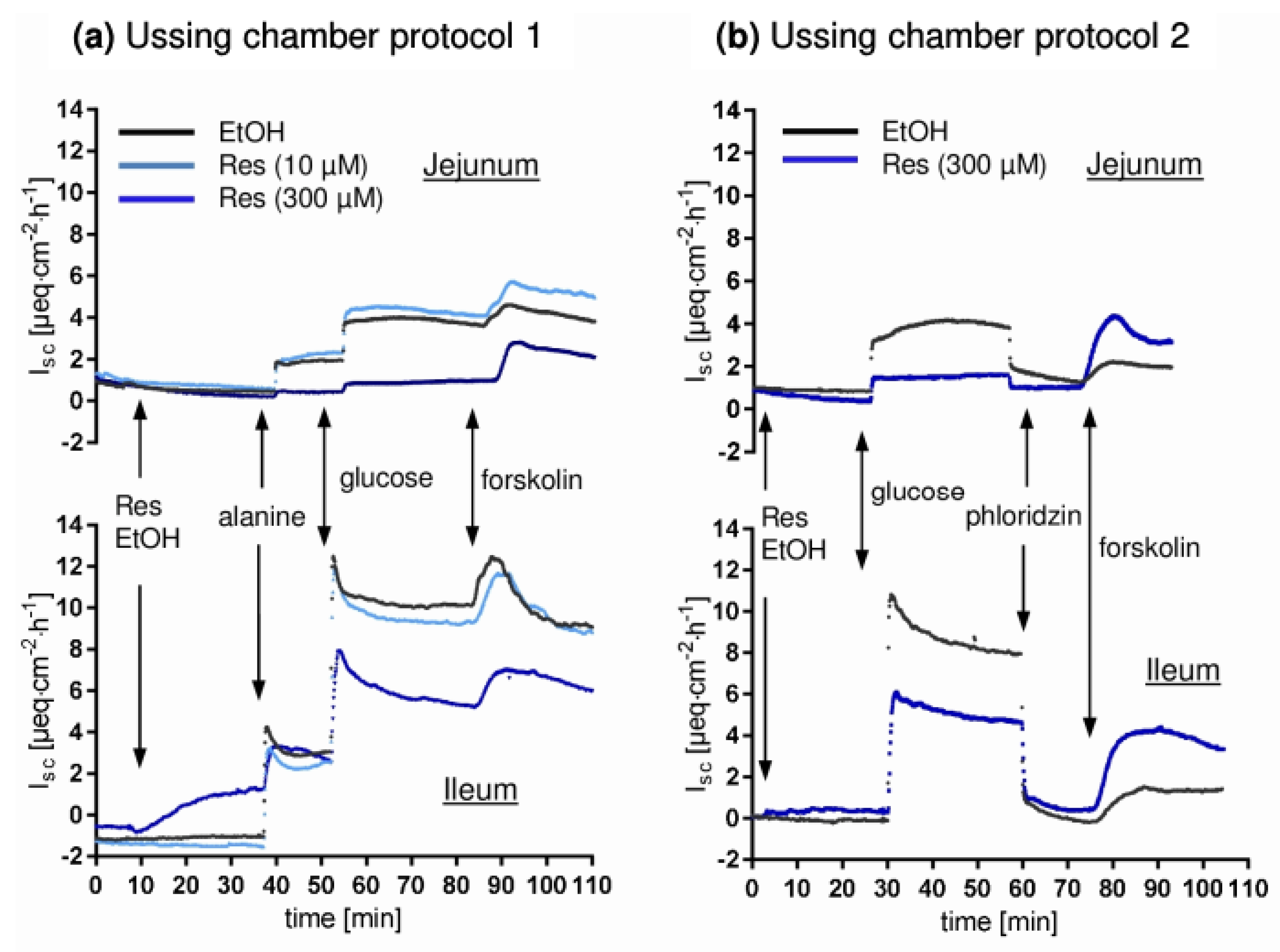
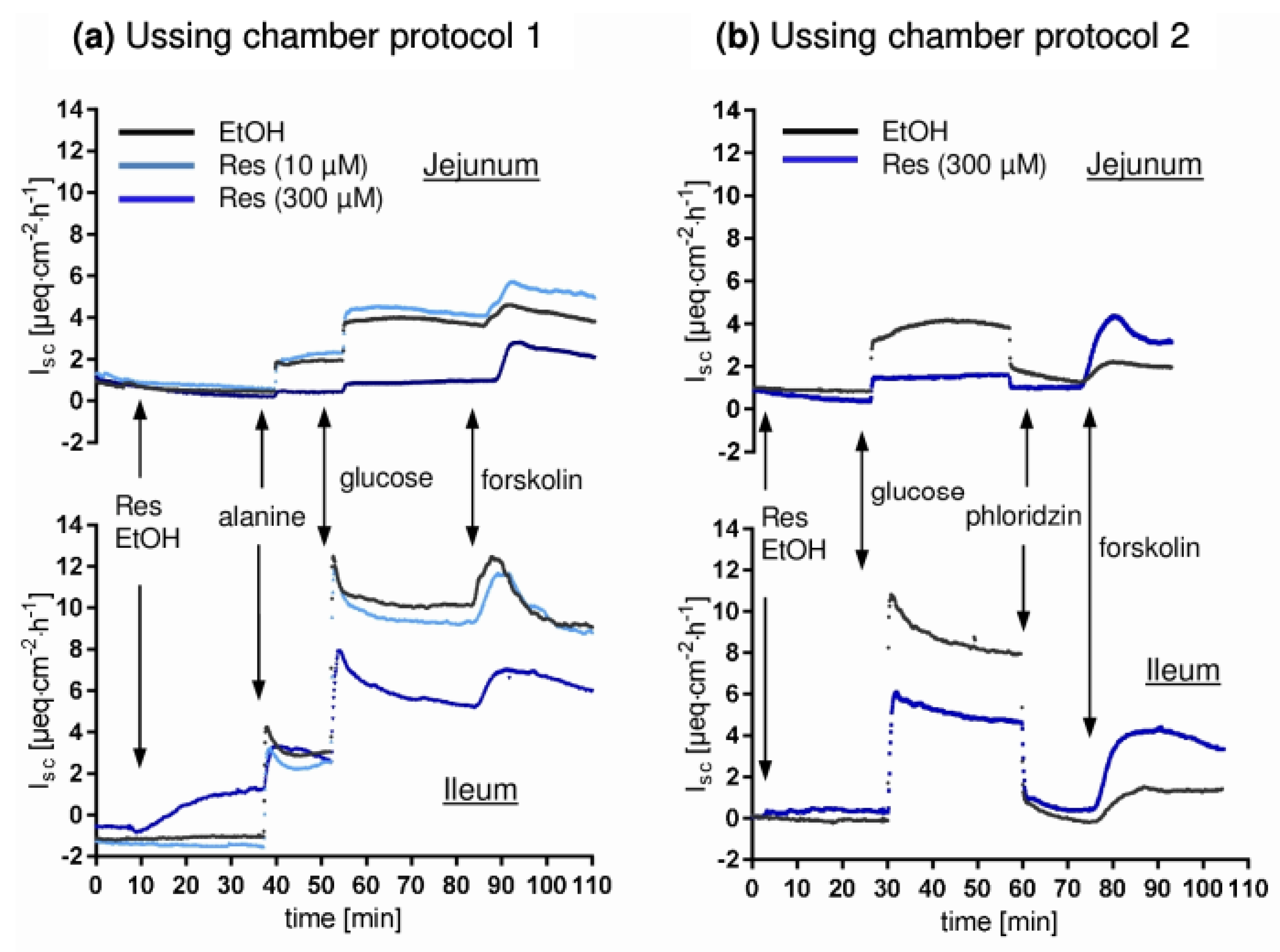
2.2. Ussing Chamber Experiments
2.3. Tissue Incubations for Expression Analysis
2.4. Tissue Preparations
2.5. Western Blot
2.6. Data Analysis and Statistics
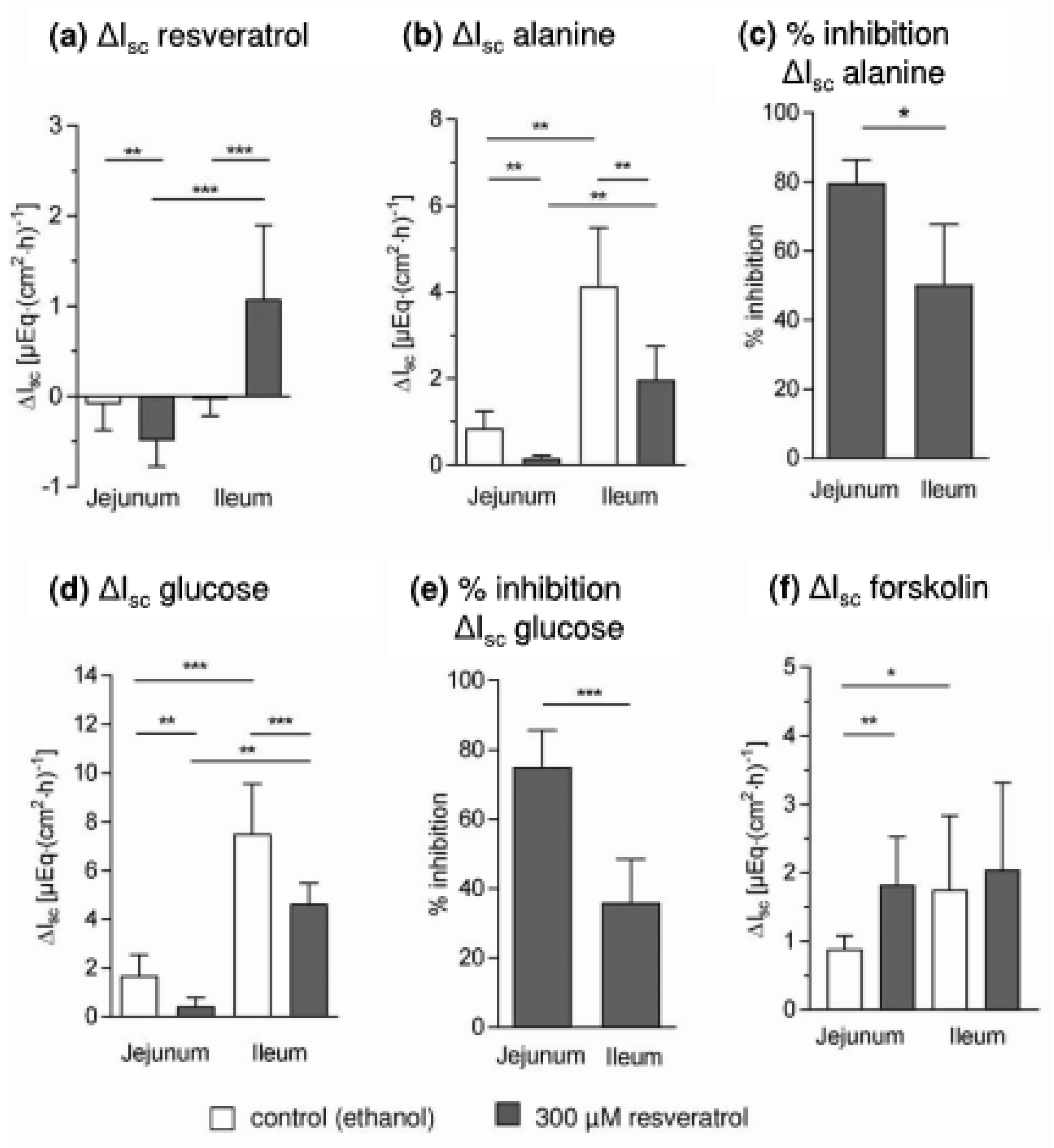
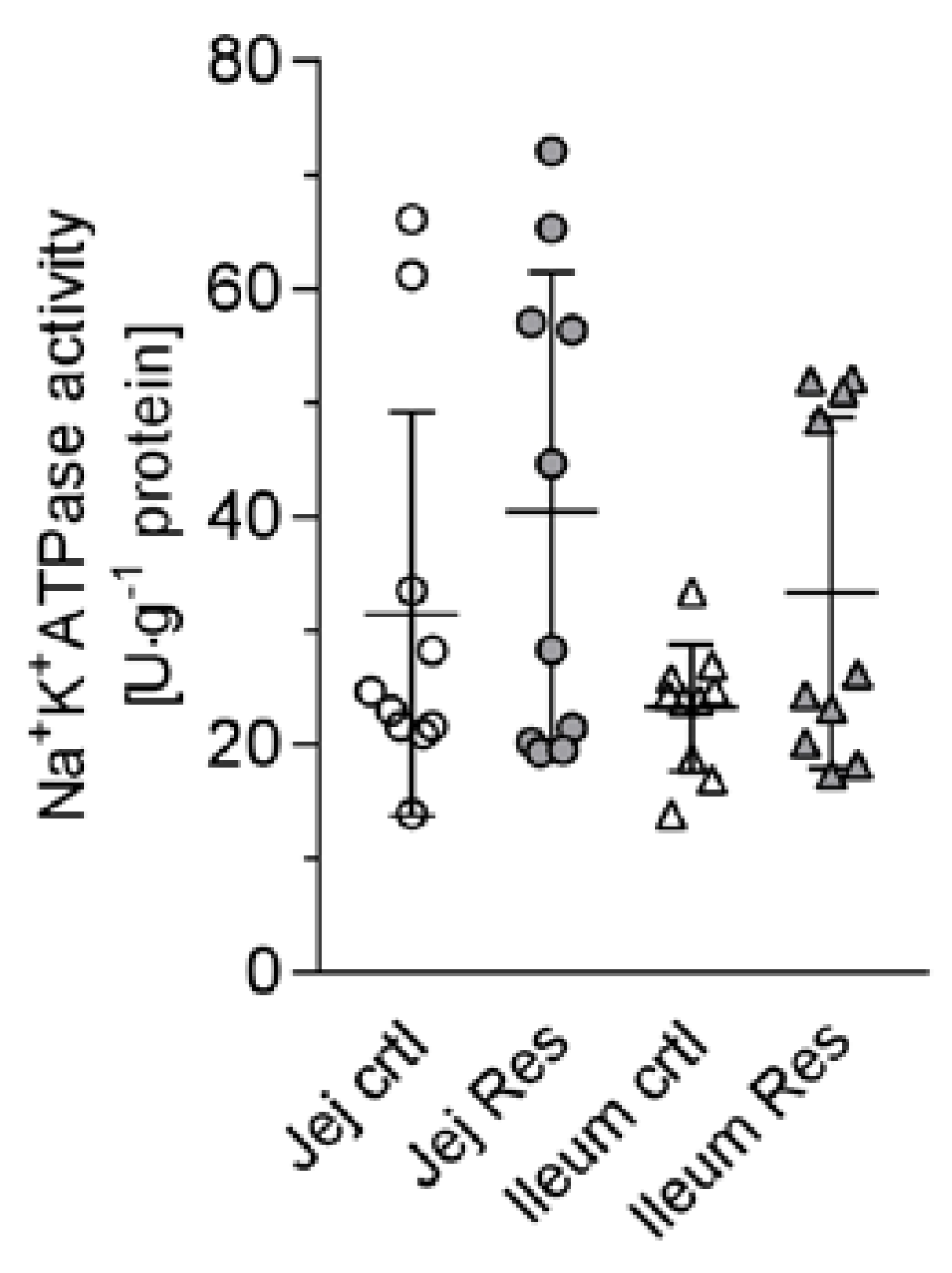
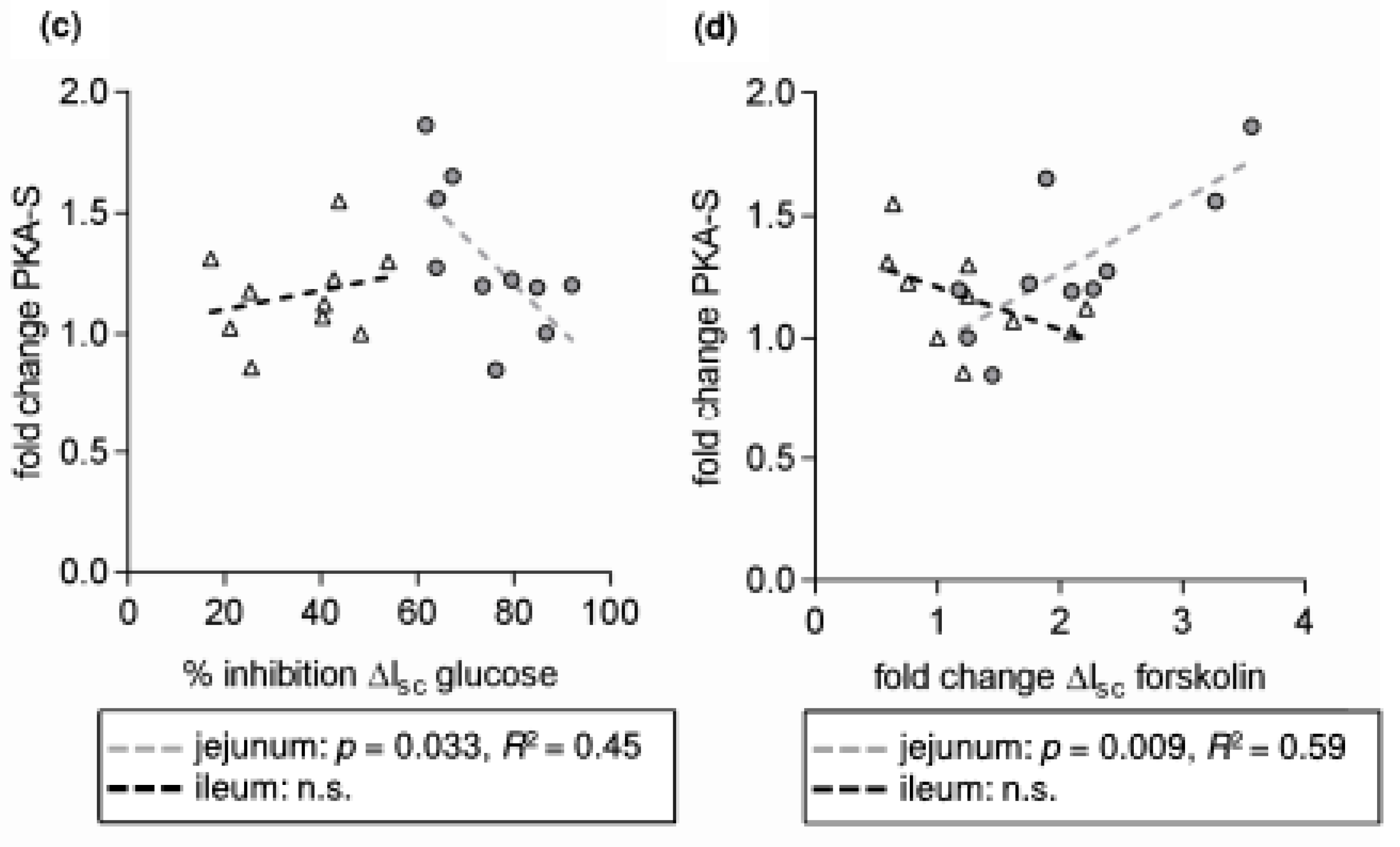
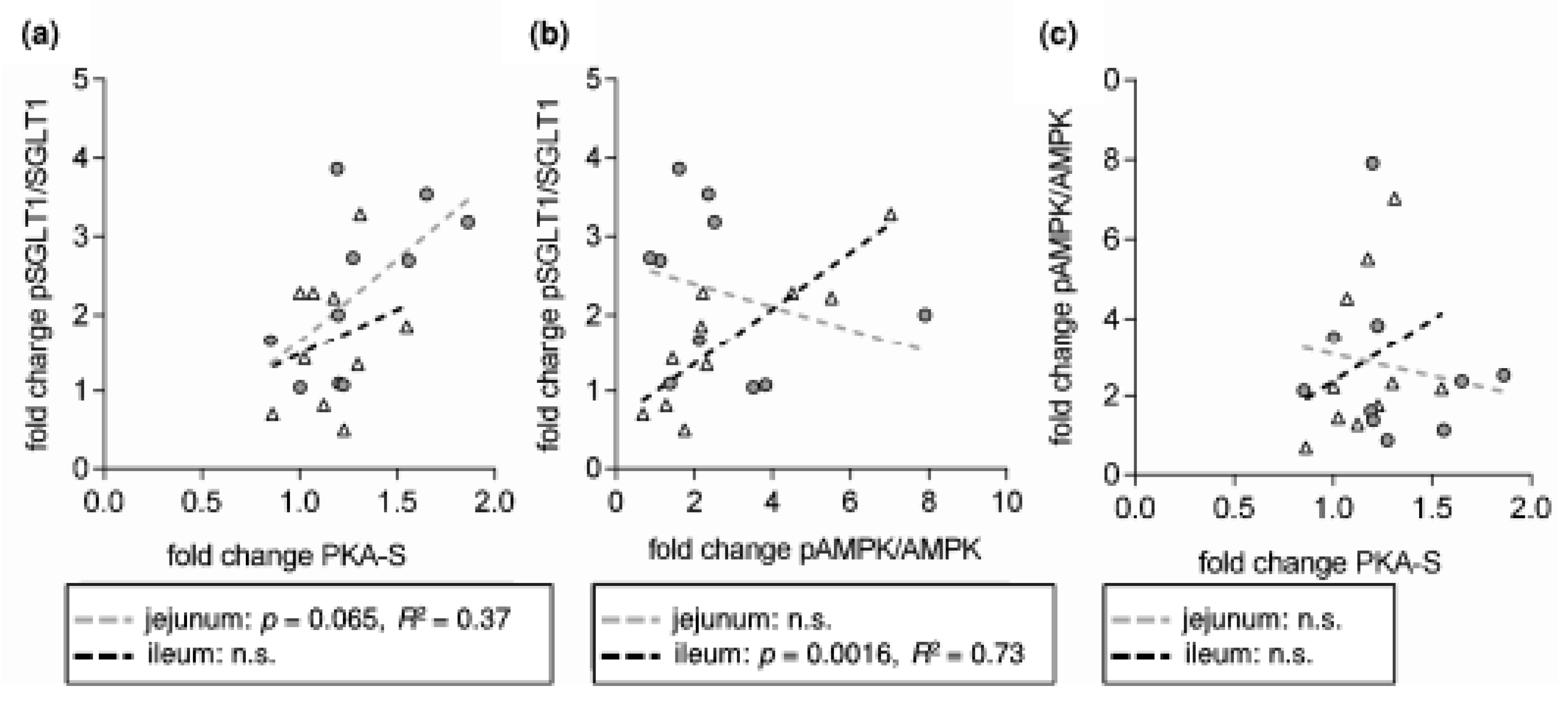
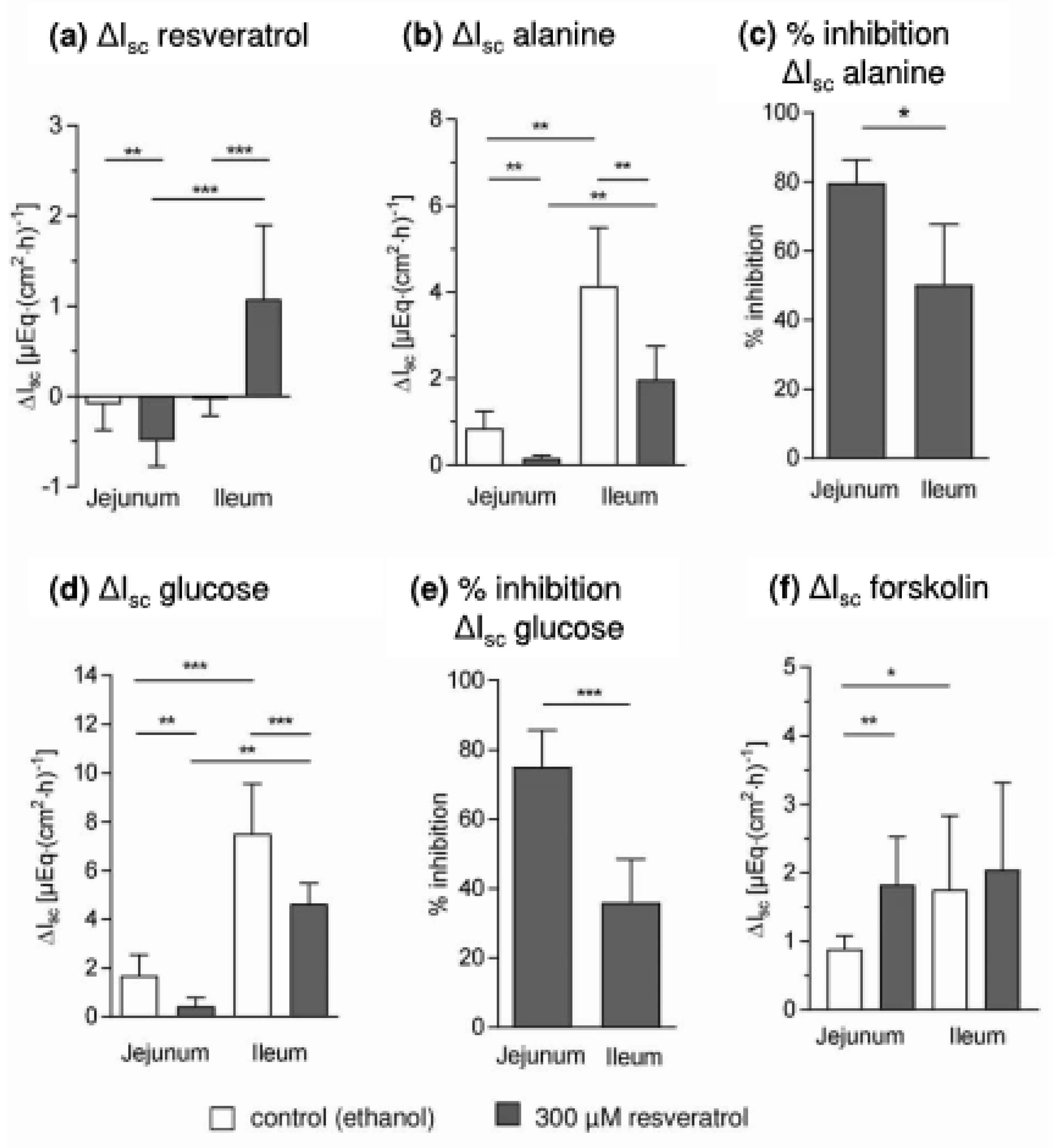
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Delmas, D.; Aires, V.; Limagne, E.; Dutartre, P.; Mazue, F.; Ghiringhelli, F.; Latruffe, N. Transport, stability, and biological activity of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Chung, H.J.; Lee, H.K.; D’Souza, R.; Jeon, Y.; Kim, H.J.; Kweon, S.J.; Hong, S.T. Treatment of obesity with the resveratrol-enriched rice DJ-526. Sci. Rep. 2014, 4, 3879. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Shin, W.C.; Ryu, H.S.; Lee, D.W.; Moon, E.; Seo, C.S.; Hwang, E.; Lee, H.S.; Ahn, M.H.; Jeon, Y.; et al. Creation of resveratrol-enriched rice for the treatment of metabolic syndrome and related diseases. PLoS ONE 2013, 8, e57930. [Google Scholar] [CrossRef] [PubMed]
- 4Blumenstein, I.; Keseru, B.; Wolter, F.; Stein, J. The chemopreventive agent resveratrol stimulates cyclic AMP-dependent chloride secretion in vitro. Clin. Cancer Res. 2005, 11, 5651–5656. [Google Scholar] [CrossRef] [PubMed]
- Geillinger, K.E.; Kipp, A.P.; Schink, K.; Roder, P.V.; Spanier, B.; Daniel, H. Nrf2 regulates the expression of the peptide transporter PEPT1 in the human colon carcinoma cell line Caco-2. Biochim. Biophys. Acta 2014, 1840, 1747–1754. