The Role of n-3 Polyunsaturated Fatty Acids in the Prevention and Treatment of Breast Cancer


Abstract
:1. Introduction
2. The Effects of n-3 PUFA in Human BC Studies

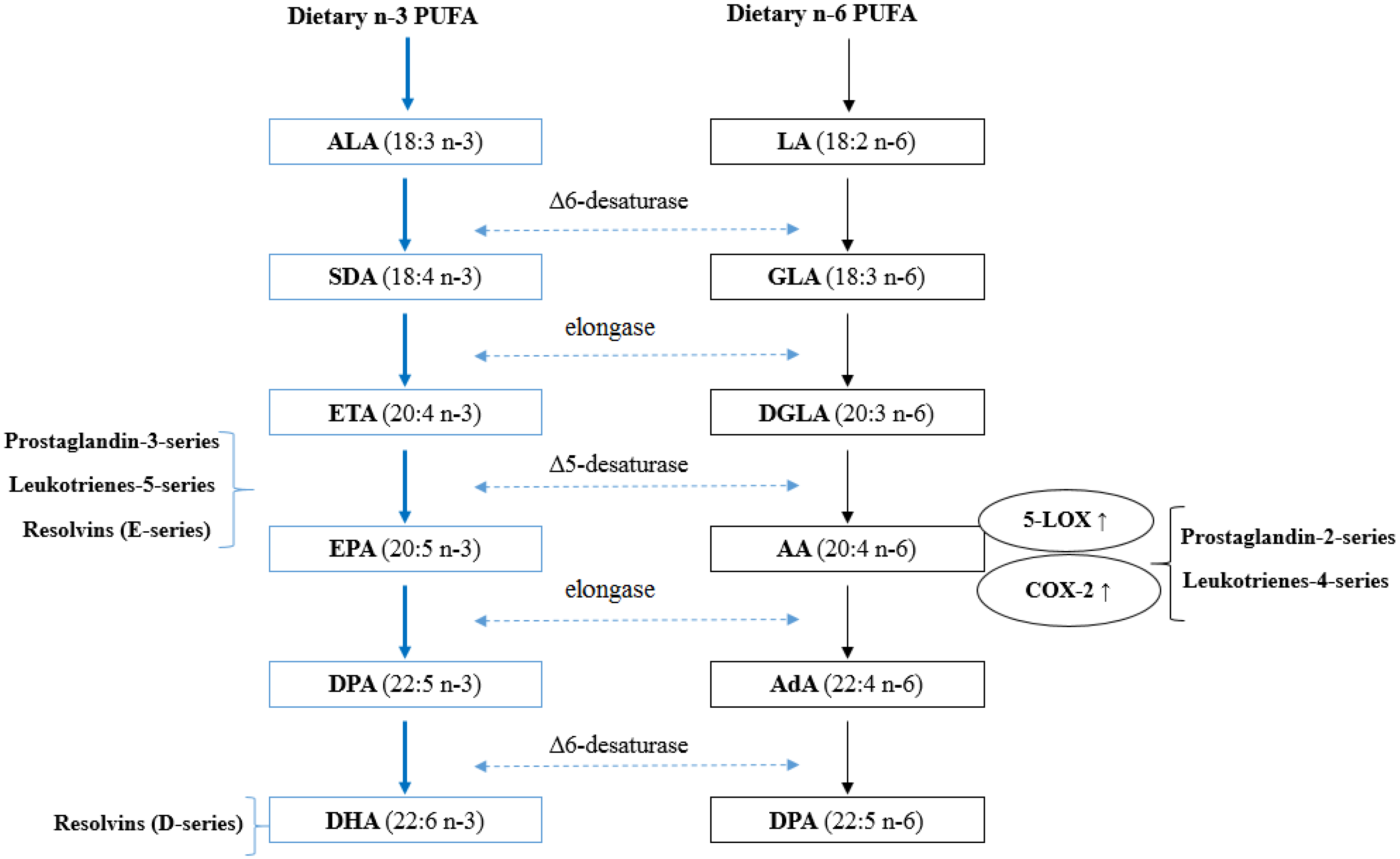{kind=link}
{kind=link}
| Year | Country | Subjects | Method of Assessment | n-3/n-6 PUFA Source | BC Risk | Reference |
|---|---|---|---|---|---|---|
| 2005 | Japan | 26,291 women 40–79 years 129 BC cases | FFQ 1 | Animal and fish fat, vegetable oil, SFA, MUFA and PUFA | ↑ fish fat, EPA + DHA ↓ BC risk | [35] |
| 2003 | Singapore | 35,298 women 45–74 years 342 BC cases | FFQ | Fish/shellfish, saturated, monounsaturated and polyunsaturated fat | ↑ n-3 PUFA from fish/shellfish ↓ BC risk ↑ n-6 PUFA ( low marine n-3) ↑ BC risk | [36] |
| 2010 | US | 35,016 postmenopausal 50–76 years 880 BC cases | FFQ | Dietary fish oil supplement | ↑ fish oil ↓ risk of invasive ductal carcinomas | [40] |
| 2009 | France | 56,007 women 40–65 years 1650 BC case | FFQ | ALA and n-6 PUFA from fruit, nuts and vegetable oils; Long chain n-3 PUFA from meals | no association between total n-3 and BC risk ↑ ALA ↓BC risk ↑ long chain n-3 PUFA ↓ BC risk (at highest quintile of n-6 PUFA) | [42] |
| 2003 | Denmark | 23,693 postmenopausal 50–64 years 424 BC cases | FFQ | Fish | ↑ intake of fish ↑ ER + BC incidence | [43] |
| 2011 | China | 72,571 women 40–70 years 712 BC cases | FFQ | Fish, marine-derived n-3 PUFA red meat | ↑ n-6/n-3 PUFA ratio ↑ BC risk | [37] |
| Year | Country | Subjects Characteristics | Method of Assessment | n-3/n-6 PUFA Source | BC Risk | Reference |
|---|---|---|---|---|---|---|
| 2007 | Japan | 103 incident BC cases 309 controls | erythrocyte membrane FFQ | dietary food intake including soy and meat products, fish and other seafood, vegetables | ↑ dietary intake of n-3 fatty acids ↓ BC risk ↑ long chain n-3 PUFA in erythrocyte ↓ BC risk ↑ saturated fatty ↑ BC risk | [46] |
| 2007 | China | 322 incident BC cases 1030 controls | erythrocyte membrane | ↑ total n-3 fatty acids and EPA ↓ BC risk | [47] | |
| 2009 | China | 155 NPFC 1 185 PFC 2 241 BC, 1030 controls | erythrocyte membrane FFQ | dietary food intake | ↑ EPA ↓ risk of NPFC ↓ progression of PFC to BC ↑ γ-linolenic acid ↑ risk of NPFC, PFC and BC | [38] |
| 2002 | US | 73 BC patients 74 controls | breast adipose tissue | ↑ EPA and DHA ↓ n-6/n-3 PUFA ratio ↓ BC risk ↑ n-6 PUFA ↑ BC risk | [18] | |
| 2003 | US | 565 incident BC 554 controls | FFQ | daily fat intake | ↓ n-6/n-3 PUFA ratio ↓ BC risk (premenopausal) ↑EPA, DHA ↓ BC risk (21% and 18%, respectively) | [41] |
| 2009 | Denmark | 463 BC cases 1098 controls | Gluteal adipose tissue biopsy | dietary food intake | No association between total or individual marine n-3 PUFA in adipose tissue and risk of BC | [44] |
| 2012 | Mexican | 1000 incident BC cases 1074 controls | Interview and FFQ | dietary food intake | ↑ n-3 PUFA ↓ BC risk (obese women) ↑ n-6 PUFA ↑ BC risk (premenopausal) | [19] |
| 2009 | South Korea | 358 incident BC patients 360 controls | FFQ | fatty and lean fish | ↑ fatty fish consumption ↓ BC risk ↑EPA and DHA derived from fish ↓ BC risk | [39] |
3. PUFA—Potential Mechanisms of Action
3.1. Influence on Cell Plasma Membrane Composition
3.2. Inhibition of Arachidonic Acid (AA) Derived Eicosanoid Biosynthesis

3.3. Influence on Gene Expression, Transcription Factor Activity and Signal Transduction

3.3.1. EGFR and HER-2
3.3.2. Peroxisome Proliferator-Activated Receptor Gamma (PPARγ)
3.3.3. Bax/Bcl-2
3.3.4. PI3K/Akt, NF-κB
3.3.5. Cell Proliferation Marker: Ki-67 and PCNA
4. The Effect of n-3 PUFA Mixtures on BC Development
4.1. Animal Studies
4.1.1. Breast Cancer Studies in Xenograft Rodent Models
4.1.2. Breast Cancer Studies in Transgenic Rodent Models
| Animal Model | n-3 PUFA Source | Feeding Period | Main Findings | Mechanism | Reference |
|---|---|---|---|---|---|
| Athymic nu/nu mice MDA-MB 231 | 3% w/w fish oil concentrate (10.2 g/kg EPA, 7.2 g/kg DHA, 3.0 g/kg ALA) | 7-week (fed after tumor established) | ↓ tumor growth rate ↑ effectiveness of doxorubucin | ↑ EPA incorporation into tumor ↑ lipid peroxidation in tumor | [30] |
| Athymic nu/nu mice (NCr-nu/nu) MDA-MB 435 | 40 or 80 g/kg EPA, DHA | 13-week (fed before transplantation) | ↓ tumor growth, size ↓ tumor weight | ↑ EPA, DHA in tumor phospholipids ↓ LA, AA in tumor phospholipids ↓ AA-derived eicosanoids | [20] |
| Inbred F44 rats R3230AC | 5% marine oil supplementation (18% EPA, 12% DHA) | 4-week (fed before transplantation) | ↓ tumor weight, volume | ↑ EPA, DHA, AA incorporation into tumor ↓ Prostaglandins 2 series | [112] |
| BALB/cAnN mice Mouse BC cell | 10% or 20% w/w menhaden fish oil | 7-week (fed before transplantation) | ↑ tumor latency ↓ tumor growth rate | NA | [114] |
| Athymic nude mice MCF-7 | 19% w/w menhaden oil (1.9 g/kg ALA, 19.4 g/kg EPA, 24.3 g/kg DHA) | 6 or 8-week (fed after tumor established) | ↓ tumor volume | ↑ lipid peroxidation in tumor | [113] |
| Animal Model | n-3 PUFA Source | Feeding Period | Main Findings | Mechanism | Reference |
|---|---|---|---|---|---|
| MMTV-HER-2/neu | 22.50 kcal% menhaden oil (15 g/kg EPA, 10.8 g/kg DHA) | 28-week (fed before tumor development) | ↓ atypical ductal hyperplasia ↓ cell proliferation prevented HER-2/neu at early stages | ↓ Ki-67 expression ↓ COX-2 expression | [77] |
| MMTV-HER-2/neu | 22.50 kcal% menhaden oil (15 g/kg EPA, 10.8 g/kg DHA) | 52-week (fed before tumor development) | ↓ tumor incidence and multiplicity ↑ tumor latency ↓ mammary gland dysplasia | NA | [115] |
| MMTV-neu (ndl)-YD5 × fat1 | 3% w/w menhaden oil (0.5 g/kg ALA, 4.1 g/kg EPA, 3 g/kg DHA) | 20-week (lifelong treatment, fed before tumor development) | ↓ tumor volume and multiplicity | ↑ EPA, DHA and overall n-3 in mammary tissues ↓ n-6/n-3 ratio in tumor phospholipids | [116] |
| MMTV-neu (ndl)-YD5 | 3% w/w menhaden oil (0.5 g/kg ALA, 4.1 g/kg EPA, 3 g/kg DHA) 9% w/w menhaden oil (1.3 g/kg ALA, 12.4 g/kg EPA, 9 g/kg DHA) | 20-week (lifelong treatment, fed before tumor development) | ↓ tumor volume and multiplicity ↑ tumor latency (all in a dose-dependent manner) | ↑ EPA, DPA in mammary tissues ↑ EPA, DHA in tumor phospholipids ↓ LA, AA, n-6/n-3 PUFA ratio in both mammary and tumor tissues in a dose-dependent manner | [21] |
4.1.3. Breast Cancer Studies in Chemically-Induced Rodent Models
| Carcinogen | n-3 PUFA Source | Feeding Period | Main Findings | Mechanism | Reference |
|---|---|---|---|---|---|
| MNU | Fish oil 2%–10% w/w n-3 PUFA in diet | 18-week (at the same time as MNU administration) | Absolute n-3 diet: ↓ body weight, no tumor occurrence (10% w/w n-3 PUFA) 1:1n-6/n-3 diet ↓ tumor incidence and multiplicity (5% w/w n-3 PUFA) | ↑ EPA, DHA in mammary ↓ FAS, COX-2, 5-LOX | [64] |
