Effect of Probiotic Bacteria on Microbial Host Defense, Growth, and Immune Function in Human Immunodeficiency Virus Type-1 Infection

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Impact of Nutrient Status on HIV-1 Infection
3. Role of Microbial Translocation on Inflammation and HIV-1 Progression
4. Balance of CD4+ Th17 and T Regulatory (Treg) Lymphocytes and HIV-1 Pathogenesis
5. Effect of HIV-1 and Probiotics on Microflora and Microbiota
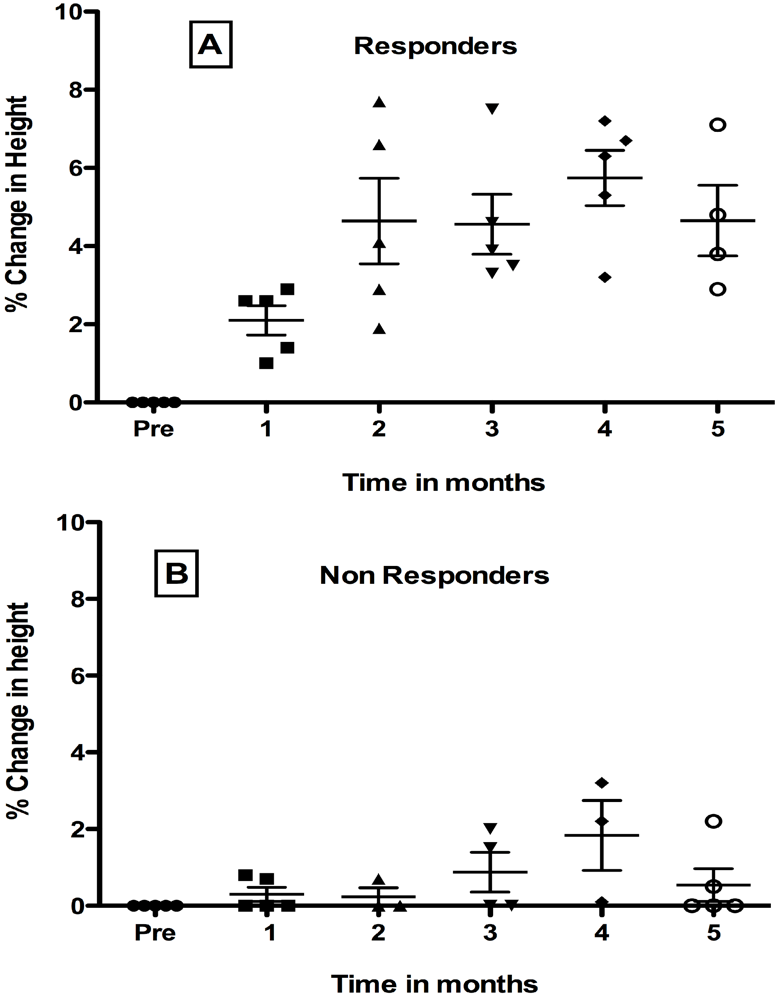
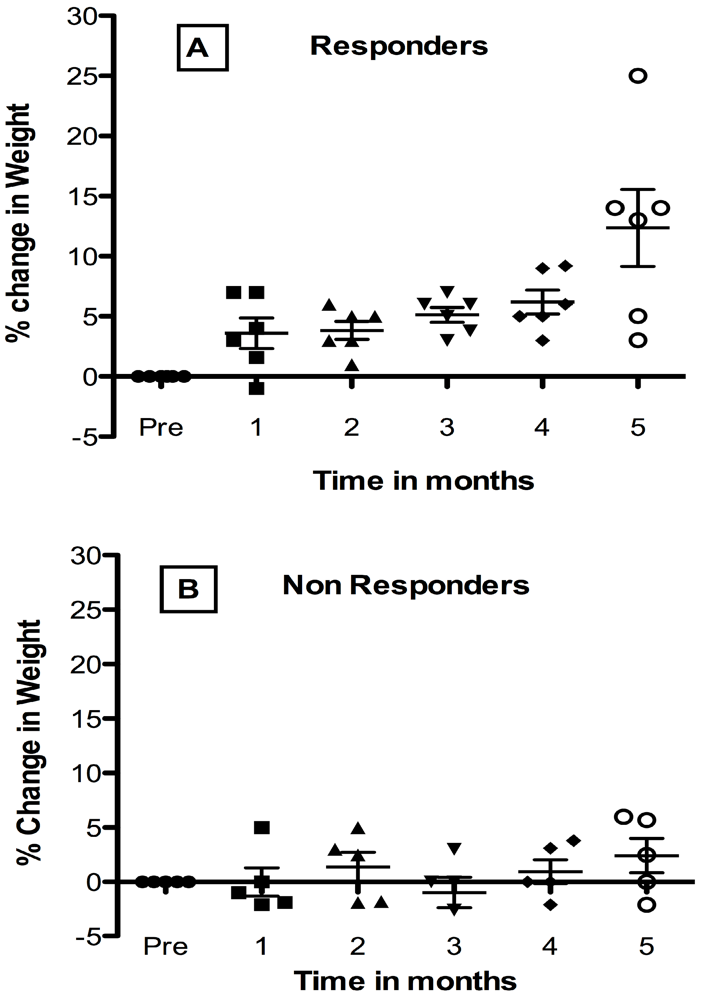
6. Growth Abnormalities in Children with HIV-1 Infection and Response to Probiotic Lactobacilli
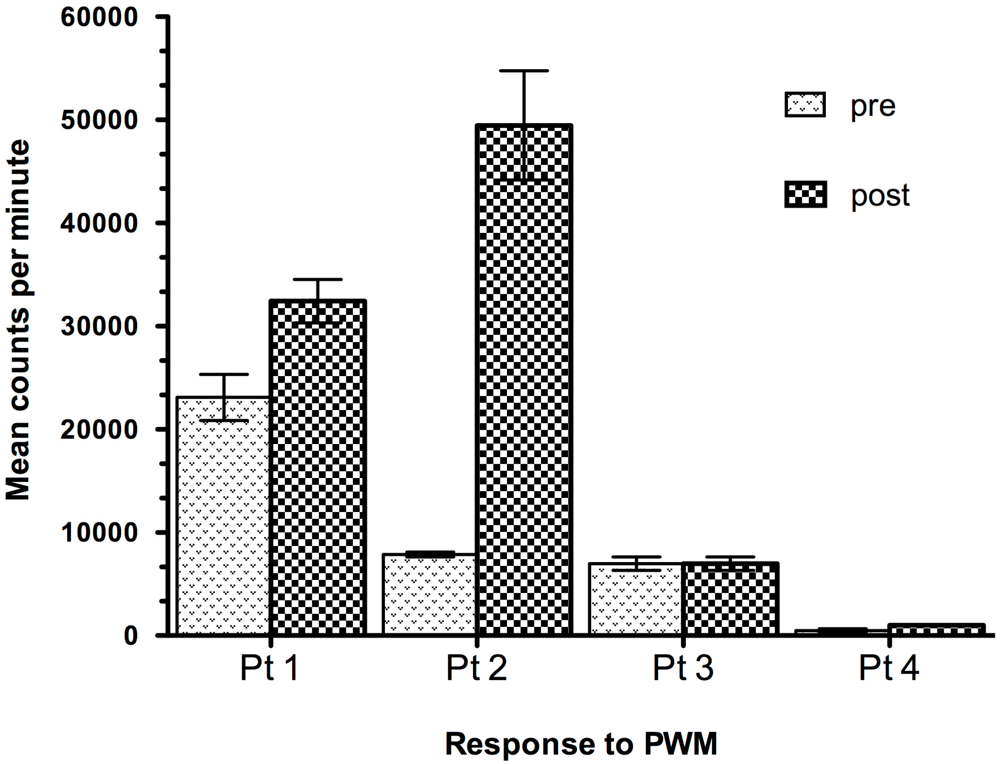
7. Effect of Oral Administration of Lactobacillus in HIV-1 Infected Children with Failure-to Thrive (FTT)



8. Summary
Acknowledgements
References
- Bengmark, S.; Jeppsson, B. Gastrointestinal surface protection and mucosa reconditioning. JPEN J. Parenter. Enteral. Nutr. 1995, 19, 410–415. [Google Scholar]
- Dicks, L.M.; Fraser, T.; ten Doeschate, K.; van Reenen, C.A. Lactic acid bacteria population in children diagnosed with human immunodeficiency virus. J. Paediatr. Child Health 2009, 45, 567–572. [Google Scholar]
- Dwyer, J.T. Nutrition support of HIV+ patients. Henry Ford Hosp. Med. J. 1991, 39, 60–65. [Google Scholar]
- Kaufman, F.R.; Gomperts, E.D. Growth failure in boys with hemophilia and HIV infection. Am. J. Pediatr. Hematol. Oncol. 1989, 11, 292–294. [Google Scholar]
- Kotler, D.P. Wasting syndrome: Nutritional support in HIV infection. AIDS Res. Hum. Retrovir. 1994, 10, 931–934. [Google Scholar]
- Shannon, K.M.; Ammann, A.J. Acquired immune deficiency syndrome in childhood. J. Pediatr. 1985, 106, 332–342. [Google Scholar]
- Cunningham-Rundles, S.; Ahrne, S.; Bengmark, S.; Johann-Liang, R.; Marshall, F.; Metakis, L.; Califano, C.; Dunn, A.M.; Grassey, C.; Hinds, G.; et al. Probiotics and immune response. Am. J. Gastroenterol. 2000, 95, S22–S25. [Google Scholar] [PubMed]
- Cunningham-Rundles, S.; Lin, D.H. Nutrition and the immune system of the gut. Nutrition 1998, 14, 573–579. [Google Scholar]
- Fergusson, P.; Tomkins, A. HIV prevalence and mortality among children undergoing treatment for severe acute malnutrition in sub-Saharan Africa: A systematic review and meta-analysis. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 541–548. [Google Scholar]
- Cunningham-Rundles, S.; McNeeley, D.F.; Moon, A. Mechanisms of nutrient modulation of the immune response. J. Allergy Clin. Immunol. 2005, 115 quiz 1129, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Noyer, C.M.; Simon, D.; Borczuk, A.; Brandt, L.J.; Lee, M.J.; Nehra, V. A double-blind placebo-controlled pilot study of glutamine therapy for abnormal intestinal permeability in patients with AIDS. Am. J. Gastroenterol. 1998, 93, 972–975. [Google Scholar]
- Bushen, O.Y.; Davenport, J.A.; Lima, A.B.; Piscitelli, S.C.; Uzgiris, A.J.; Silva, T.M.; Leite, R.; Kosek, M.; Dillingham, R.A.; Girao, A.; et al. Diarrhea and reduced levels of antiretroviral drugs: Improvement with glutamine or alanyl-glutamine in a randomized controlled trial in northeast Brazil. Clin. Infect. Dis. 2004, 38, 1764–1770. [Google Scholar]
- Kotler, D.P. Nutritional effects and support in the patient with acquired immunodeficiency syndrome. J. Nutr. 1992, 122, 723–727. [Google Scholar]
- Irlam, J.H.; Visser, M.M.; Rollins, N.N.; Siegfried, N. Micronutrient supplementation in children and adults with HIV infection. Cochrane Database Syst. Rev. 2010, 12. [Google Scholar] [CrossRef]
- Fawzi, W.; Msamanga, G.; Spiegelman, D.; Hunter, D.J. Studies of vitamins and minerals and HIV transmission and disease progression. J. Nutr. 2005, 135, 938–944. [Google Scholar]
- Koethe, J.R.; Lukusa, A.; Giganti, M.J.; Chi, B.H.; Nyirenda, C.K.; Limbada, M.I.; Banda, Y.; Stringer, J.S. Association between weight gain and clinical outcomes among malnourished adults initiating antiretroviral therapy in Lusaka, Zambia. J. Acquir. Immune Defic. Syndr. 2010, 53, 507–513. [Google Scholar]
- Winter, H.S.; Oleske, J.M.; Hughes, M.D.; McKinney, R.E., Jr.; Elgie, C.; Powell, C.; Purdue, L.; Puga, A.M.; Jimenez, E.; Scott, G.B.; et al. Randomized controlled trial of feeding a concentrated formula to infants born to women infected by human immunodeficiency virus. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 222–232. [Google Scholar]
- Luoto, R.; Kalliomaki, M.; Laitinen, K.; Delzenne, N.M.; Cani, P.D.; Salminen, S.; Isolauri, E. Initial dietary and microbiological environments deviate in normal-weight compared to overweight children at 10 years of age. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 90–95. [Google Scholar]
- Backhed, F. 99th Dahlem conference on infection, inflammation and chronic inflammatory disorders: The normal gut microbiota in health and disease. Clin. Exp. Immunol. 2010, 160, 80–84. [Google Scholar]