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Liu, Z.; Huo, X.; Wang, C.; Meng, Q.; Liu, Q.; Sun, H.; Sun, P.; Yang, X.; Shu, X.; et al. Enhancement effect of resveratrol on the intestinal absorption of bestatin by regulating PEPT1, MDR1 and MRP2 in vivo and in vitro. Int. J. Pharm. 2015, 495, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Guschlbauer, M.; Klinger, S.; Burmester, M.; Horn, J.; Kulling, S.E.; Breves, G. Trans-Resveratrol and epsilon-viniferin decrease glucose absorption in porcine jejunum and ileum in vitro. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 165, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.M.; Loo, D.D.; Hirayama, B.A. Biology of human sodium glucose transporters. Physiol. Rev. 2011, 91, 733–794. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, J.R.; Loo, D.D.; Wright, E.M. Regulation of Na+/glucose cotransporter expression by protein kinases in Xenopus laevis oocytes. J. Biol. Chem. 1996, 271, 14740–14746. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, Y.; Eguchi, T.; Ishida, H. Mechanism of beta-adrenergic agonist-induced transmural transport of glucose in rat small intestine. Regulation of phosphorylation of SGLT1 controls the function. Biochim. Biophys. Acta 1997, 1357, 306–318. [Google Scholar] [CrossRef]
- Sakar, Y.; Meddah, B.; Faouzi, M.A.; Cherrah, Y.; Bado, A.; Ducroc, R. Metformin-induced regulation of the intestinal D-glucose transporters. J. Physiol. Pharmacol. 2010, 61, 301–307. [Google Scholar] [PubMed]