| MNU | Fish oil concentrate Low n-6/n-3 = 1:14.6 High n-6/n-3 = 1:0.7 | 2-week (at the same time as MNU administration) | Low vs. high ratio n-6/n-3 PUFA diet: ↓ tumor incidence (21%), ↓ tumor multiplicity (30%), tumor burden (80%) ↑ apoptotic index (129%) | ↓ Ki-67 ↑ Bax, Bax/Bcl2, PPARγ ↓ NF-κB p65, pAkt, IGF-IR | [87] |
| MNU | EPA/DHA alone: 95 g/kg EPA/DHA EPA + DHA: 47.5 g/kg EPA + 47.5 g/kg DHA | 20-week (at the same time as MNU administration) | DHA alone vs EPA + DHA vs EPA alone: ↓ tumor incidence: 23%, 73%, 65% ↓ tumor multiplicity: 0.23, 1.67, 1.59 DHA is more effectively than EPA | NA | [120] |
| DMBA | Maxepa (fish oil concentrate): 90 mg EPA + 60 mg DHA per day | 24-week study 35-week study (before DMBA injection) | ↓ DNA single-strand breaks ↓ cell proliferation | ↓ Ki-67, Her-2/neu | [26] |
| DMBA | Maxepa: 90 mg EPA + 60 mg DHA per day | 24-week study 35-week study (before DMBA injection) | ↓ tumor incidence (23%), tumor multiplicity (42%) ↑ cell apoptosis ↓cell proliferation | ↓ Bcl-2 ↑Bax ↑ p53 | [27,28] |
| DMBA | Fish oil (0.5%ALA, 16% EPA, 1.2% DPA, 8% DHA in fish oil) | NA | ↓ tumor incidence with fish oil consumption: adulthood < in utero < puberty < perinatal ↓ tumor multiplicity with fish oil consumption: adulthood > puberty > perinatal > in utero | ↓ maternal serum estradiol | [29] |
| DMBA | Menhaden oil Low-fat n-3 PUFA diet: 4.6 g/kg EPA + 3.2 g/kg DHA High fat n-3 PUFA diet: 9.1 g/kg EPA + 6.3 g/kg DHA | 20-day (before DMBA injection) | Low n-3 diet: ↓ tumor incidence ↓ TEBs ↓ cell proliferation ↑ cell apoptosis; High n-3 diets exert opposite effects | Low n-3 diet: ↓ COX-2, PCNA ↑ PPARγ ↑ lipid peroxidation High n-3 diet: ↑ pAkt ↑ lipid peroxidation | [110] |
4.2. Cell Culture Studies
| Cell Type | n-3 PUFA Source | Main Finding | Mechanism | Reference |
|---|---|---|---|---|
| MDA-MB-231 | EPA/DHA alone: 75 μM or 100 μM EPA + DHA combination: 45 μM EPA + 30 μM DHA or 60 μM EPA + 40 μM DHA (in presence/absence of LA) | ↓ cell viability, cell proliferation ↑ DNA fragmentation, cell apoptosis DHA was more potent than EPA | ↓ pAkt ↓ NF-κB and DNA binding activity | [96] |
| MDA-MB-231 | 0.5–2.5 μg/mL of EPA, DHA (1.7–8.2 μM EPA, 1.5–7.6 μMDHA) | ↓ tumor cells growth (DHA > EPA, dose-dependent) | ↓ LA composition in cell lipids ↓ AA-derived eicosanoid synthesis | [121] |
| MDA-MB-231 | EPA/DHA alone: 75 μM or 100 μM EPA + DHA combination: 45 μM EPA + 30 μM DHA or 60 μM EPA + 40 μM DHA (in presence/absence of LA) | ↓cell growth (48%–62%) | ↑ EPA, DHA, DPA and total n-3 in lipid rafts ↓ EGFR levels ↑ pEGFR | [22] |
| MDA-MB-231 MCF-7 | EPA (230 μM), DHA (200 μM) | ↓ cell viability ↑ cell apoptosis | ↓ Bcl-2 ↑pro-caspase-8 ↓ pEGFR ↓ EGFR (only DHA) ↓ AA ↑ EPA, DPA, DHA in total cell lipids | [71] |
| MDA-MB-231 MCF-7 | 3–100 μM of EPA, DHA | At 50 μM EPA, 30 μM DHA ↑ cell apoptosis ↓ cell growth At 50 μM EPA, DHA ↑ G2/M duration DHA was more potent than EPA | ↓ phosphorylation of cyclin B1 ↓ activity of CDK1-cyclin B1 | [86] |
| MCF-7 | 100 μM of EPA, DHA | ↓ cell growth (30% by EPA, 54% by DHA) ↑ cell differentiation (30% by EPA, 65% by DHA) No significant effects on cell apoptosis and cell cycle DHA was more potent than EPA | ↑ PPARγ (DHA only) | [125] |
| MCF-7 MCF-10A | 6–30 μM of ALA, EPA, DHA | All n-3 PUFA ↓ MCF-7 cell growth (EPA, DHA > ALA, dose-dependent) AA ↓ MCF-7 cell growth (similar as ALA) | NA | [122] |
| ER+ and ER− cells | 20 μg/mL of ALA, EPA, DHA (72 μM ALA, 66 μM EPA, 61 μM DHA) | EPA, DHA ↓ cell proliferation (all cell lines) ALA ↓ estrogen independent BC cell proliferation | ↑ lipid peroxidation | [124] |
5. The Effect of Individual n-3 PUFA on BC Development
5.1. ALA and BC
5.1.1. Inefficient Conversion from ALA to EPA and DHA
5.1.2. Individual Effect of ALA on Breast Cancer
| n-3 PUFA | Amount of Fatty Acid | Effect | Mechanism | Reference |
|---|---|---|---|---|
| ALA | NA | Moderate decrease BC risk | NA | [45] |
| ~22.8 g of ALA per kg diet | Reduced tumor cell proliferation | Inhibited HER2, EGFR expression | [78] | |
| ~22.8 g of ALA per kg diet | Inhibited MCF-7 cell proliferation | [78] | ||
| ~11 g ALA per kg diet | Reduced tumor incidence and burden | Increased BAX/Bcl-2 ratio | [93] | |
| 10.6 g ALA per kg diet | Decreased tumor growth rate | Inhibited HER2 expression | [105] | |
| 72 μM ALA | Moderate inhibited ER-negative cell proliferation, not affect MCF-7 | NA | [124] | |
| 30 μM of ALA | Slightly inhibited MCF-7 | NA | [122] | |
| NA | Inversely associated with BC risk | NA | [133] | |
| NA | Inversely correlated with metastasis development | NA | [134] | |
| 55.9 g ALA per kg diet | Reduced tumor growth and metastasis | NA | [135] | |
| 8 g ALA per kg diet | Decreased tumor growth rate | NA | [136] | |
| 10 g ALA per kg diet | Reduced tumor burden and increased survival rate | NA | [137] | |
| 2.5-40 μM of ALA | enhanced cytotoxic effects of Trastuzumab (at 10 μM of ALA) | Down-regulated HER2 (at 20 μM of ALA) | [138] | |
| 10 μM of ALA | Diminished proteolytic cleavage of the extracellular domain of HER2 | Inhibited HER-2 activity | [139] | |
| ~21.2 g of ALA per kg diet | Minimal inhibited tumor growth w/wo Trastuzumab | NA | [140] | |
| 52.8 g of ALA per kg diet | Inhibited mammary tumor development | NA | [141] | |
| EPA | 40–80 g of EPA per kg diet | Slowed down tumor growth, reduced tumor burden | Decreased AA derived-eicosanoid | [20] |
| 3–100 μM of EPA | Induced BC cell apoptosis (at 50 μM of EPA) | NA | [86] | |
| 40–200 μM of EPA | Restored the growth inhibitory effect of Tamoxifen (at 40 μM of EPA) | Decreased pAkt (at 20 μM of EPA) | [97] | |
| 20–80 g of EPA per kg diet | Inhibited the development of lung metastasis | NA | [126] | |
| 100 μM of EPA | Inhibited MCF-7 cell growth | NA | [125] | |
| 40 μM of EPA | Induced apoptosis, inhibited cell proliferation, arrested cell cycle at G0/G1 | down-regulated Bcl-2 expression | [142] | |
| 95 g of EPA per kg diet | Reduced KPL-1 cell proliferation rate and metastasis | NA | [143] | |
| 42 g of EPA per kg diet | Suppressed cell proliferation in MCF-7 xenografts in rats | NA | [144] | |
| 50 μM of EPA | Increased PPARγ at mRNA level | NA | [145] | |
| 0–200 μM of EPA | Inhibited MCF-7 cell growth (at 60 μM of EPA) | NA | [146] | |
| DHA | 120 μM of DHA | Decreased cancer cell viability, enhanced the cytotoxic activity of taxanes | Decreased the expression of Her-2/neu | [5] |
| 100 μM of DHA | Disrupted lipid rafts, induced apoptosis in HER-2 overexpressing cells | Decreased Akt activity and FAN | [6] | |
| 100 μM of DHA | Decreased MDA-MB-231 cell proliferation, enhanced EGFR inhibitors | Altered EGFR phosphorylation and localization | [56] | |