- Gori, A.; Tincati, C.; Rizzardini, G.; Torti, C.; Quirino, T.; Haarman, M.; Ben Amor, K.; van Schaik, J.; Vriesema, A.; Knol, J.; et al. Early impairment of gut function and gut flora supporting a role for alteration of gastrointestinal mucosa in human immunodeficiency virus pathogenesis. J. Clin. Microbiol. 2008, 46, 757–758. [Google Scholar] [PubMed]
- Hummelen, R.; Fernandes, A.D.; Macklaim, J.M.; Dickson, R.J.; Changalucha, J.; Gloor, G.B.; Reid, G. Deep sequencing of the vaginal microbiota of women with HIV. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Spear, G.T.; Sikaroodi, M.; Zariffard, M.R.; Landay, A.L.; French, A.L.; Gillevet, P.M. Comparison of the diversity of the vaginal microbiota in HIV-infected and HIV-uninfected women with or without bacterial vaginosis. J. Infect. Dis. 2008, 198, 1131–1140. [Google Scholar]
- Hummelen, R.; Changalucha, J.; Butamanya, N.L.; Cook, A.; Habbema, J.D.; Reid, G. Lactobacillus rhamnosus GR-1 and L. reuteri RC-14 to prevent or cure bacterial vaginosis among women with HIV. Int. J. Gynaecol. Obstet. 2010, 111, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Gori, A.; Rizzardini, G.; Van’t Land, B.; Amor, K.B.; van Schaik, J.; Torti, C.; Quirino, T.; Tincati, C.; Bandera, A.; Knol, J.; et al. Specific prebiotics modulate gut microbiota and immune activation in HAART-naive HIV-infected adults: Results of the “COPA” pilot randomized trial. Mucosal Immunol. 2011, 4, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Peoples, J.D.; Cheung, S.; Nesin, M.; Lin, H.; Tatad, A.M.; Hoang, D.; Perlman, J.M.; Cunningham-Rundles, S. Neonatal cord blood subsets and cytokine response to bacterial antigens. Am. J. Perinatol. 2009, 26, 647–657. [Google Scholar]
- Stencel-Gabriel, K.; Gabriel, I.; Wiczkowski, A.; Paul, M.; Olejek, A. Prenatal priming of cord blood T lymphocytes by microbiota in the maternal vagina. Am. J. Reprod. Immunol. 2009, 61, 246–252. [Google Scholar]
- Karlsson, H.; Hessle, C.; Rudin, A. Innate immune responses of human neonatal cells to bacteria from the normal gastrointestinal flora. Infect. Immun. 2002, 70, 6688–6696. [Google Scholar]
- Tatad, A.M.; Nesin, M.; Peoples, J.; Cheung, S.; Lin, H.; Sison, C.; Perlman, J.; Cunningham-Rundles, S. Cytokine Expression in response to bacterial antigens in preterm and term infant cord blood monocytes. Neonatology 2007, 94, 8–15. [Google Scholar]
- Fak, F.; Ahrne, S.; Molin, G.; Jeppsson, B.; Westrom, B. Microbial manipulation of the rat dam changes bacterial colonization and alters properties of the gut in her offspring. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G148–G154. [Google Scholar]
- Serwadda, D.; Mugerwa, R.D.; Sewankambo, N.K.; Lwegaba, A.; Carswell, J.W.; Kirya, G.B.; Bayley, A.C.; Downing, R.G.; Tedder, R.S.; Clayden, S.A.; et al. Slim disease: A new disease in Uganda and its association with HTLV-III infection. Lancet 1985, 2, 849–852. [Google Scholar]
- Revision of the CDC surveillance case definition for acquired immunodeficiency syndrome. MMWR Morb. Mortal. Wkly. Rep. 1987, 36 (Suppl. 1), 1–15.
- Kotler, D.P.; Tierney, A.R.; Wang, J.; Pierson, R.N., Jr. Magnitude of body-cell-mass depletion and the timing of death from wasting in AIDS. Am. J. Clin. Nutr. 1989, 50, 444–447. [Google Scholar]
- Kotler, D.P.; Reka, S.; Clayton, F. Intestinal mucosal inflammation associated with human immunodeficiency virus infection. Dig. Dis. Sci. 1993, 38, 1119–1127. [Google Scholar]
- Pantaleo, G.; Graziosi, C.; Demarest, J.F.; Butini, L.; Montroni, M.; Fox, C.H.; Orenstein, J.M.; Kotler, D.P.; Fauci, A.S. HIV infection is active and progressive in lymphoid tissue during the clinically latent stage of disease. Nature 1993, 362, 355–358. [Google Scholar]
- Wanke, C. Pathogenesis and Consequences of HIV-Associated Wasting. J. Acquir. Immune Defic. Syndr. 2004, 37 (Suppl. 5), S277–S279. [Google Scholar] [CrossRef] [PubMed]
- Moldawer, L.L.; Sattler, F.R. Human immunodeficiency virus-associated wasting and mechanisms of cachexia associated with inflammation. Semin. Oncol. 1998, 25, 73–81. [Google Scholar]
- Batterham, M.J. Investigating heterogeneity in studies of resting energy expenditure in persons with HIV/AIDS: A meta-analysis. Am. J. Clin. Nutr. 2005, 81, 702–713. [Google Scholar]
- De Pee, S.; Semba, R.D. Role of nutrition in HIV infection: Review of evidence for more effective programming in resource-limited settings. Food Nutr. Bull. 2010, 31, S313–S344. [Google Scholar]
- Kotler, D.; Wanke, C. Management of HIV wasting syndrome: A consensus conference. J. Acquir. Immune Defic. Syndr. 2004, 37, S261–S288. [Google Scholar]