- Lang, F.; Foller, M. Regulation of ion channels and transporters by AMP-activated kinase (AMPK). Channels 2013, 8, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Sopjani, M.; Bhavsar, S.K.; Fraser, S.; Kemp, B.E.; Foller, M.; Lang, F. Regulation of Na+-coupled glucose carrier SGLT1 by AMP-activated protein kinase. Mol. Membr. Biol. 2010, 27, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Umbach, J.A.; Coady, M.J.; Wright, E.M. Intestinal Na+/glucose cotransporter expressed in Xenopus oocytes is electrogenic. Biophys. J. 1990, 57, 1217–1224. [Google Scholar] [CrossRef]
- Lapointe, J.Y.; Hudson, R.L.; Schultz, S.G. Current-voltage relations of sodium-coupled sugar transport across the apical membrane of Necturus small intestine. J. Membr. Biol. 1986, 93, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Heitzmann, D.; Warth, R. Physiology and pathophysiology of potassium channels in gastrointestinal epithelia. Physiol. Rev. 2008, 88, 1119–1182. [Google Scholar] [CrossRef] [PubMed]
- Warth, R. Potassium channels in epithelial transport. Pflugers Arch. 2003, 446, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Murtazina, R.; Kovbasnjuk, O.; Donowitz, M.; Li, X. Na+/H+ exchanger NHE3 activity and trafficking are lipid Raft-dependent. J. Biol. Chem. 2006, 281, 17845–17855. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Kim, M.O.; Ryu, J.M.; Han, H.J. Regulation of SGLT expression and localization through Epac/PKA-dependent caveolin-1 and F-actin activation in renal proximal tubule cells. Biochim. Biophys. Acta 2012, 1823, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Runembert, I.; Queffeulou, G.; Federici, P.; Vrtovsnik, F.; Colucci-Guyon, E.; Babinet, C.; Briand, P.; Trugnan, G.; Friedlander, G.; Terzi, F. Vimentin affects localization and activity of sodium-glucose cotransporter SGLT1 in membrane rafts. J. Cell Sci. 2002, 115, 713–724. [Google Scholar] [PubMed]