| 0–200 μM of DHA | Reduced MCF-7 cell viability and DNA synthesis (at 25 μM of DHA) | Increased lipid peroxidation, capase 8 activation | [146] | |
| 20 or 100 μM of DHA | Inhibited MDA-MB-231 cell proliferation, promoted nuclear condensation | Increased caspase-3 activity (at 100 μM of DHA) | [147] | |
| 10–160 μM of DHA | Inhibited MCF-7 cell growth and induced apoptosis (at 40 μM of DHA) | Downregulated Bcl-2, increased Bax/Bcl-2 ratio | [148] | |
| 270 μM of DHA | 50% inhibitory KPL-1 cell growth after 72 h treatment | Downregulated Bcl-2, increased Bax/Bcl-2 ratio | [149] | |
| 40 g of DHA per kg diet | Decreased tumor growth rate and final tumor weight, increased apoptosis | Reduced tumor PGE2, decreased Ki-67 | [150] | |
| 32 g of DHA per kg diet | Reduced tumor incidence | Increased BRCA1 at protein level | [151] | |
| 30 μM of DHA | 50% inhibitory MCF7 cell growth after 96 h treatment | Increased BRCA1/2 at transcriptional level | [152] | |
| NA | Increased response of the tumor to chemotherapies, increased survival rate | [153] |
| BC Cell Type | ALA | EPA | DHA |
|---|---|---|---|
| MDA-MB-231 (ER−) | ✓ | ✓ | ✓ |
| MDA-MB 435 (ER−) | NA | ✓ | ✓ |
| MCF-10A (ER−) | — | ✓ | ✓ |
| HBL-100 (ER−) | ✓ | ✓ | ✓ |
| MCF-7 (ER+) | — | ✓ | ✓ |
| ZR-75 (ER+) | — | ✓ | ✓ |
| T-47-D (ER+) | — | ✓ | ✓ |
| SK-Br3 and BT-474 (HER-2/neu positive) | ✓ | NA | ✓ |
5.2. Individual Effect of EPA on Breast Cancer
5.3. Individual Effect of DHA on Breast Cancer
6. Plant-Derived n-3 (ALA) vs. Marine-Based n-3 (EPA, DHA)
7. Conclusions
Acknowledgments
Conflict of Interest
References
- DeSantis, C.; Ma, J.; Bryan, L.; Jemal, A. Breast cancer statistics, 2013. CA Cancer J. Clin. 2014, 64, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Canadian Cancer Statistics. Canadian Cancer Society’s Advisory; Canadian Cancer statistics: Toronto, ON, Canada, 2013. [Google Scholar]
- Gerber, M. Omega-3 fatty acids and cancers: A systematic update review of epidemiological studies. Br. J. Nutr. 2012, 107 (Suppl. 2), S228–S239. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, M.; Tsugane, S. Risk factors for breast cancer: Epidemiological evidence from Japanese studies. Cancer Sci. 2011, 102, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Menendez, J.A.; Lupu, R.; Colomer, R. Exogenous supplementation with omega-3 polyunsaturated fatty acid docosahexaenoic acid (DHA; 22:6n-3) synergistically enhances taxane cytotoxicity and downregulates Her-2/neu (c-erbB-2) oncogene expression in human breast cancer cells. Eur. J. Cancer Prev. 2005, 14, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Ravacci, G.R.; Brentani, M.M.; Tortelli, T., Jr.; Torrinhas, R.S.; Saldanha, T.; Torres, E.A.; Waitzberg, D.L. Lipid raft disruption by docosahexaenoic acid induces apoptosis in transformed human mammary luminal epithelial cells harboring HER-2 overexpression. J. Nutr. Biochem. 2013, 24, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.S.; Hu, X.J.; Zhao, Y.M.; Yang, J.; Li, D. Intake of fish and marine n-3 polyunsaturated fatty acids and risk of breast cancer: Meta-analysis of data from 21 independent prospective cohort studies. BMJ 2013, 346. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Saadatian-Elahi, M.; Norat, T.; Goudable, J.; Riboli, E. Biomarkers of dietary fatty acid intake and the risk of breast cancer: A meta-analysis. Int. J. Cancer 2004, 111, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Boyd, N.F.; Stone, J.; Vogt, K.N.; Connelly, B.S.; Martin, L.J.; Minkin, S. Dietary fat and breast cancer risk revisited: A meta-analysis of the published literature. Br. J. Cancer 2003, 89, 1672–1685. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.M.; Ma, D.W. Are all n-3 polyunsaturated fatty acids created equal? Lipids Health Dis. 2009, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Bidu, C.; Bellenger, S.; Narce, M.; Bellenger, J. n-3 polyunsaturated fatty acids and HER2-positive breast cancer: Interest of the fat-1 transgenic mouse model over conventional dietary supplementation. Biochimie 2014, 96, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Murray, M. ω-3 Polyunsaturated fatty acids and their metabolites as inhibitors of mammalian tumorigenesis. Phytochem. Rev. 2014, 13, 139–156. [Google Scholar] [CrossRef]
- Akhtar, S.; Ismail, T.; Riaz, M. Flaxseed—A miraculous defense against some critical maladies. Pak. J. Pharm. Sci. 2013, 26, 199–208. [Google Scholar] [PubMed]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine of the National Academies. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; The National Academies Press: Washington, DC, USA, 2005. [Google Scholar]
- Vannice, G.; Rasmussen, H. Position of the academy of nutrition and dietetics: Dietary fatty acids for healthy adults. J. Acad. Nutr. Diet. 2014, 114, 136–153. [Google Scholar] [CrossRef] [PubMed]
- Bagga, D.; Anders, K.H.; Wang, H.J.; Glaspy, J.A. Long-chain n-3-to-n-6 polyunsaturated fatty acid ratios in breast adipose tissue from women with and without breast cancer. Nutr. Cancer 2002, 42, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Chajes, V.; Torres-Mejia, G.; Biessy, C.; Ortega-Olvera, C.; ngeles-Llerenas, A.; Ferrari, P.; Lazcano-Ponce, E.; Romieu, I. omega-3 and omega-6 Polyunsaturated fatty acid intakes and the risk of breast cancer in Mexican women: Impact of obesity status. Cancer Epidemiol. Biomark. Prev. 2012, 21, 319–326. [Google Scholar] [CrossRef]
- Rose, D.P.; Connolly, J.M.; Rayburn, J.; Coleman, M. Influence of diets containing eicosapentaenoic or docosahexaenoic acid on growth and metastasis of breast cancer cells in nude mice. J. Natl. Cancer Inst. 1995, 87, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Leslie, M.A.; Abdelmagid, S.A.; Perez, K.; Muller, W.J.; Ma, D.W. Mammary tumour development is dose-dependently inhibited by n-3 polyunsaturated fatty acids in the MMTV-neu(ndl)-YD5 transgenic mouse model. Lipids Health Dis. 2014, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Schley, P.D.; Brindley, D.N.; Field, C.J. (n-3) PUFA alter raft lipid composition and decrease epidermal growth factor receptor levels in lipid rafts of human breast cancer cells. J. Nutr. 2007, 137, 548–553. [Google Scholar] [PubMed]
- Silva, V.; Barazzoni, R.; Singer, P. Biomarkers of fish oil omega-3 polyunsaturated fatty acids intake in humans. Nutr. Clin. Pract. 2014, 29, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, R.G.; Hoover, R.N.; Pike, M.C.; Hildesheim, A.; Nomura, A.M.; West, D.W.; Wu-Williams, A.H.; Kolonel, L.N.; Horn-Ross, P.L.; Rosenthal, J.F.; et al. Migration patterns and breast cancer risk in Asian-American women. J. Natl. Cancer Inst. 1993, 85, 1819–1827. [Google Scholar] [CrossRef]