- Crenn, P.; Rakotoanbinina, B.; Raynaud, J.J.; Thuillier, F.; Messing, B.; Melchior, J.C. Hyperphagia contributes to the normal body composition and protein-energy balance in HIV-infected asymptomatic men. J. Nutr. 2004, 134, 2301–2306. [Google Scholar]
- Crenn, P.; de Truchis, P.; Neveux, N.; Galperine, T.; Cynober, L.; Melchior, J.C. Plasma citrulline is a biomarker of enterocyte mass and an indicator of parenteral nutrition in HIV-infected patients. Am. J. Clin. Nutr. 2009, 90, 587–594. [Google Scholar]
- Ndekha, M.; van Oosterhout, J.J.; Saloojee, H.; Pettifor, J.; Manary, M. Nutritional status of Malawian adults on antiretroviral therapy 1 year after supplementary feeding in the first 3 months of therapy. Trop. Med. Int. Health 2009, 14, 1059–1063. [Google Scholar]
- Falutz, J.; Tsoukas, C.; Cardno, T. Serum zinc in homosexual men. Clin. Chem. 1989, 35, 704–705. [Google Scholar]
- Dworkin, B.M. Selenium deficiency in HIV infection and the acquired immunodeficiency syndrome (AIDS). Chem. Biol. Interact. 1994, 91, 181–186. [Google Scholar]
- Harriman, G.R.; Smith, P.D.; Horne, M.K.; Fox, C.H.; Koenig, S.; Lack, E.E.; Lane, H.C.; Fauci, A.S. Vitamin B12 malabsorption in patients with acquired immunodeficiency syndrome. Arch. Intern. Med. 1989, 149, 2039–2041. [Google Scholar]
- Mburu, A.S.; Thurnham, D.I.; Mwaniki, D.L.; Muniu, E.M.; Alumasa, F.M. The influence of inflammation on plasma zinc concentration in apparently healthy, HIV+ Kenyan adults and zinc responses after a multi-micronutrient supplement. Eur. J. Clin. Nutr. 2010, 64, 510–517. [Google Scholar]
- Tang, A.M.; Graham, N.M.; Semba, R.D.; Saah, A.J. Association between serum vitamin A and E levels and HIV-1 disease progression. Aids 1997, 11, 613–620. [Google Scholar]
- Tang, A.M.; Graham, N.M.; Chandra, R.K.; Saah, A.J. Low serum vitamin B-12 concentrations are associated with faster human immunodeficiency virus type 1 (HIV-1) disease progression. J. Nutr. 1997, 127, 345–351. [Google Scholar]
- Forrester, J.E.; Wang, X.D.; Knox, T.A.; Borek, C.G.; Tang, A.M.; Johnson, E.J. Factors associated with serum retinol, alpha-tocopherol, carotenoids, and selenium in Hispanics with problems of HIV, chronic hepatitis C, and drug. J. Public Health Policy 2009, 30, 285–299. [Google Scholar]
- Jones, C.Y.; Tang, A.M.; Forrester, J.E.; Huang, J.; Hendricks, K.M.; Knox, T.A.; Spiegelman, D.; Semba, R.D.; Woods, M.N. Micronutrient levels and HIV disease status in HIV-infected patients on highly active antiretroviral therapy in the Nutrition for Healthy Living cohort. J. Acquir. Immune Defic. Syndr. 2006, 43, 475–482. [Google Scholar]
- Baum, M.K.; Lai, S.; Sales, S.; Page, J.B.; Campa, A. Randomized, controlled clinical trial of zinc supplementation to prevent immunological failure in HIV-infected adults. Clin. Infect. Dis. 2010, 50, 1653–1660. [Google Scholar]
- Dao, C.N.; Patel, P.; Overton, E.T.; Rhame, F.; Pals, S.L.; Johnson, C.; Bush, T.; Brooks, J.T. Low vitamin D among HIV-infected adults: Prevalence of and risk factors for low vitamin D Levels in a cohort of HIV-infected adults and comparison to prevalence among adults in the US general population. Clin. Infect. Dis. 2011, 52, 396–405. [Google Scholar]
- Torres, C.; Sanchez de la Torre, M.; Garcia-Moruja, C.; Carrero, A.J.; Trujillo Mdel, M.; Fibla, J.; Caruz, A. Immunophenotype of vitamin D receptor polymorphism associated to risk of HIV-1 infection and rate of disease progression. Curr. HIV Res. 2010, 8, 487–492. [Google Scholar]
- Hendricks, K.M.; Mwamburi, D.M.; Newby, P.K.; Wanke, C.A. Dietary patterns and health and nutrition outcomes in men living with HIV infection. Am. J. Clin. Nutr. 2008, 88, 1584–1592. [Google Scholar]
- Woods, M.N.; Tang, A.M.; Forrester, J.; Jones, C.; Hendricks, K.; Ding, B.; Knox, T.A. Effect of dietary intake and protease inhibitors on serum vitamin B12 levels in a cohort of human immunodeficiency virus-positive patients. Clin. Infect. Dis. 2003, 37 (Suppl. 2), S124–S131. [Google Scholar] [CrossRef] [PubMed]
- Stone, C.A.; Kawai, K.; Kupka, R.; Fawzi, W.W. Role of selenium in HIV infection. Nutr. Rev. 2010, 68, 671–681. [Google Scholar]
- Hurwitz, B.E.; Klaus, J.R.; Llabre, M.M.; Gonzalez, A.; Lawrence, P.J.; Maher, K.J.; Greeson, J.M.; Baum, M.K.; Shor-Posner, G.; Skyler, J.S.; Schneiderman, N. Suppression of human immunodeficiency virus type 1 viral load with selenium supplementation: A randomized controlled trial. Arch. Intern. Med. 2007, 167, 148–154. [Google Scholar]
- Sankaran, S.; Guadalupe, M.; Reay, E.; George, M.D.; Flamm, J.; Prindiville, T.; Dandekar, S. Gut mucosal T cell responses and gene expression correlate with protection against disease in long-term HIV-1-infected nonprogressors. Proc. Natl. Acad. Sci. USA 2005, 102, 9860–9865. [Google Scholar]
