Effects of Germinated Brown Rice and Its Bioactive Compounds on the Expression of the Peroxisome Proliferator-Activated Receptor Gamma Gene

Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Rice Samples
2.3. Preparation of Extracts
2.4. Cell Culture
2.5. Cell Viability Assay
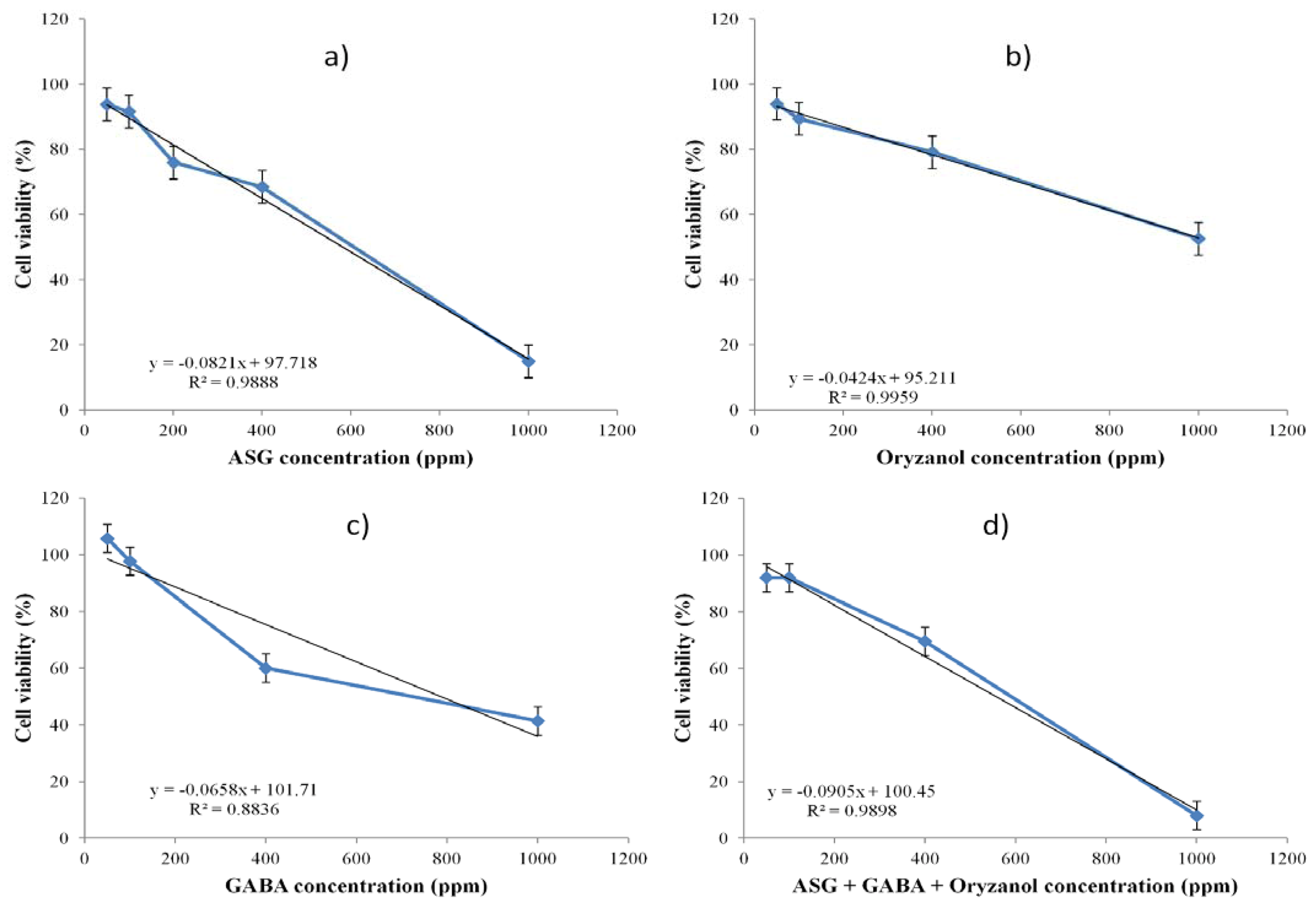
2.6. Treatment of HEP-G2 Cells
2.7. Gene Expression Study

{kind=link}
{kind=link}
| Gene name | Forward primer sequence | Reverse primer sequence |
|---|---|---|
| Insulin Receptor | AGGTGACACTATAGAATAAAGACAGTGAGCTGTTCGAGC | GTACGACTCACTATAGGGAAGTGCCTGAAGAGGTTTTTCTG |
| 3-hydroxy-3-methyl-glutaryl-CoA reductase | AGGTGACACTATAGAATAAATGGCAACAACAGAAGGTTGT | GTACGACTCACTATAGGGAGAAACGGATATAAAGGTTGCGT |
| Peroxisome proliferator-activated receptor gamma | AGGTGACACTATAGAATACAGAAATGACCATGGTTGACA | GTACGACTCACTATAGGGAGGCTCTTCATGAGGCTTATTG |
| Peptidylprolyl isomerase A *,# | AGGTGACACTATAGAATACACACGGCTCACATTGCAT | GTACGACTCACTATAGGGACACGAACAGCAAAGCGA |
| Beta actin * | AGGTGACACTATAGAATAGATCATTGCTCCTCCTGAGC | GTACGACTCACTATAGGGAAAAGCCATGCCAATCTCATC |
| Glyceraldehyde-3-phosphate dehydrogenase * | AGGTGACACTATAGAATAAAGGTGAAGGTCGGAGTCAA | GTACGACTCACTATAGGGAGATCTCGCTCCTGGAAGATG |
2.8. Statistical Analysis
3. Results and Discussion
3.1. Analysis of Extracts
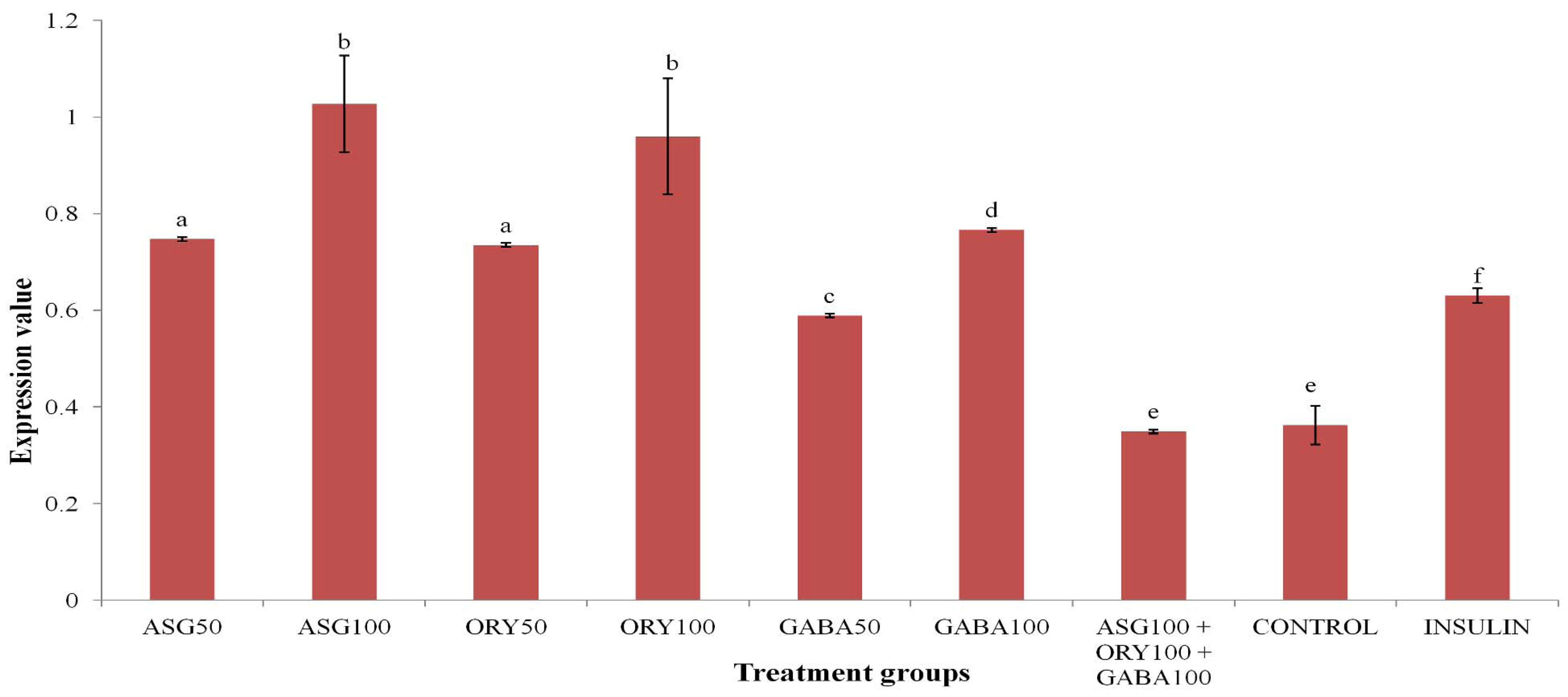
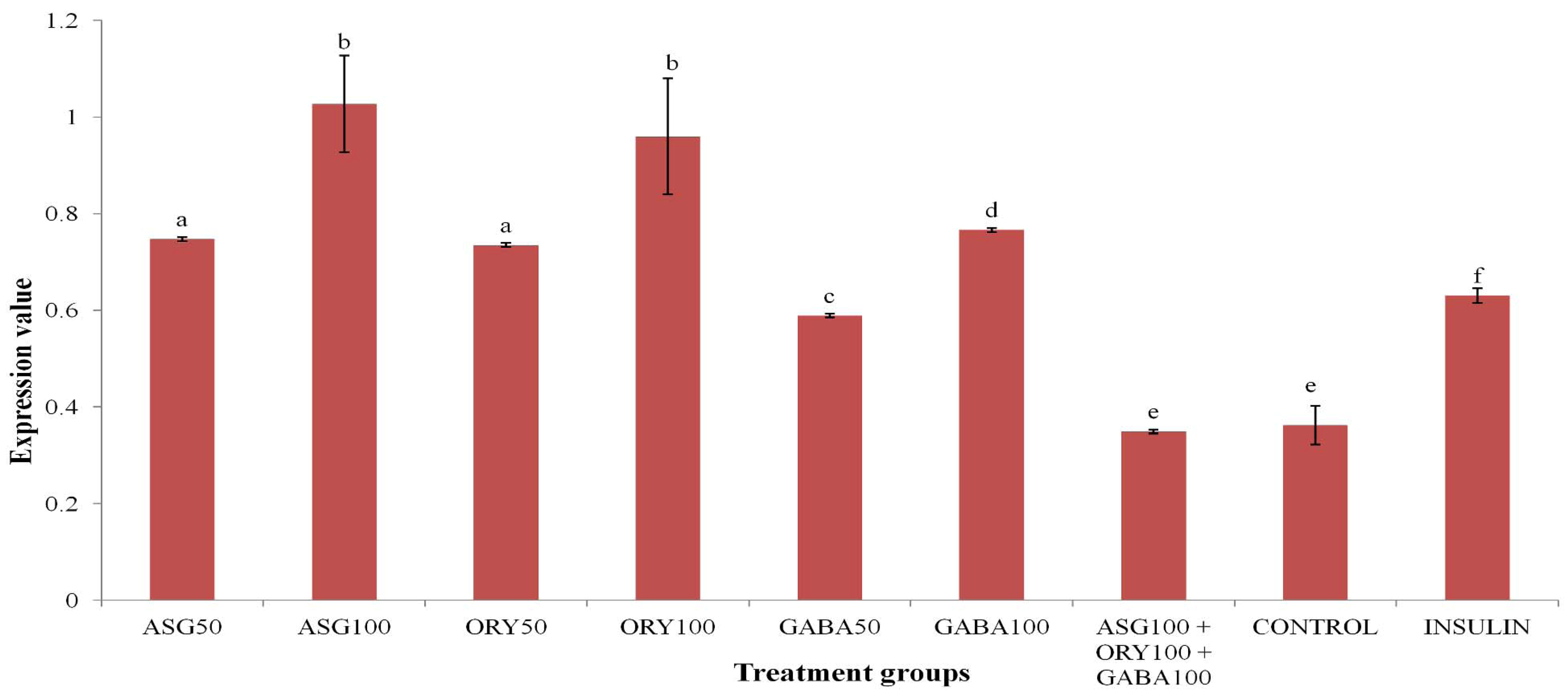
3.2. HEP-G2 PPARγ Gene Expression
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Imam, M.U.; Azmi, N.H.; Bhanger, M.I.; Ismail, N.; Ismail, M. Antidiabetic Properties of Germinated Brown Rice: A Systematic Review. Evid. Based Complement Alternat. Med. 2012, 2012. [Google Scholar] [CrossRef]