- Singh, G.K.; Hiatt, R.A. Trends and disparities in socioeconomic and behavioural characteristics, life expectancy, and cause-specific mortality of native-born and foreign-born populations in the United States, 1979–2003. Int. J. Epidemiol. 2006, 35, 903–919. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.; Janarthan, M.; Ghosh, B.; Rana, B.; Rana, A.; Chatterjee, M. Fish oil regulates cell proliferation, protect DNA damages and decrease HER-2/neu and c-Myc protein expression in rat mammary carcinogenesis. Clin. Nutr. 2010, 29, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.; Chakraborty, T.; Ghosh, B.; Chatterjee, M.; Panda, A.; Srivastava, S.; Rana, A.; Chatterjee, M. Dietary fish oil associated with increased apoptosis and modulated expression of Bax and Bcl-2 during 7,12-dimethylbenz(α)anthracene-induced mammary carcinogenesis in rats. Prostaglandins Leukot. Essent. Fatty Acids 2008, 79, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.; Chakraborty, T.; Damodaran, S.; Samanta, K.; Rana, B.; Chatterjee, M. Protective role of fish oil (Maxepa) on early events of rat mammary carcinogenesis by modulation of DNA-protein crosslinks, cell proliferation and p53 expression. Cancer Cell Int. 2007, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Su, H.M.; Hsieh, P.H.; Chen, H.F. A maternal high n-6 fat diet with fish oil supplementation during pregnancy and lactation in rats decreases breast cancer risk in the female offspring. J. Nutr. Biochem. 2010, 21, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Hardman, W.E.; Avula, C.P.; Fernandes, G.; Cameron, I.L. Three percent dietary fish oil concentrate increased efficacy of doxorubicin against MDA-MB 231 breast cancer xenografts. Clin. Cancer Res. 2001, 7, 2041–2049. [Google Scholar] [PubMed]
- Larsson, S.C.; Kumlin, M.; Ingelman-Sundberg, M.; Wolk, A. Dietary long-chain n-3 fatty acids for the prevention of cancer: A review of potential mechanisms. Am. J. Clin. Nutr. 2004, 79, 935–945. [Google Scholar] [PubMed]
- Terry, P.D.; Rohan, T.E.; Wolk, A. Intakes of fish and marine fatty acids and the risks of cancers of the breast and prostate and of other hormone-related cancers: A review of the epidemiologic evidence. Am. J. Clin. Nutr. 2003, 77, 532–543. [Google Scholar] [PubMed]
- Stoll, B.A. n-3 fatty acids and lipid peroxidation in breast cancer inhibition. Br. J. Nutr. 2002, 87, 193–198. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, M.; Ma, D.W. Role of dietary fatty acids in mammary gland development and breast cancer. Breast Cancer Res. 2010, 12, 211. [Google Scholar] [CrossRef] [PubMed]
- Wakai, K.; Tamakoshi, K.; Date, C.; Fukui, M.; Suzuki, S.; Lin, Y.; Niwa, Y.; Nishio, K.; Yatsuya, H.; Kondo, T.; et al. Dietary intakes of fat and fatty acids and risk of breast cancer: A prospective study in Japan. Cancer Sci. 2005, 96, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Gago-Dominguez, M.; Yuan, J.M.; Sun, C.L.; Lee, H.P.; Yu, M.C. Opposing effects of dietary n-3 and n-6 fatty acids on mammary carcinogenesis: The Singapore Chinese Health Study. Br. J. Cancer 2003, 89, 1686–1692. [Google Scholar] [CrossRef] [PubMed]
- Murff, H.J.; Shu, X.O.; Li, H.; Yang, G.; Wu, X.; Cai, H.; Wen, W.; Gao, Y.T.; Zheng, W. Dietary polyunsaturated fatty acids and breast cancer risk in Chinese women: A prospective cohort study. Int. J. Cancer 2011, 128, 1434–1441. [Google Scholar] [CrossRef] [PubMed]
- Shannon, J.; King, I.B.; Lampe, J.W.; Gao, D.L.; Ray, R.M.; Lin, M.G.; Stalsberg, H.; Thomas, D.B. Erythrocyte fatty acids and risk of proliferative and nonproliferative fibrocystic disease in women in Shanghai, China. Am. J. Clin. Nutr. 2009, 89, 265–276. [Google Scholar] [CrossRef]
- Kim, J.; Lim, S.Y.; Shin, A.; Sung, M.K.; Ro, J.; Kang, H.S.; Lee, K.S.; Kim, S.W.; Lee, E.S. Fatty fish and fish omega-3 fatty acid intakes decrease the breast cancer risk: A case-control study. BMC Cancer 2009, 9, 216. [Google Scholar] [CrossRef] [PubMed]
- Brasky, T.M.; Lampe, J.W.; Potter, J.D.; Patterson, R.E.; White, E. Specialty supplements and breast cancer risk in the VITamins And Lifestyle (VITAL) Cohort. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1696–1708. [Google Scholar] [CrossRef]
- Goodstine, S.L.; Zheng, T.; Holford, T.R.; Ward, B.A.; Carter, D.; Owens, P.H.; Mayne, S.T. Dietary (n-3)/(n-6) fatty acid ratio: Possible relationship to premenopausal but not postmenopausal breast cancer risk in U.S. women. J. Nutr. 2003, 133, 1409–1414. [Google Scholar] [PubMed]
- Thiebaut, A.C.; Chajes, V.; Gerber, M.; Boutron-Ruault, M.C.; Joulin, V.; Lenoir, G.; Berrino, F.; Riboli, E.; Benichou, J.; Clavel-Chapelon, F. Dietary intakes of omega-6 and omega-3 polyunsaturated fatty acids and the risk of breast cancer. Int. J. Cancer 2009, 124, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Stripp, C.; Overvad, K.; Christensen, J.; Thomsen, B.L.; Olsen, A.; Moller, S.; Tjonneland, A. Fish intake is positively associated with breast cancer incidence rate. J. Nutr. 2003, 133, 3664–3669. [Google Scholar] [PubMed]
- Witt, P.M.; Christensen, J.H.; Schmidt, E.B.; Dethlefsen, C.; Tjonneland, A.; Overvad, K.; Ewertz, M. Marine n-3 polyunsaturated fatty acids in adipose tissue and breast cancer risk: A case-cohort study from Denmark. Cancer Causes Control 2009, 20, 1715–1721. [Google Scholar] [CrossRef] [PubMed]
- Maillard, V.; Bougnoux, P.; Ferrari, P.; Jourdan, M.L.; Pinault, M.; Lavillonniere, F.; Body, G.; Le, F.O.; Chajes, V. n-3 and n-6 fatty acids in breast adipose tissue and relative risk of breast cancer in a case-control study in Tours, France. Int. J. Cancer 2002, 98, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Kuriki, K.; Hirose, K.; Wakai, K.; Matsuo, K.; Ito, H.; Suzuki, T.; Hiraki, A.; Saito, T.; Iwata, H.; Tatematsu, M.; et al. Breast cancer risk and erythrocyte compositions of n-3 highly unsaturated fatty acids in Japanese. Int. J. Cancer 2007, 121, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Shannon, J.; King, I.B.; Moshofsky, R.; Lampe, J.W.; Gao, D.L.; Ray, R.M.; Thomas, D.B. Erythrocyte fatty acids and breast cancer risk: A case-control study in Shanghai, China. Am. J. Clin. Nutr. 2007, 85, 1090–1097. [Google Scholar] [PubMed]
- Patterson, R.E.; Flatt, S.W.; Newman, V.A.; Natarajan, L.; Rock, C.L.; Thomson, C.A.; Caan, B.J.; Parker, B.A.; Pierce, J.P. Marine fatty acid intake is associated with breast cancer prognosis. J. Nutr. 2011, 141, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Signori, C.; DuBrock, C.; Richie, J.P.; Prokopczyk, B.; Demers, L.M.; Hamilton, C.; Hartman, T.J.; Liao, J.; El-Bayoumy, K.; Manni, A. Administration of omega-3 fatty acids and Raloxifene to women at high risk of breast cancer: Interim feasibility and biomarkers analysis from a clinical trial. Eur. J. Clin. Nutr. 2012, 66, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.U.; Chen, J.M.; Li, T.; Strasser-Weippl, K.; Goss, P.E. Dietary flaxseed alters tumor biological markers in postmenopausal breast cancer. Clin. Cancer Res. 2005, 11, 3828–3835. [Google Scholar] [CrossRef] [PubMed]