- Stone, J.D.; Heise, C.C.; Miller, C.J.; Halsted, C.H.; Dandekar, S. Development of malabsorption and nutritional complications in simian immunodeficiency virus-infected rhesus macaques. Aids 1994, 8, 1245–1256. [Google Scholar]
- Stein, T.P.; Koerner, B.; Schluter, M.D.; Leskiw, M.J.; Gaprindachvilli, T.; Richards, E.W.; Cope, F.O.; Condolucci, D. Weight loss, the gut and the inflammatory response in aids patients. Cytokine 1997, 9, 143–147. [Google Scholar]
- Lane, H.C.; Masur, H.; Edgar, L.C.; Whalen, G.; Rook, A.H.; Fauci, A.S. Abnormalities of B-cell activation and immunoregulation in patients with the acquired immunodeficiency syndrome. N. Engl. J. Med. 1983, 309, 453–458. [Google Scholar]
- Giorgi, J.V.; Hultin, L.E.; McKeating, J.A.; Johnson, T.D.; Owens, B.; Jacobson, L.P.; Shih, R.; Lewis, J.; Wiley, D.J.; Phair, J.P.; et al. Shorter survival in advanced human immunodeficiency virus type 1 infection is more closely associated with T lymphocyte activation than with plasma virus burden or virus chemokine coreceptor usage. J. Infect. Dis. 1999, 179, 859–870. [Google Scholar] [PubMed]
- Grossman, Z.; Meier-Schellersheim, M.; Paul, W.E.; Picker, L.J. Pathogenesis of HIV infection: What the virus spares is as important as what it destroys. Nat. Med. 2006, 12, 289–295. [Google Scholar]
- Mattapallil, J.J.; Douek, D.C.; Hill, B.; Nishimura, Y.; Martin, M.; Roederer, M. Massive infection and loss of memory CD4+ T cells in multiple tissues during acute SIV infection. Nature 2005, 434, 1093–1097. [Google Scholar]
- Guadalupe, M.; Reay, E.; Sankaran, S.; Prindiville, T.; Flamm, J.; McNeil, A.; Dandekar, S. Severe CD4+ T-cell depletion in gut lymphoid tissue during primary human immunodeficiency virus type 1 infection and substantial delay in restoration following highly active antiretroviral therapy. J. Virol. 2003, 77, 11708–11717. [Google Scholar]
- Brenchley, J.M.; Price, D.A.; Schacker, T.W.; Asher, T.E.; Silvestri, G.; Rao, S.; Kazzaz, Z.; Bornstein, E.; Lambotte, O.; Altmann, D.; et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat. Med. 2006, 12, 1365–1371. [Google Scholar] [PubMed]
- Emu, B.; Sinclair, E.; Favre, D.; Moretto, W.J.; Hsue, P.; Hoh, R.; Martin, J.N.; Nixon, D.F.; McCune, J.M.; Deeks, S.G. Phenotypic, functional, and kinetic parameters associated with apparent T-cell control of human immunodeficiency virus replication in individuals with and without antiretroviral treatment. J. Virol. 2005, 79, 14169–14178. [Google Scholar] [PubMed]
- Yin, L.; Rodriguez, C.A.; Hou, W.; Potter, O.; Caplan, M.J.; Goodenow, M.M.; Sleasman, J.W. Antiretroviral therapy corrects HIV-1-induced expansion of CD8+ CD45RA+ CD27− CD11a(bright) activated T cells. J. Allergy Clin. Immunol. 2008, 122, 166–172. [Google Scholar] [PubMed]
- Silvestri, G.; Paiardini, M.; Pandrea, I.; Lederman, M.M.; Sodora, D.L. Understanding the benign nature of SIV infection in natural hosts. J. Clin. Invest. 2007, 117, 3148–3154. [Google Scholar]
- Estes, J.D.; Harris, L.D.; Klatt, N.R.; Tabb, B.; Pittaluga, S.; Paiardini, M.; Barclay, G.R.; Smedley, J.; Pung, R.; Oliveira, K.M.; et al. Damaged intestinal epithelial integrity linked to microbial translocation in pathogenic simian immunodeficiency virus infections. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef]
- Hofer, U.; Schlaepfer, E.; Baenziger, S.; Nischang, M.; Regenass, S.; Schwendener, R.; Kempf, W.; Nadal, D.; Speck, R.F. Inadequate clearance of translocated bacterial products in HIV-infected humanized mice. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef] [Green Version]
- Rempel, H.; Sun, B.; Calosing, C.; Pillai, S.K.; Pulliam, L. Interferon-alpha drives monocyte gene expression in chronic unsuppressed HIV-1 infection. Aids 2010, 24, 1415–1423. [Google Scholar]
- Rajasuriar, R.; Booth, D.; Solomon, A.; Chua, K.; Spelman, T.; Gouillou, M.; Schlub, T.E.; Davenport, M.; Crowe, S.; Elliott, J.; et al. Biological determinants of immune reconstitution in HIV-infected patients receiving antiretroviral therapy: The role of interleukin 7 and interleukin 7 receptor alpha and microbial translocation. J. Infect. Dis. 2010, 202, 1254–1264. [Google Scholar] [CrossRef] [PubMed]
- Piconi, S.; Trabattoni, D.; Gori, A.; Parisotto, S.; Magni, C.; Meraviglia, P.; Bandera, A.; Capetti, A.; Rizzardini, G.; Clerici, M. Immune activation, apoptosis, and Treg activity are associated with persistently reduced CD4+ T-cell counts during antiretroviral therapy. Aids 2010, 24, 1991–2000. [Google Scholar] [CrossRef] [PubMed]
- Wallet, M.A.; Rodriguez, C.A.; Yin, L.; Saporta, S.; Chinratanapisit, S.; Hou, W.; Sleasman, J.W.; Goodenow, M.M. Microbial translocation induces persistent macrophage activation unrelated to HIV-1 levels or T-cell activation following therapy. Aids 2010, 24, 1281–1290. [Google Scholar]