- Patil, S.B.; Khan, M.K. Germinated brown rice as a value added rice product: A review. J. Food Sci. Technol. 2011, 48, 661–667. [Google Scholar]
- Jacobs, D.R.; Tapsell, L.C. Food, not nutrients, is the fundamental unit in nutrition. Nutr. Rev. 2007, 65, 439–450. [Google Scholar] [CrossRef]
- Desvergne, B.A.; Wahli, W. Peroxisome Proliferator-Activated Receptors: Nuclear control of metabolism. Endocr. Rev. 1999, 20, 649–688. [Google Scholar]
- Apostoli, A.; Nicol, C.J. PPAR Medicines and Human Disease: The ABCs of it all. PPAR Res. 2012, 2012. [Google Scholar] [CrossRef]
- Lehrke, M.; Lazar, M. The many faces of PPARγ. Cell 2005, 123, 993–999. [Google Scholar]
- Menéndez-Gutiérrez, M.; Roszer, T.; Ricote, M. Biology and therapeutic applications of peroxisome proliferator-activated receptors. Curr. Top. Med. Chem. 2012, 12, 548–584. [Google Scholar]
- Azhar, S. Peroxisome proliferator-activated receptors, metabolic syndrome and cardiovascular disease. Future Cardiol. 2012, 6, 657–691. [Google Scholar]
- Campbell, S.E.; Stone, W.L.; Whaley, S.G.; Qui, M.; Krishnan, K. Gamma (gamma) tocopherol upregulates peroxisome proliferator activated receptor (PPAR) gamma (gamma) expression in SW 480 human colon cancer cell lines. BMC Cancer 2003, 3, 25. [Google Scholar]
- Ho, J.N.; Son, M.E.; Lim, W.C.; Lim, S.T.; Cho, H.Y. Anti-obesity effects of germinated brown rice extract through down-regulation of lipogenic genes in high fat diet-induced obese mice. Biosci. Biotechnol. Biochem. 2012, 76, 1068–1074. [Google Scholar]