- Menendez, J.A.; Lupu, R. Fatty acid synthase and the lipogenic phenotype in cancer pathogenesis. Nat. Rev. Cancer 2007, 7, 763–777. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Suburu, J.; Chen, H.; Chen, Y.Q. Mechanisms of omega-3 polyunsaturated fatty acids in prostate cancer prevention. Biomed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Wassall, S.R.; Stillwell, W. Docosahexaenoic acid domains: The ultimate non-raft membrane domain. Chem. Phys. Lipids 2008, 153, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Chenais, B.; Blanckaert, V. The Janus face of lipids in human breast cancer: How polyunsaturated Fatty acids affect tumor cell hallmarks. Int. J. Breast Cancer 2012, 2012. [Google Scholar] [CrossRef]
- Murai, T. The role of lipid rafts in cancer cell adhesion and migration. Int. J. Cell Biol. 2012, 2012. [Google Scholar] [CrossRef]
- Rogers, K.R.; Kikawa, K.D.; Mouradian, M.; Hernandez, K.; McKinnon, K.M.; Ahwah, S.M.; Pardini, R.S. Docosahexaenoic acid alters epidermal growth factor receptor-related signaling by disrupting its lipid raft association. Carcinogenesis 2010, 31, 1523–1530. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.W.; Seo, J.; Davidson, L.A.; Callaway, E.S.; Fan, Y.Y.; Lupton, J.R.; Chapkin, R.S. n-3 PUFA alter caveolae lipid composition and resident protein localization in mouse colon. FASEB J. 2004, 18, 1040–1042. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Calder, P.C. Conversion of α-linolenic acid to longer-chain polyunsaturated fatty acids in human adults. Reprod. Nutr. Dev. 2005, 45, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Young, L.R.; Kurzer, M.S.; Thomas, W.; Redmon, J.B.; Raatz, S.K. Effect of dietary fat and omega-3 fatty acids on urinary eicosanoids and sex hormone concentrations in postmenopausal women: A randomized controlled feeding trial. Nutr. Cancer 2011, 63, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Healy, D.A.; Wallace, F.A.; Miles, E.A.; Calder, P.C.; Newsholm, P. Effect of low-to-moderate amounts of dietary fish oil on neutrophil lipid composition and function. Lipids 2000, 35, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Benoit, V.; Relic, B.; Leval, X.X.; Chariot, A.; Merville, M.P.; Bours, V. Regulation of HER-2 oncogene expression by cyclooxygenase-2 and prostaglandin E2. Oncogene 2004, 23, 1631–1635. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.A.; Brueggemeier, R.W. Prostaglandin E2 regulates aromatase activity and expression in human adipose stromal cells via two distinct receptor subtypes. J. Clin. Endocrinol. Metab. 2003, 88, 2810–2816. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Taylor, P.C.; Nelson, G.J.; Schmidt, P.C.; Ferretti, A.; Erickson, K.L.; Yu, R.; Chandra, R.K.; Mackey, B.E. Docosahexaenoic acid ingestion inhibits natural killer cell activity and production of inflammatory mediators in young healthy men. Lipids 1999, 34, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Wei, N.; Wang, B.; Zhang, Q.Y.; Mi, M.T.; Zhu, J.D.; Yu, X.P.; Yuan, J.L.; Chen, K.; Wang, J.; Chang, H. Effects of different dietary fatty acids on the fatty acid compositions and the expression of lipid metabolic-related genes in mammary tumor tissues of rats. Nutr. Cancer 2008, 60, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Novel eicosanoid and docosanoid mediators: Resolvins, docosatrienes, and neuroprotectins. Curr. Opin. Clin. Nutr. Metab. Care 2005, 8, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Arita, M.; Hong, S.; Gotlinger, K. Resolvins, docosatrienes, and neuroprotectins, novel omega-3-derived mediators, and their endogenous aspirin-triggered epimers. Lipids 2004, 39, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Weylandt, K.H.; Kang, J.X.; Wiedenmann, B.; Baumgart, D.C. Lipoxins and resolvins in inflammatory bowel disease. Inflamm. Bowel. Dis. 2007, 13, 797–799. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X.; Weylandt, K.H. Modulation of inflammatory cytokines by omega-3 fatty acids. Subcell. Biochem. 2008, 49, 133–143. [Google Scholar] [PubMed]
- Calder, P.C. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, 1505S–1519S. [Google Scholar] [PubMed]
- Zandi, R.; Larsen, A.B.; Andersen, P.; Stockhausen, M.T.; Poulsen, H.S. Mechanisms for oncogenic activation of the epidermal growth factor receptor. Cell Signal. 2007, 19, 2013–2023. [Google Scholar] [CrossRef] [PubMed]
- Corsetto, P.A.; Montorfano, G.; Zava, S.; Jovenitti, I.E.; Cremona, A.; Berra, B.; Rizzo, A.M. Effects of n-3 PUFAs on breast cancer cells through their incorporation in plasma membrane. Lipids Health Dis. 2011, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, R.; Gerald, W.L.; Li, A.R.; Pan, Q.; Lal, P.; Ladanyi, M.; Chen, B. EGFR gene amplification in breast cancer: Correlation with epidermal growth factor receptor mRNA and protein expression and HER-2 status and absence of EGFR-activating mutations. Mod. Pathol. 2005, 18, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Hynes, N.E.; Stern, D.F. The biology of erbB-2/neu/HER-2 and its role in cancer. Biochim. Biophys. Acta 1994, 1198, 165–184. [Google Scholar] [PubMed]
- Tovey, S.M.; Brown, S.; Doughty, J.C.; Mallon, E.A.; Cooke, T.G.; Edwards, J. Poor survival outcomes in HER2-positive breast cancer patients with low-grade, node-negative tumours. Br. J. Cancer 2009, 100, 680–683. [Google Scholar] [CrossRef] [PubMed]
- Moasser, M.M. The oncogene HER2: Its signaling and transforming functions and its role in human cancer pathogenesis. Oncogene 2007, 26, 6469–6487. [Google Scholar] [CrossRef] [PubMed]
- Bollig-Fischer, A.; Dziubinski, M.; Boyer, A.; Haddad, R.; Giroux, C.N.; Ethier, S.P. HER-2 signaling, acquisition of growth factor independence, and regulation of biological networks associated with cell transformation. Cancer Res. 2010, 70, 7862–7873. [Google Scholar] [CrossRef] [PubMed]
- Yee, L.D.; Agarwal, D.; Rosol, T.J.; Lehman, A.; Tian, M.; Hatton, J.; Heestand, J.; Belury, M.A.; Clinton, S.K. The inhibition of early stages of HER-2/neu-mediated mammary carcinogenesis by dietary n-3 PUFAs. Mol. Nutr. Food Res. 2013, 57, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Truan, J.S.; Chen, J.M.; Thompson, L.U. Flaxseed oil reduces the growth of human breast tumors (MCF-7) at high levels of circulating estrogen. Mol. Nutr. Food Res. 2010, 54, 1414–1421. [Google Scholar] [CrossRef] [PubMed]