- Park, S.J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.; Ke, H.; Rehmann, H.; Taussig, R.; Brown, A.L.; et al. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell 2012, 148, 421–433. [Google Scholar] [CrossRef] [PubMed]
- El-Mowafy, A.M.; Alkhalaf, M. Resveratrol activates adenylyl-cyclase in human breast cancer cells: A novel, estrogen receptor-independent cytostatic mechanism. Carcinogenesis 2003, 24, 869–873. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, M.L.; Zhou, Y.; Yi, L.; Gao, Y.X.; Ran, L.; Chen, S.H.; Zhang, T.; Zhou, X.; Zou, D.; et al. Resveratrol improves hepatic steatosis by inducing autophagy through the cAMP signaling pathway. Mol. Nutr. Food Res. 2015, 59, 1443–1457. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.C.; Zierath, J.R. AMP-activated protein kinase signaling in metabolic regulation. J. Clin. Investig. 2006, 116, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Lan, F.; Weikel, K.A.; Cacicedo, J.M.; Ido, Y. Resveratrol-Induced AMP-Activated Protein Kinase Activation Is Cell-Type Dependent: Lessons from Basic Research for Clinical Application. Nutrients 2017, 9, 751. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Liu, B. Resveratrol via activation of LKB1-AMPK signaling suppresses oxidative stress to prevent endothelial dysfunction in diabetic mice. Clin. Exp. Hypertens. 