- Gordon, S.N.; Klatt, N.R.; Bosinger, S.E.; Brenchley, J.M.; Milush, J.M.; Engram, J.C.; Dunham, R.M.; Paiardini, M.; Klucking, S.; Danesh, A.; et al. Severe depletion of mucosal CD4+ T cells in AIDS-free simian immunodeficiency virus-infected sooty mangabeys. J. Immunol. 2007, 179, 3026–3034. [Google Scholar] [PubMed]
- Brenchley, J.M.; Paiardini, M.; Knox, K.S.; Asher, A.I.; Cervasi, B.; Asher, T.E.; Scheinberg, P.; Price, D.A.; Hage, C.A.; Kholi, L.M.; et al. Differential Th17 CD4 T-cell depletion in pathogenic and nonpathogenic lentiviral infections. Blood 2008, 112, 2826–2835. [Google Scholar] [PubMed]
- Ancuta, P.; Monteiro, P.; Sekaly, R.P. Th17 lineage commitment and HIV-1 pathogenesis. Curr. Opin. HIV AIDS 2010, 5, 158–165. [Google Scholar]
- Gaboriau-Routhiau, V.; Rakotobe, S.; Lecuyer, E.; Mulder, I.; Lan, A.; Bridonneau, C.; Rochet, V.; Pisi, A.; de Paepe, M.; Brandi, G.; et al. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity 2009, 31, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Aujla, S.J.; Dubin, P.J.; Kolls, J.K. Th17 cells and mucosal host defense. Semin. Immunol. 2007, 19, 377–382. [Google Scholar]
- Mitsdoerffer, M.; Lee, Y.; Jager, A.; Kim, H.J.; Korn, T.; Kolls, J.K.; Cantor, H.; Bettelli, E.; Kuchroo, V.K. Proinflammatory T helper type 17 cells are effective B-cell helpers. Proc. Natl. Acad. Sci. USA 2010, 107, 14292–14297. [Google Scholar]
- Dong, C. TH17 cells in development: An updated view of their molecular identity and genetic programming. Nat. Rev. Immunol. 2008, 8, 337–348. [Google Scholar]
- Chiba, Y.; Shida, K.; Nagata, S.; Wada, M.; Bian, L.; Wang, C.; Shimizu, T.; Yamashiro, Y.; Kiyoshima-Shibata, J.; Nanno, M.; Nomoto, K. Well-controlled proinflammatory cytokine responses of Peyer’s patch cells to probiotic Lactobacillus casei. Immunology 2010, 130, 352–362. [Google Scholar]
- Cecchinato, V.; Trindade, C.J.; Laurence, A.; Heraud, J.M.; Brenchley, J.M.; Ferrari, M.G.; Zaffiri, L.; Tryniszewska, E.; Tsai, W.P.; Vaccari, M.; et al. Altered balance between Th17 and Th1 cells at mucosal sites predicts AIDS progression in simian immunodeficiency virus-infected macaques. Mucosal Immunol. 2008, 1, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Favre, D.; Lederer, S.; Kanwar, B.; Ma, Z.M.; Proll, S.; Kasakow, Z.; Mold, J.; Swainson, L.; Barbour, J.D.; Baskin, C.R.; et al. Critical loss of the balance between Th17 and T regulatory cell populations in pathogenic SIV infection. PLoS Pathog. 2009, 5. [Google Scholar] [CrossRef]
- Perez-Cano, F.J.; Dong, H.; Yaqoob, P. In vitro immunomodulatory activity of Lactobacillus fermentum CECT5716 and Lactobacillus salivarius CECT5713: Two probiotic strains isolated from human breast milk. Immunobiology 2010, 215, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Smits, H.H.; Engering, A.; van der Kleij, D.; de Jong, E.C.; Schipper, K.; van Capel, T.M.; Zaat, B.A.; Yazdanbakhsh, M.; Wierenga, E.A.; van Kooyk, Y.; et al. Selective probiotic bacteria induce IL-10-producing regulatory T cells in vitro by modulating dendritic cell function through dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin. J. Allergy Clin. Immunol. 2005, 115, 1260–1267. [Google Scholar] [PubMed]
- Prendergast, A.; Prado, J.G.; Kang, Y.H.; Chen, F.; Riddell, L.A.; Luzzi, G.; Goulder, P.; Klenerman, P. HIV-1 infection is characterized by profound depletion of CD161+ Th17 cells and gradual decline in regulatory T cells. Aids 2010, 24, 491–502. [Google Scholar]
- Denning, T.L.; Wang, Y.C.; Patel, S.R.; Williams, I.R.; Pulendran, B. Lamina propria macrophages and dendritic cells differentially induce regulatory and interleukin 17-producing T cell responses. Nat. Immunol. 2007, 8, 1086–1094. [Google Scholar]
- Mucida, D.; Park, Y.; Kim, G.; Turovskaya, O.; Scott, I.; Kronenberg, M.; Cheroutre, H. Reciprocal TH17 and regulatory T cell differentiation mediated by retinoic acid. Science 2007, 317, 256–260. [Google Scholar]
- Chaudhry, A.; Rudra, D.; Treuting, P.; Samstein, R.M.; Liang, Y.; Kas, A.; Rudensky, A.Y. CD4+ regulatory T cells control TH17 responses in a Stat3-dependent manner. Science 2009, 326, 986–991. [Google Scholar]
- Elyaman, W.; Bradshaw, E.M.; Uyttenhove, C.; Dardalhon, V.; Awasthi, A.; Imitola, J.; Bettelli, E.; Oukka, M.; van Snick, J.; Renauld, J.C.; et al. IL-9 induces differentiation of TH17 cells and enhances function of FoxP3+ natural regulatory T cells. Proc. Natl. Acad. Sci. USA 2009, 106, 12885–12890. [Google Scholar]
- Lavasani, S.; Dzhambazov, B.; Nouri, M.; Fak, F.; Buske, S.; Molin, G.; Thorlacius, H.; Alenfall, J.; Jeppsson, B.; Westrom, B. A novel probiotic mixture exerts a therapeutic effect on experimental autoimmune encephalomyelitis mediated by IL-10 producing regulatory T cells. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Kwon, H.K.; Lee, C.G.; So, J.S.; Chae, C.S.; Hwang, J.S.; Sahoo, A.; Nam, J.H.; Rhee, J.H.; Hwang, K.C.; Im, S.H. Generation of regulatory dendritic cells and CD4+ Foxp3+ T cells by probiotics administration suppresses immune disorders. Proc. Natl. Acad. Sci. USA 2010, 107, 2159–2164. [Google Scholar]