- Desvergne, B.; Wahli, W. Peroxisome proliferator-activated receptors: Nuclear control of metabolism. Endocr. Rev. 1999, 20, 649–688. [Google Scholar] [PubMed]
- Georgiadi, A.; Kersten, S. Mechanisms of gene regulation by fatty acids. Adv. Nutr. 2012, 3, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Gurnell, M.; Savage, D.B.; Chatterjee, V.K.; O’Rahilly, S. The metabolic syndrome: Peroxisome proliferator-activated receptor gamma and its therapeutic modulation. J. Clin. Endocrinol. Metab. 2003, 88, 2412–2421. [Google Scholar] [CrossRef] [PubMed]
- Flachs, P.; Rossmeisl, M.; Bryhn, M.; Kopecky, J. Cellular and molecular effects of n-3 polyunsaturated fatty acids on adipose tissue biology and metabolism. Clin. Sci. 2009, 116, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kliewer, S.A.; Sundseth, S.S.; Jones, S.A.; Brown, P.J.; Wisely, G.B.; Koble, C.S.; Devchand, P.; Wahli, W.; Willson, T.M.; Lenhard, J.M.; et al. Fatty acids and eicosanoids regulate gene expression through direct interactions with peroxisome proliferator-activated receptors alpha and gamma. Proc. Natl. Acad. Sci. USA 1997, 94, 4318–4323. [Google Scholar] [CrossRef] [PubMed]
- Clay, C.E.; Namen, A.M.; Atsumi, G.; Willingham, M.C.; High, K.P.; Kute, T.E.; Trimboli, A.J.; Fonteh, A.N.; Dawson, P.A.; Chilton, F.H. Influence of J series prostaglandins on apoptosis and tumorigenesis of breast cancer cells. Carcinogenesis 1999, 20, 1905–1911. [Google Scholar] [CrossRef] [PubMed]
- Mueller, E.; Sarraf, P.; Tontonoz, P.; Evans, R.M.; Martin, K.J.; Zhang, M.; Fletcher, C.; Singer, S.; Spiegelman, B.M. Terminal differentiation of human breast cancer through PPAR gamma. Mol. Cell 1998, 1, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Barascu, A.; Besson, P.; Le, F.O.; Bougnoux, P.; Jourdan, M.L. CDK1-cyclin B1 mediates the inhibition of proliferation induced by omega-3 fatty acids in MDA-MB-231 breast cancer cells. Int. J. Biochem. Cell Biol. 2006, 38, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zhu, Z.; McGinley, J.N.; El, B.K.; Manni, A.; Thompson, H.J. Identification of a molecular signature underlying inhibition of mammary carcinoma growth by dietary n-3 fatty acids. Cancer Res. 2012, 72, 3795–3806. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.D.; Burne, J.F.; King, M.P.; Miyashita, T.; Reed, J.C.; Raff, M.C. Bcl-2 blocks apoptosis in cells lacking mitochondrial DNA. Nature 1993, 361, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Van Slooten, H.J.; Clahsen, P.C.; van Dierendonck, J.H.; Duval, C.; Pallud, C.; Mandard, A.M.; obelle-Deroide, A.; van de Velde, C.J.; van de Vijver, M.J. Expression of Bcl-2 in node-negative breast cancer is associated with various prognostic factors, but does not predict response to one course of perioperative chemotherapy. Br. J. Cancer 1996, 74, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Oltvai, Z.N.; Milliman, C.L.; Korsmeyer, S.J. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell 1993, 74, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Raisova, M.; Hossini, A.M.; Eberle, J.; Riebeling, C.; Wieder, T.; Sturm, I.; Daniel, P.T.; Orfanos, C.E.; Geilen, C.C. The Bax/Bcl-2 ratio determines the susceptibility of human melanoma cells to CD95/Fas-mediated apoptosis. J. Investig. Dermatol. 2001, 117, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Ion, G.; Akinsete, J.A.; Hardman, W.E. Maternal consumption of canola oil suppressed mammary gland tumorigenesis in C3(1) TAg mice offspring. BMC Cancer 2010, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Vivanco, I.; Sawyers, C.L. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat. Rev. Cancer 2002, 2, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Ueno, L.; Vogt, P.K. Akt-mediated regulation of NFκB and the essentialness of NFκB for the oncogenicity of PI3K and Akt. Int. J. Cancer 2009, 125, 2863–2870. [Google Scholar] [CrossRef] [PubMed]
- Schley, P.D.; Jijon, H.B.; Robinson, L.E.; Field, C.J. Mechanisms of omega-3 fatty acid-induced growth inhibition in MDA-MB-231 human breast cancer cells. Breast Cancer Res. Treat. 2005, 92, 187–195. [Google Scholar] [CrossRef] [PubMed]
- DeGraffenried, L.A.; Friedrichs, W.E.; Fulcher, L.; Fernandes, G.; Silva, J.M.; Peralba, J.M.; Hidalgo, M. Eicosapentaenoic acid restores tamoxifen sensitivity in breast cancer cells with high Akt activity. Ann. Oncol. 2003, 14, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, K.M.; Anderson, N.G. The protein kinase B/Akt signalling pathway in human malignancy. Cell Signal. 2002, 14, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.R.; Ahmed, S.O.; Ahmed, M.; Khan, O.S.; Al, A.S.; Platanias, L.C.; Al-Kuraya, K.S.; Uddin, S. Cross-talk between NFκB and the PI3-kinase/AKT pathway can be targeted in primary effusion lymphoma (PEL) cell lines for efficient apoptosis. PLoS One 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Dolcet, X.; Llobet, D.; Pallares, J.; Matias-Guiu, X. NF-κB in development and progression of human cancer. Virchows Arch. 2005, 446, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.T.; Kral, J.G. The NF-κB/IκB signaling system: A molecular target in breast cancer therapy. J. Surg. Res. 2005, 123, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Mayo, M.W.; Wang, C.Y.; Cogswell, P.C.; Rogers-Graham, K.S.; Lowe, S.W.; Der, C.J.; Baldwin, A.S., Jr. Requirement of NF-κB activation to suppress p53-independent apoptosis induced by oncogenic Ras. Science 1997, 278, 1812–1815. [Google Scholar] [CrossRef] [PubMed]
- Yerushalmi, R.; Woods, R.; Ravdin, P.M.; Hayes, M.M.; Gelmon, K.A. Ki67 in breast cancer: Prognostic and predictive potential. Lancet Oncol. 2010, 11, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Scholzen, T.; Gerdes, J. The Ki-67 protein: From the known and the unknown. J. Cell. Physiol. 2000, 182, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Saggar, J.K.; Chen, J.; Corey, P.; Thompson, L.U. Dietary flaxseed lignan or oil combined with tamoxifen treatment affects MCF-7 tumor growth through estrogen receptor- and growth factor-signaling pathways. Mol. Nutr. Food Res. 2010, 54, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Taftachi, R.; Ayhan, A.; Ekici, S.; Ergen, A.; Ozen, H. Proliferating-cell nuclear antigen (PCNA) as an independent prognostic marker in patients after prostatectomy: A comparison of PCNA and Ki-67. BJU Int. 2005, 95, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Paunesku, T.; Mittal, S.; Protić, M.; Oryhon, J.; Korolev, S.V.; Joachimiak, A.; Woloschak, G.E. Proliferating cell nuclear antigen (PCNA): Ringmaster of the genome. Int. J. Radiat. Biol. 2001, 77, 1007–1021. [Google Scholar] [CrossRef] [PubMed]