2016, 38, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.P.; Reoma, J.L.; Gamm, D.M.; Uhler, M.D. LKB1, a novel serine/threonine protein kinase and potential tumour suppressor, is phosphorylated by cAMP-dependent protein kinase (PKA) and prenylated in vivo. Biochem. J. 2000, 345, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Djouder, N.; Tuerk, R.D.; Suter, M.; Salvioni, P.; Thali, R.F.; Scholz, R.; Vaahtomeri, K.; Auchli, Y.; Rechsteiner, H.; Brunisholz, R.A.; et al. PKA phosphorylates and inactivates AMPKalpha to promote efficient lipolysis. EMBO J. 2010, 29, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Hurley, R.L.; Barre, L.K.; Wood, S.D.; Anderson, K.A.; Kemp, B.E.; Means, A.R.; Witters, L.A. Regulation of AMP-activated protein kinase by multisite phosphorylation in response to agents that elevate cellular cAMP. J. Biol. Chem. 2006, 281, 36662–36672. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Role of resveratrol in regulation of membrane transporters and integrity of human erythrocytes. Biochem. Biophys. Res. Commun. 2014, 453, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Bertorello, A.M.; Katz, A.I. Short-term regulation of renal Na-K-ATPase activity: Physiological relevance and cellular mechanisms. Am. J. Physiol. 1993, 265, F743–F755. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, S.; Mohan, A.; Narayanan, S.; Sethuraman, S.; Krishnan, U.M. Dose-dependent interaction of trans-resveratrol with biomembranes: Effects on antioxidant property. J. Med. Chem. 2013, 56, 970–981. [Google Scholar] [CrossRef] [PubMed]
- Delmas, D.; Aires, V.; Colin, D.J.; Limagne, E.; Scagliarini, A.; Cotte, A.K.; Ghiringhelli, F. Importance of lipid microdomains, rafts, in absorption, delivery, and biological effects of resveratrol. Ann. N. Y. Acad. Sci. 2013, 1290, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Colin, D.; Limagne, E.; Jeanningros, S.; Jacquel, A.; Lizard, G.; Athias, A.; Gambert, P.; Hichami, A.; Latruffe, N.; Solary, E.; et al. Endocytosis of resveratrol via lipid rafts and activation of downstream signaling pathways in cancer cells. Cancer Prev. Res. (Phila.) 