- Ortuño Sahagún, D.; Márquez-Aguirre, A.L.; Quintero-Fabián, S.; López-Roa, R.I.; Rojas-Mayorquín, A.E. Modulation of PPAR-γ by Nutraceutics as Complementary Treatment for Obesity-Related Disorders and Inflammatory Diseases. PPAR Res. 2012, 2012. [Google Scholar] [CrossRef]
- Kurtz, T.W. Treating the metabolic syndrome: Telmisartan as a peroxisome proliferator-activated receptor-gamma activator. Acta Diabetol. 2005, 42, S9–S16. [Google Scholar] [CrossRef]
- Sani, I.M.; Iqbal, S.; Chan, K.W.; Ismail, M. Effect of Acid and Base Catalyzed Hydrolysis on the Yield of Phenolics and Antioxidant Activity of Extracts from Germinated Brown Rice (GBR). Molecules 2012, 17, 7584–7594. [Google Scholar]
- Azlan, A.; Ismail, M.; Abdul Hamid, A. Extraction and Determination of Oryzanol in Rice Bran of Mixed Herbarium UKMB: AZ 6807: MR 185, AZ 6808: MR 211, AZ6809: MR 29. ASEAN Food J. 2008, 15, 89–96. [Google Scholar]
- Rozan, P.; Kuo, Y.H.; Lambein, F. Free Amino Acids Present in Commercially Available Seedlings Sold for Human Consumption. A Potential Hazard for Consumers. J. Agric. Food Chem. 2000, 48, 716–723. [Google Scholar]
- Usuki, S.; Ariga, T.; Dasgupta, S.; Kasama, T.; Morikawa, K.; Nonaka, S.; Okuhara, Y.; Kise, M.; Yu, R.K. Structural analysis of novel bioactive acylated steryl glucosides in pre-germinated brown rice. J. Lipid Res. 2008, 49, 2188–2196. [Google Scholar] [CrossRef]