- Malkas, L.H.; Herbert, B.S.; bdel-Aziz, W.; Dobrolecki, L.E.; Liu, Y.; Agarwal, B.; Hoelz, D.; Badve, S.; Schnaper, L.; Arnold, R.J.; et al. A cancer-associated PCNA expressed in breast cancer has implications as a potential biomarker. Proc. Natl. Acad. Sci. USA 2006, 103, 19472–19477. [Google Scholar] [CrossRef] [PubMed]
- Strzalka, W.; Ziemienowicz, A. Proliferating cell nuclear antigen (PCNA): A key factor in DNA replication and cell cycle regulation. Ann. Bot. 2011, 107, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- Olivo, S.E.; Hilakivi-Clarke, L. Opposing effects of prepubertal low- and high-fat n-3 polyunsaturated fatty acid diets on rat mammary tumorigenesis. Carcinogenesis 2005, 26, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Baek, S.H. Mouse models for breast cancer metastasis. Biochem. Biophys. Res. Commun. 2010, 394, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Karmali, R.A.; Marsh, J.; Fuchs, C. Effect of omega-3 fatty acids on growth of a rat mammary tumor. J. Natl. Cancer Inst. 1984, 73, 457–461. [Google Scholar] [PubMed]
- Gonzalez, M.J.; Schemmel, R.A.; Gray, J.I.; Dugan, L., Jr.; Sheffield, L.G.; Welsch, C.W. Effect of dietary fat on growth of MCF-7 and MDA-MB231 human breast carcinomas in athymic nude mice: Relationship between carcinoma growth and lipid peroxidation product levels. Carcinogenesis 1991, 12, 1231–1235. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, N.E.; Lim, D.; Erickson, K.L. Alteration of murine mammary tumorigenesis by dietary enrichment with n-3 fatty acids in fish oil. Cancer Lett. 1998, 124, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yee, L.D.; Young, D.C.; Rosol, T.J.; Vanbuskirk, A.M.; Clinton, S.K. Dietary (n-3) polyunsaturated fatty acids inhibit HER-2/neu-induced breast cancer in mice independently of the PPARgamma ligand rosiglitazone. J. Nutr. 2005, 135, 983–988. [Google Scholar] [PubMed]
- MacLennan, M.B.; Clarke, S.E.; Perez, K.; Wood, G.A.; Muller, W.J.; Kang, J.X.; Ma, D.W. Mammary tumor development is directly inhibited by lifelong n-3 polyunsaturated fatty acids. J. Nutr. Biochem. 2013, 24, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Dankort, D.; Maslikowski, B.; Warner, N.; Kanno, N.; Kim, H.; Wang, Z.; Moran, M.F.; Oshima, R.G.; Cardiff, R.D.; Muller, W.J. Grb2 and Shc adapter proteins play distinct roles in Neu (ErbB-2)-induced mammary tumorigenesis: Implications for human breast cancer. Mol. Cell Biol. 2001, 21, 1540–1551. [Google Scholar] [CrossRef] [PubMed]
- Yee, L.D.; Lester, J.L.; Cole, R.M.; Richardson, J.R.; Hsu, J.C.; Li, Y.; Lehman, A.; Belury, M.A.; Clinton, S.K. Omega-3 fatty acid supplements in women at high risk of breast cancer have dose-dependent effects on breast adipose tissue fatty acid composition. Am. J. Clin. Nutr. 2010, 91, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Luijten, M.; Verhoef, A.; Dormans, J.A.; Beems, R.B.; Cremers, H.W.; Nagelkerke, N.J.; Adlercreutz, H.; Penalvo, J.L.; Piersma, A.H. Modulation of mammary tumor development in Tg.NK (MMTV/c-neu) mice by dietary fatty acids and life stage-specific exposure to phytoestrogens. Reprod. Toxicol. 2007, 23, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Yuri, T.; Danbara, N.; Tsujita-Kyutoku, M.; Fukunaga, K.; Takada, H.; Inoue, Y.; Hada, T.; Tsubura, A. Dietary docosahexaenoic acid suppresses N-methyl-N-nitrosourea-induced mammary carcinogenesis in rats more effectively than eicosapentaenoic acid. Nutr. Cancer 2003, 45, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Rose, D.P.; Connolly, J.M. Effects of fatty acids and inhibitors of eicosanoid synthesis on the growth of a human breast cancer cell line in culture. Cancer Res. 1990, 50, 7139–7144. [Google Scholar] [PubMed]
- Grammatikos, S.I.; Subbaiah, P.V.; Victor, T.A.; Miller, W.M. n-3 and n-6 fatty acid processing and growth effects in neoplastic and non-cancerous human mammary epithelial cell lines. Br. J. Cancer 1994, 70, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Kachhap, S.K.; Dange, P.P.; Santani, R.H.; Sawant, S.S.; Ghosh, S.N. Effect of omega-3 fatty acid (docosahexanoic acid) on BRCA1 gene expression and growth in MCF-7 cell line. Cancer Biother. Radiopharm. 2001, 16, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Chajes, V.; Sattler, W.; Stranzl, A.; Kostner, G.M. Influence of n-3 fatty acids on the growth of human breast cancer cells in vitro: Relationship to peroxides and vitamin-E. Breast Cancer Res. Treat. 1995, 34, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Chamras, H.; Ardashian, A.; Heber, D.; Glaspy, J.A. Fatty acid modulation of MCF-7 human breast cancer cell proliferation, apoptosis and differentiation. J. Nutr. Biochem. 2002, 13, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Rose, D.P.; Connolly, J.M.; Coleman, M. Effect of ω-3 fatty acids on the progression of metastases after the surgical excision of human breast cancer cell solid tumors growing in nude mice. Clin. Cancer Res. 1996, 2, 1751–1756. [Google Scholar] [PubMed]
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, interconversion, and dose response of n-3 fatty acids in humans. Am. J. Clin. Nutr. 2006, 83, 1467S–1476S. [Google Scholar] [PubMed]
- Harper, C.R.; Edwards, M.J.; DeFilippis, A.P.; Jacobson, T.A. Flaxseed oil increases the plasma concentrations of cardioprotective (n-3) fatty acids in humans. J. Nutr. 2006, 136, 83–87. [Google Scholar] [PubMed]
- Pawlosky, R.J.; Hibbeln, J.R.; Novotny, J.A.; Salem, N., Jr. Physiological compartmental analysis of α-linolenic acid metabolism in adult humans. J. Lipid Res. 2001, 42, 1257–1265. [Google Scholar] [PubMed]
- Brenna, J.T. Efficiency of conversion of α-linolenic acid to long chain n-3 fatty acids in man. Curr. Opin. Clin. Nutr. Metab Care 2002, 5, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, O.; Takahashi, M.; Shigematsu, T.; Shimamura, K.; Kimura, J.; Ezaki, H.; Gotoh, T. Long-term effects of dietary α-linolenic acid from perilla oil on serum fatty acids composition and on the risk factors of coronary heart disease in Japanese elderly subjects. J. Nutr. Sci. Vitaminol. 1999, 45, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Ghafoorunissa; Vani, A.; Laxmi, R.; Sesikeran, B. Effects of dietary α-linolenic acid from blended oils on biochemical indices of coronary heart disease in Indians. Lipids 2002, 37, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Klein, V.; Chajes, V.; Germain, E.; Schulgen, G.; Pinault, M.; Malvy, D.; Lefrancq, T.; Fignon, A.; Le, F.O.; Lhuillery, C.; Bougnoux, P. Low α-linolenic acid content of adipose breast tissue is associated with an increased risk of breast cancer. Eur. J. Cancer 2000, 36, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Bougnoux, P.; Koscielny, S.; Chajes, V.; Descamps, P.; Couet, C.; Calais, G. α-Linolenic acid content of adipose breast tissue: A host determinant of the risk of early metastasis in breast cancer. Br. J. Cancer 1994, 70, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, K.L.; Johnston, P.V. Effect of dietary α-linolenic acid on growth, metastasis, fatty acid profile and prostaglandin production of two murine mammary adenocarcinomas. J. Nutr. 1990, 120, 1601–1609. [Google Scholar] [PubMed]