2011, 4, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Nunes, C.; Reis, S. Resveratrol induces ordered domains formation in biomembranes: Implication for its pleiotropic action. Biochim. Biophys. Acta 2016, 1858, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Varum, F.J.; Veiga, F.; Sousa, J.S.; Basit, A.W. Mucus thickness in the gastrointestinal tract of laboratory animals. J. Pharm. Pharmacol. 2012, 64, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Nejdfors, P.; Ekelund, M.; Jeppsson, B.; Westrom, B.R. Mucosal in vitro permeability in the intestinal tract of the pig, the rat, and man: Species- and region-related differences. Scand. J. Gastroenterol. 2000, 35, 501–507. [Google Scholar] [PubMed]
- Klinger, S.; Lange, P.; Brandt, E.; Hustedt, K.; Schroder, B.; Breves, G.; Herrmann, J. Degree of SGLT1 phosphorylation is associated with but does not determine segment-specific glucose transport features in the porcine small intestines. Physiol. Rep. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Skalhegg, B.S.; Tasken, K. Specificity in the cAMP/PKA signaling pathway. Differential expression, regulation, and subcellular localization of subunits of PKA. Front. Biosci. 2000, 5, D678–D693. [Google Scholar] [PubMed]
- Cooper, D.M. Compartmentalization of adenylate cyclase and cAMP signalling. Biochem. Soc. Trans. 2005, 33, 1319–1322. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Murtazina, R.; Cha, B.; Chakraborty, M.; Sarker, R.; Chen, T.E.; Lin, Z.; Hogema, B.M.; de Jonge, H.R.; Seidler, U.; et al. d-glucose acts via sodium/glucose cotransporter 1 to increase NHE3 in mouse jejunal brush border by a Na+/H+ exchange regulatory factor 2-dependent process. Gastroenterology 2011, 140, 560–571. [Google Scholar] [CrossRef] [PubMed]
- Gray, P.C.; Scott, J.D.; Catterall, W.A. Regulation of ion channels by cAMP-dependent protein kinase and A-kinase anchoring proteins. Curr. Opin. Neurobiol. 1998, 8, 330–334. [Google Scholar] [CrossRef]
- Scott, J.D.; Dessauer, C.W.; Tasken, K. Creating order from chaos: Cellular regulation by kinase anchoring. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 187–210. [Google Scholar] [CrossRef] [PubMed]
- Vingtdeux, V.; Giliberto, L.; Zhao, H.; Chandakkar, P.; Wu, Q.; Simon, J.E.; Janle, E.M.; Lobo, J.; Ferruzzi, M.G.; Davies, P.; et al. AMP-activated protein kinase signaling activation by resveratrol modulates amyloid-beta peptide metabolism. J. Biol. Chem. 2010, 285, 9100–9113. [Google Scholar] [CrossRef] [PubMed]
- McCalley, A.E.; Kaja, S.; Payne, A.J.; Koulen, P. Resveratrol and calcium signaling: Molecular mechanisms and clinical relevance. Molecules 2014, 19, 7327–7340. [Google Scholar] [CrossRef] [PubMed]
- Alka, K.; Dolly, J.O.; Ryan, B.J.; Henehan, G.T. New inhibitors of the Kvbeta subunit from mammalian Kv1 potassium channels. Int. J. Biochem. Cell. Biol. 2014, 55C, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.H.; Chen, J.C.; He, Y.L.; Xu, J.J.; Mei, Y.A. Resveratrol inhibits K(v)2.2 currents through the estrogen receptor GPR30-mediated PKC pathway. Am. J. Physiol. Cell Physiol. 