- De Roock, S.; van Elk, M.; van Dijk, M.E.; Timmerman, H.M.; Rijkers, G.T.; Prakken, B.J.; Hoekstra, M.O.; de Kleer, I.M. Lactic acid bacteria differ in their ability to induce functional regulatory T cells in humans. Clin. Exp. Allergy 2010, 40, 103–110. [Google Scholar]
- Wolf, B.W.; Wheeler, K.B.; Ataya, D.G.; Garleb, K.A. Safety and tolerance of Lactobacillus reuteri supplementation to a population infected with the human immunodeficiency virus. Food Chem. Toxicol. 1998, 36, 1085–1094. [Google Scholar]
- Nugent, R.P.; Krohn, M.A.; Hillier, S.L. Reliability of diagnosing bacterial vaginosis is improved by a standardized method of gram stain interpretation. J. Clin. Microbiol. 1991, 29, 297–301. [Google Scholar]
- Cu-Uvin, S.; Hogan, J.W.; Caliendo, A.M.; Harwell, J.; Mayer, K.H.; Carpenter, C.C. Association between bacterial vaginosis and expression of human immunodeficiency virus type 1 RNA in the female genital tract. Clin. Infect. Dis. 2001, 33, 894–896. [Google Scholar]
- Sha, B.E.; Zariffard, M.R.; Wang, Q.J.; Chen, H.Y.; Bremer, J.; Cohen, M.H.; Spear, G.T. Female genital-tract HIV load correlates inversely with Lactobacillus species but positively with bacterial vaginosis and Mycoplasma hominis. J. Infect. Dis. 2005, 191, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Spear, G.T.; Gilbert, D.; Sikaroodi, M.; Doyle, L.; Green, L.; Gillevet, P.M.; Landay, A.L.; Veazey, R.S. Identification of rhesus macaque genital microbiota by 16S pyrosequencing shows similarities to human bacterial vaginosis: Implications for use as an animal model for HIV vaginal infection. AIDS Res. Hum. Retrovir. 2010, 26, 193–200. [Google Scholar]
- Mitchell, C.; Moreira, C.; Fredricks, D.; Paul, K.; Caliendo, A.M.; Kurpewski, J.; Ingersoll, J.; Cu-Uvin, S. Detection of fastidious vaginal bacteria in women with HIV infection and bacterial vaginosis. Infect. Dis. Obstet. Gynecol. 2009, 2009. [Google Scholar] [CrossRef]
- Farquhar, C.; Mbori-Ngacha, D.; Overbaugh, J.; Wamalwa, D.; Harris, J.; Bosire, R.; John-Stewart, G. Illness during pregnancy and bacterial vaginosis are associated with in-utero HIV-1 transmission. Aids 2010, 24, 153–155. [Google Scholar] [PubMed]
- Anukam, K.C.; Osazuwa, E.; Osemene, G.I.; Ehigiagbe, F.; Bruce, A.W.; Reid, G. Clinical study comparing probiotic Lactobacillus GR-1 and RC-14 with metronidazole vaginal gel to treat symptomatic bacterial vaginosis. Microbes Infect. 2006, 8, 2772–2776. [Google Scholar]
- Senok, A.C.; Verstraelen, H.; Temmerman, M.; Botta, G.A. Probiotics for the treatment of bacterial vaginosis. Cochrane Database Syst. Rev. 2009. [Google Scholar] [CrossRef]
- Parent, D.; Bossens, M.; Bayot, D.; Kirkpatrick, C.; Graf, F.; Wilkinson, F.E.; Kaiser, R.R. Therapy of bacterial vaginosis using exogenously-applied Lactobacilli acidophili and a low dose of estriol: A placebo-controlled multicentric clinical trial. Arzneimittelforschung 1996, 46, 68–73. [Google Scholar]
- Anukam, K.; Osazuwa, E.; Ahonkhai, I.; Ngwu, M.; Osemene, G.; Bruce, A.W.; Reid, G. Augmentation of antimicrobial metronidazole therapy of bacterial vaginosis with oral probiotic Lactobacillus rhamnosus GR-1 and Lactobacillus reuteri RC-14: Randomized, double-blind, placebo controlled trial. Microbes Infect. 2006, 8, 1450–1454. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, L.S.; Ferreira, S.M.; Silva, A., Jr.; Villoria, G.E.; Costinha, L.H.; Souto, R.; Uzeda, M.D.; Colombo, A.P. Association of T CD4 lymphocyte levels and subgingival microbiota of chronic periodontitis in HIV-infected Brazilians under HAART. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2004, 97, 196–203. [Google Scholar]
- Aas, J.A.; Barbuto, S.M.; Alpagot, T.; Olsen, I.; Dewhirst, F.E.; Paster, B.J. Subgingival plaque microbiota in HIV positive patients. J. Clin. Periodontol. 2007, 34, 189–195. [Google Scholar]
- Tsang, C.S.; Samaranayake, L.P. Predominant cultivable subgingival microbiota of healthy and HIV-infected ethnic Chinese. APMIS 2001, 109, 117–126. [Google Scholar]
- Murray, P.A.; Grassi, M.; Winkler, J.R. The microbiology of HIV-associated periodontal lesions. J. Clin. Periodontol. 1989, 16, 636–642. [Google Scholar]
- Silva-Boghossian, C.; Castro, G.F.; Teles, R.P.; de Souza, I.P.; Colombo, A.P. Salivary microbiotaof HIV-positive children and its correlation with HIV status, oral diseases, and total secretory IgA. Int. J. Paediatr. Dent. 2008, 18, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Anukam, K.C.; Osazuwa, E.O.; Osadolor, H.B.; Bruce, A.W.; Reid, G. Yogurt containing probiotic Lactobacillus rhamnosus GR-1 and L. reuteri RC-14 helps resolve moderate diarrhea and increases CD4 count in HIV/AIDS patients. J. Clin. Gastroenterol. 2008, 42, 239–243. [Google Scholar] [PubMed]