- Hardman, W.E. Dietary canola oil suppressed growth of implanted MDA-MB 231 human breast tumors in nude mice. Nutr. Cancer 2007, 57, 177–183. [Google Scholar] [CrossRef]
- Mabasa, L.; Cho, K.; Walters, M.W.; Bae, S.; Park, C.S. Maternal dietary canola oil suppresses growth of mammary carcinogenesis in female rat offspring. Nutr. Cancer 2013, 65, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Menendez, J.A.; Vazquez-Martin, A.; Ropero, S.; Colomer, R.; Lupu, R. HER2 (erbB-2)-targeted effects of the omega-3 polyunsaturated fatty acid, α-linolenic acid (ALA; 18:3n-3), in breast cancer cells: The “fat features” of the “Mediterranean diet” as an “anti-HER2 cocktail”. Clin. Transl. Oncol. 2006, 8, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Menendez, J.A.; Ropero, S.; Lupu, R.; Colomer, R. Dietary fatty acids regulate the activation status of Her-2/neu (c-erbB-2) oncogene in breast cancer cells. Ann. Oncol. 2004, 15, 1719–1721. [Google Scholar] [CrossRef] [PubMed]
- Mason, J.K.; Fu, M.H.; Chen, J.; Yu, Z.; Thompson, L.U. Dietary flaxseed-trastuzumab interactive effects on the growth of HER2-overexpressing human breast tumors (BT-474). Nutr. Cancer 2013, 65, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Hirose, M.; Masuda, A.; Ito, N.; Kamano, K.; Okuyama, H. Effects of dietary perilla oil, soybean oil and safflower oil on 7,12-dimethylbenz (a) anthracene (DMBA) and 1,2-dimethyl-hydrazine (DMH)-induced mammary gland and colon carcinogenesis in female SD rats. Carcinogenesis 1990, 11, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.C.; Wan, J.M. Induction of apoptosis in HL-60 cells by eicosapentaenoic acid (EPA) is associated with downregulation of bcl-2 expression. Cancer Lett. 1999, 145, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Senzaki, H.; Iwamoto, S.; Ogura, E.; Kiyozuka, Y.; Arita, S.; Kurebayashi, J.; Takada, H.; Hioki, K.; Tsubura, A. Dietary effects of fatty acids on growth and metastasis of KPL-1 human breast cancer cells in vivo and in vitro. Anticancer Res. 1998, 18, 1621–1627. [Google Scholar] [PubMed]
- Sauer, L.A.; Dauchy, R.T.; Blask, D.E.; Krause, J.A.; Davidson, L.K.; Dauchy, E.M. Eicosapentaenoic acid suppresses cell proliferation in MCF-7 human breast cancer xenografts in nude rats via a pertussis toxin-sensitive signal transduction pathway. J. Nutr. 2005, 135, 2124–2129. [Google Scholar] [PubMed]
- Chambrier, C.; Bastard, J.P.; Rieusset, J.; Chevillotte, E.; Bonnefont-Rousselot, D.; Therond, P.; Hainque, B.; Riou, J.P.; Laville, M.; Vidal, H. Eicosapentaenoic acid induces mRNA expression of peroxisome proliferator-activated receptor gamma. Obes. Res. 2002, 10, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Wang, P.; Yamabe, N.; Fukui, M.; Jay, T.; Zhu, B.T. Docosahexaenoic acid induces apoptosis in MCF-7 cells in vitro and in vivo via reactive oxygen species formation and caspase 8 activation. PLoS One 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Blanckaert, V.; Ulmann, L.; Mimouni, V.; Antol, J.; Brancquart, L.; Chenais, B. Docosahexaenoic acid intake decreases proliferation, increases apoptosis and decreases the invasive potential of the human breast carcinoma cell line MDA-MB-231. Int. J. Oncol. 2010, 36, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.C.; Wong, E.Y.; Ooi, V.E. Docosahexaenoic acid from a cultured microalga inhibits cell growth and induces apoptosis by upregulating Bax/Bcl-2 ratio in human breast carcinoma MCF-7 cells. Ann. N. Y. Acad. Sci. 2004, 1030, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Tsujita-Kyutoku, M.; Yuri, T.; Danbara, N.; Senzaki, H.; Kiyozuka, Y.; Uehara, N.; Takada, H.; Hada, T.; Miyazawa, T.; Ogawa, Y.; et al. Conjugated docosahexaenoic acid suppresses KPL-1 human breast cancer cell growth in vitro and in vivo: Potential mechanisms of action. Breast Cancer Res. 2004, 6, R291–R299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connolly, J.M.; Gilhooly, E.M.; Rose, D.P. Effects of reduced dietary linoleic acid intake, alone or combined with an algal source of docosahexaenoic acid, on MDA-MB-231 breast cancer cell growth and apoptosis in nude mice. Nutr. Cancer 1999, 35, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Jourdan, M.L.; Maheo, K.; Barascu, A.; Goupille, C.; de Latour, M.P.; Bougnoux, P.; Rio, P.G. Increased BRCA1 protein in mammary tumours of rats fed marine omega-3 fatty acids. Oncol. Rep. 2007, 17, 713–719. [Google Scholar] [PubMed]
- Bernard-Gallon, D.J.; Vissac-Sabatier, C.; ntoine-Vincent, D.; Rio, P.G.; Maurizis, J.C.; Fustier, P.; Bignon, Y.J. Differential effects of n-3 and n-6 polyunsaturated fatty acids on BRCA1 and BRCA2 gene expression in breast cell lines. Br. J. Nutr. 2002, 87, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Bougnoux, P.; Chajes, V.; Germain, E.; Hubert, B.; Lhuillery, C.; Le, F.O.; Body, G.; Calais, G. Cytotoxic drug efficacy correlates with adipose tissue docosahexaenoic acid level in locally advanced breast carcinoma. Lipids 1999, 34, S109. [Google Scholar] [CrossRef] [PubMed]
- Bougnoux, P.; Hajjaji, N.; Ferrasson, M.N.; Giraudeau, B.; Couet, C.; Le, F.O. Improving outcome of chemotherapy of metastatic breast cancer by docosahexaenoic acid: A phase II trial. Br. J. Cancer 2009, 101, 1978–1985. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Veigas, J.M.; Williams, P.J.; Fernandes, G. DHA is a more potent inhibitor of breast cancer metastasis to bone and related osteolysis than EPA. Breast Cancer Res. Treat. 2013, 141, 341–352. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Ma, D.W.L. The Role of n-3 Polyunsaturated Fatty Acids in the Prevention and Treatment of Breast Cancer. Nutrients 2014, 6, 5184-5223. https://doi.org/10.3390/nu6115184
Liu J, Ma DWL. The Role of n-3 Polyunsaturated Fatty Acids in the Prevention and Treatment of Breast Cancer. Nutrients. 2014; 6(11):5184-5223. https://doi.org/10.3390/nu6115184
Chicago/Turabian StyleLiu, Jiajie, and David W. L. Ma. 2014. "The Role of n-3 Polyunsaturated Fatty Acids in the Prevention and Treatment of Breast Cancer" Nutrients 6, no. 11: 5184-5223. https://doi.org/10.3390/nu6115184