2013, 305, C547–C557. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, R.S.; Bogard, A.S.; Gros, R.; Feldman, R.D. Choreographing the adenylyl cyclase signalosome: Sorting out the partners and the steps. Naunyn Schmiedebergs Arch. Pharmacol. 2012, 385, 5–12. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Jejunum | Ileum |
|---|---|---|
| SGLT1 | 1.41 ± 0.87 | 0.82 ± 0.45 |
| PEPT1 | 1.17 ± 0.76 | 2.27 ± 2.21 |
| NHE3 | 1.76 ± 2.12 | 1.06 ± 0.96 |
| pNHE3 Ser552 | 1.44 ± 1.57 | 1.35 ± 0.93 |
| pNHE3 Ser605 | 2.18 ± 1.80 | 0.62 ± 0.28 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klinger, S.; Breves, G. Resveratrol Inhibits Porcine Intestinal Glucose and Alanine Transport: Potential Roles of Na+/K+-ATPase Activity, Protein Kinase A, AMP-Activated Protein Kinase and the Association of Selected Nutrient Transport Proteins with Detergent Resistant Membranes. Nutrients 2018, 10, 302. https://doi.org/10.3390/nu10030302
Klinger S, Breves G. Resveratrol Inhibits Porcine Intestinal Glucose and Alanine Transport: Potential Roles of Na+/K+-ATPase Activity, Protein Kinase A, AMP-Activated Protein Kinase and the Association of Selected Nutrient Transport Proteins with Detergent Resistant Membranes. Nutrients. 2018; 10(3):302. https://doi.org/10.3390/nu10030302
Chicago/Turabian StyleKlinger, Stefanie, and Gerhard Breves. 2018. "Resveratrol Inhibits Porcine Intestinal Glucose and Alanine Transport: Potential Roles of Na+/K+-ATPase Activity, Protein Kinase A, AMP-Activated Protein Kinase and the Association of Selected Nutrient Transport Proteins with Detergent Resistant Membranes" Nutrients 10, no. 3: 302. https://doi.org/10.3390/nu10030302