- Reid, G.; Anand, S.; Bingham, M.O.; Mbugua, G.; Wadstrom, T.; Fuller, R.; Anukam, K.; Katsivo, M. Probiotics for the developing world. J. Clin. Gastroenterol. 2005, 39, 485–488. [Google Scholar]
- Resta-Lenert, S.; Barrett, K.E. Live probiotics protect intestinal epithelial cells from the effects of infection with enteroinvasive Escherichia coli (EIEC). Gut 2003, 52, 988–997. [Google Scholar]
- Resta-Lenert, S.C.; Barrett, K.E. Modulation of intestinal barrier properties by probiotics: Role in reversing colitis. Ann. N. Y. Acad. Sci. 2009, 1165, 175–182. [Google Scholar]
- Guarino, A.; Canani, R.B.; Spagnuolo, M.I.; Albano, F.; Di Benedetto, L. Oral bacterial therapy reduces the duration of symptoms and of viral excretion in children with mild diarrhea. J. Pediatr.Gastroenterol. Nutr. 1997, 25, 516–519. [Google Scholar]
- Miettinen, M.; Vuopio-Varkila, J.; Varkila, K. Production of human tumor necrosis factor alpha, interleukin-6, and interleukin-10 is induced by lactic acid bacteria. Infect. Immun. 1996, 64, 5403–5405. [Google Scholar] [PubMed]
- Salminen, M.K.; Tynkkynen, S.; Rautelin, H.; Poussa, T.; Saxelin, M.; Ristola, M.; Valtonen, V.; Jarvinen, A. The efficacy and safety of probiotic Lactobacillus rhamnosus GG on prolonged, noninfectious diarrhea in HIV Patients on antiretroviral therapy: A randomized, placebo-controlled, crossover study. HIV Clin. Trials 2004, 5, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Kerac, M.; Bunn, J.; Seal, A.; Thindwa, M.; Tomkins, A.; Sadler, K.; Bahwere, P.; Collins, S. Probiotics and prebiotics for severe acute malnutrition (PRONUT study): A double-blind efficacy randomised controlled trial in Malawi. Lancet 2009, 374, 136–144. [Google Scholar]
- Trois, L.; Cardoso, E.M.; Miura, E. Use of probiotics in HIV-infected children: A randomized double-blind controlled study. J. Trop. Pediatr. 2008, 54, 19–24. [Google Scholar]
- Monachese, M.; Cunningham-Rundles, S.; Diaz, M.A.; Guerrant, R.; Hummelen, R.; Kemperman, R.; Kerac, M.; Kort, R.; Merenstein, D.J.; Panigrahi, P.; et al. Probiotics and prebiotics to combat enteric infections and HIV in the developing world: A consensus report. Gut Microbes 2011, 2, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Black, R.E.; Morris, S.S.; Bryce, J. Where and why are 10 million children dying every year? Lancet 2003, 361, 2226–2234. [Google Scholar] [CrossRef] [PubMed]
- Grantham-McGregor, S.; Cheung, Y.B.; Cueto, S.; Glewwe, P.; Richter, L.; Strupp, B. Developmental potential in the first 5 years for children in developing countries. Lancet 2007, 369, 60–70. [Google Scholar]
- Moye, J., Jr.; Rich, K.C.; Kalish, L.A.; Sheon, A.R.; Diaz, C.; Cooper, E.R.; Pitt, J.; Handelsman, E. Natural history of somatic growth in infants born to women infected by human immunodeficiency virus. Women and Infants Transmission Study Group. J. Pediatr. 1996, 128, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Cunningham-Rundles, S.; Ahrn, S.; Abuav-Nussbaum, R.; Dnistrian, A. Development of immunocompetence: Role of micronutrients and microorganisms. Nutr. Rev. 2002, 60, S68–S72. [Google Scholar]
- Taha, T.E.; Dallabetta, G.A.; Canner, J.K.; Chiphangwi, J.D.; Liomba, G.; Hoover, D.R.; Miotti, P.G. The effect of human immunodeficiency virus infection on birthweight, and infant and child mortality in urban Malawi. Int. J. Epidemiol. 1995, 24, 1022–1029. [Google Scholar]
- Filteau, S.; Baisley, K.; Chisenga, M.; Kasonka, L.; Gibson, R.S. Provision of micronutrient-fortified food from 6 months of age does not permit HIV-exposed, uninfected Zambian children to catch up in growth to HIV-unexposed children: A randomised controlled trial. J. Acquir. Immune Defic. Syndr. 2011, 56, 166–175. [Google Scholar]
- Nielsen, S.; Nielsen, D.S.; Lauritzen, L.; Jakobsen, M.; Michaelsen, K.F. Impact of diet on the intestinal microbiota in 10-month-old infants. J. Pediatr. Gastroenterol. Nutr. 2007, 44, 613–618. [Google Scholar]
- Iwata, M.; Hirakiyama, A.; Eshima, Y.; Kagechika, H.; Kato, C.; Song, S.Y. Retinoic acid imprints gut-homing specificity on T cells. Immunity 2004, 21, 527–538. [Google Scholar]
- Mora, J.R.; Iwata, M.; Eksteen, B.; Song, S.Y.; Junt, T.; Senman, B.; Otipoby, K.L.; Yokota, A.; Takeuchi, H.; Ricciardi-Castagnoli, P.; et al. Generation of gut-homing IgA-secreting B cells by intestinal dendritic cells. Science 2006, 314, 1157–1160. [Google Scholar] [PubMed]
- Cunningham-Rundles, S.; Lin, H.; Ho-Lin, D.; Dnistrian, A.; Cassileth, B.R.; Perlman, J.M. Role of nutrients in the development of neonatal immune response. Nutr. Rev. 2009, 67 (Suppl. 2), S152–S163. [Google Scholar] [CrossRef] [PubMed]