- Phillips, K.; Ruggio, D.; Ashraf-Khorassani, M. Analysis of Steryl Glucosides in Foods and Dietary Supplements by Solid-Phase Extraction and Gas Chromatography. J. Food Lipids 2005, 12, 124–140. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Vacca, M.; Degirolamo, C.; Mariani-Costantini, R.; Palasciano, G.; Moschetta, A. Lipid-sensing nuclear receptors in the pathophysiology and treatment of the metabolic syndrome. Wiley Interdiscip. Rev. Syst. Biol. Med. 2011, 3, 562–587. [Google Scholar] [CrossRef]
- Westerbacka, J.; Cornér, A.; Kannisto, K.; Kolak, M.; Makkonen, J.; Korsheninnikova, E.; Nyman, T.; Hamsten, A.; Fisher, R.; Yki-Järvinen, H. Acute in vivo effects of insulin on gene expression in adipose tissue in insulin-resistant and insulin-sensitive subjects. Diabetologia 2006, 49, 132–140. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Imam, M.U.; Ismail, M.; Ithnin, H.; Tubesha, Z.; Omar, A.R. Effects of Germinated Brown Rice and Its Bioactive Compounds on the Expression of the Peroxisome Proliferator-Activated Receptor Gamma Gene. Nutrients 2013, 5, 468-477. https://doi.org/10.3390/nu5020468
Imam MU, Ismail M, Ithnin H, Tubesha Z, Omar AR. Effects of Germinated Brown Rice and Its Bioactive Compounds on the Expression of the Peroxisome Proliferator-Activated Receptor Gamma Gene. Nutrients. 2013; 5(2):468-477. https://doi.org/10.3390/nu5020468
Chicago/Turabian StyleImam, Mustapha Umar, Maznah Ismail, Hairuszah Ithnin, Zaki Tubesha, and Abdul Rahman Omar. 2013. "Effects of Germinated Brown Rice and Its Bioactive Compounds on the Expression of the Peroxisome Proliferator-Activated Receptor Gamma Gene" Nutrients 5, no. 2: 468-477. https://doi.org/10.3390/nu5020468