- Steenhout, P.G.; Rochat, F.; Hager, C. The effect of Bifidobacterium lactis on the growth of infants: A pooled analysis of randomized controlled studies. Ann. Nutr. Metab. 2009, 55, 334–340. [Google Scholar]
- Taye, B.; Shiferaw, S.; Enquselassie, F. The impact of malnutrition in survival of HIV infected children after initiation of antiretroviral treatment (ART). Ethiop. Med. J. 2010, 48, 1–10. [Google Scholar]
- Miller, T.L.; Mawn, B.E.; Orav, E.J.; Wilk, D.; Weinberg, G.A.; Nicchitta, J.; Furuta, L.; Cutroni, R.; McIntosh, K.; Burchett, S.K.; et al. The effect of protease inhibitor therapy on growth and body composition in human immunodeficiency virus type 1-infected children. Pediatrics 2001, 107. [Google Scholar] [CrossRef]
- Weigel, R.; Phiri, S.; Chiputula, F.; Gumulira, J.; Brinkhof, M.; Gsponer, T.; Tweya, H.; Egger, M.; Keiser, O. Growth response to antiretroviral treatment in HIV-infected children: A cohort study from Lilongwe, Malawi. Trop. Med. Int. Health 2010, 15, 934–944. [Google Scholar]
- Chantry, C.J.; Frederick, M.M.; Meyer, W.A., III; Handelsman, E.; Rich, K.; Paul, M.E.; Diaz, C.; Cooper, E.R.; Foca, M.; Adeniyi-Jones, S.K.; et al. Endocrine abnormalities and impaired growth in human immunodeficiency virus-infected children. Pediatr. Infect. Dis. J. 2007, 26, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Chantry, C.J.; Hughes, M.D.; Alvero, C.; Cervia, J.S.; Hodge, J.; Borum, P.; Moye, J., Jr. Insulin-like growth factor-1 and lean body mass in HIV-infected children. J. Acquir. Immune Defic. Syndr. 2008, 48, 437–443. [Google Scholar]
- Lochner, M.; Ohnmacht, C.; Presley, L.; Bruhns, P.; Si-Tahar, M.; Sawa, S.; Eberl, G. Microbiota-induced tertiary lymphoid tissues aggravate inflammatory disease in the absence of RORgamma t and LTi cells. J. Exp. Med. 2011, 208, 125–134. [Google Scholar]
- Newell, M.L.; Borja, M.C.; Peckham, C. Height, weight, and growth in children born to mothers with HIV-1 infection in Europe. Pediatrics 2003, 111, e52–e60. [Google Scholar] [CrossRef] [PubMed]
- Pediatric HIV Surveillance (through 2009). Available online: http://www.cdc.gov/hiv/graphics/pediatri.htm (accessed on 15 October 2011).
- Miller, T.L.; Easley, K.A.; Zhang, W.; Orav, E.J.; Bier, D.M.; Luder, E.; Ting, A.; Shearer, W.T.; Vargas, J.H.; Lipshultz, S.E. Maternal and infant factors associated with failure to thrive in children with vertically transmitted human immunodeficiency virus-1 infection: The prospective, P2C2 human immunodeficiency virus multicenter study. Pediatrics 2001, 108, 1287–1296. [Google Scholar]
- Carey, V.J.; Yong, F.H.; Frenkel, L.M.; McKinney, R.M. Growth velocity assessment in paediatric AIDS: Smoothing, penalized quantile regression and the definition of growth failure. Stat. Med. 2004, 23, 509–526. [Google Scholar]
- Cunningham-Rundles, S.; Ahrne, S.; Peoples, J.; Tatad, F.; Mohamed, M.; Nesin, M. Role of Probiotics in the Modulation of Host Immune Response. In Ending the War Metaphor: The Changing Agenda for Unravellng the Host Microbe Relationships Institute of Medicine; National Academy Press: Washington,DC,USA, 2006; pp. 220–229. [Google Scholar]
- Cunningham-Rundles, S.; Nesin, M. Bacterial Infections in the Immunologically Compromised Host. In Persistent Bacterial Infections; Nataro, J., Blaser, M., Cunningham-Rundles, S., Eds.; American Society of Microbiology Press: Washington,DC,USA, 2000; pp. 145–164. [Google Scholar]
- Merlini, E.; Bai, F.; Bellistri, G.M.; Tincati, C.; d’Arminio Monforte, A.; Marchetti, G. Evidence for polymicrobic flora translocating in peripheral blood of HIV-infected patients with poor immune response to antiretroviral therapy. PLoS One 2011, 6. [Google Scholar] [CrossRef]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cunningham-Rundles, S.; Ahrné, S.; Johann-Liang, R.; Abuav, R.; Dunn-Navarra, A.-M.; Grassey, C.; Bengmark, S.; Cervia, J.S. Effect of Probiotic Bacteria on Microbial Host Defense, Growth, and Immune Function in Human Immunodeficiency Virus Type-1 Infection. Nutrients 2011, 3, 1042-1070. https://doi.org/10.3390/nu3121042
Cunningham-Rundles S, Ahrné S, Johann-Liang R, Abuav R, Dunn-Navarra A-M, Grassey C, Bengmark S, Cervia JS. Effect of Probiotic Bacteria on Microbial Host Defense, Growth, and Immune Function in Human Immunodeficiency Virus Type-1 Infection. Nutrients. 2011; 3(12):1042-1070. https://doi.org/10.3390/nu3121042
Chicago/Turabian StyleCunningham-Rundles, Susanna, Siv Ahrné, Rosemary Johann-Liang, Rachel Abuav, Ann-Margaret Dunn-Navarra, Claudia Grassey, Stig Bengmark, and Joseph S. Cervia. 2011. "Effect of Probiotic Bacteria on Microbial Host Defense, Growth, and Immune Function in Human Immunodeficiency Virus Type-1 Infection" Nutrients 3, no. 12: 1042-1070. https://doi.org/10.3390/nu3121042