Chemopreventive Effects of Oplopantriol A, a Novel Compound Isolated from Oplopanax horridus, on Colorectal Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Materials and Methods
2.1. Chemicals and Reagents
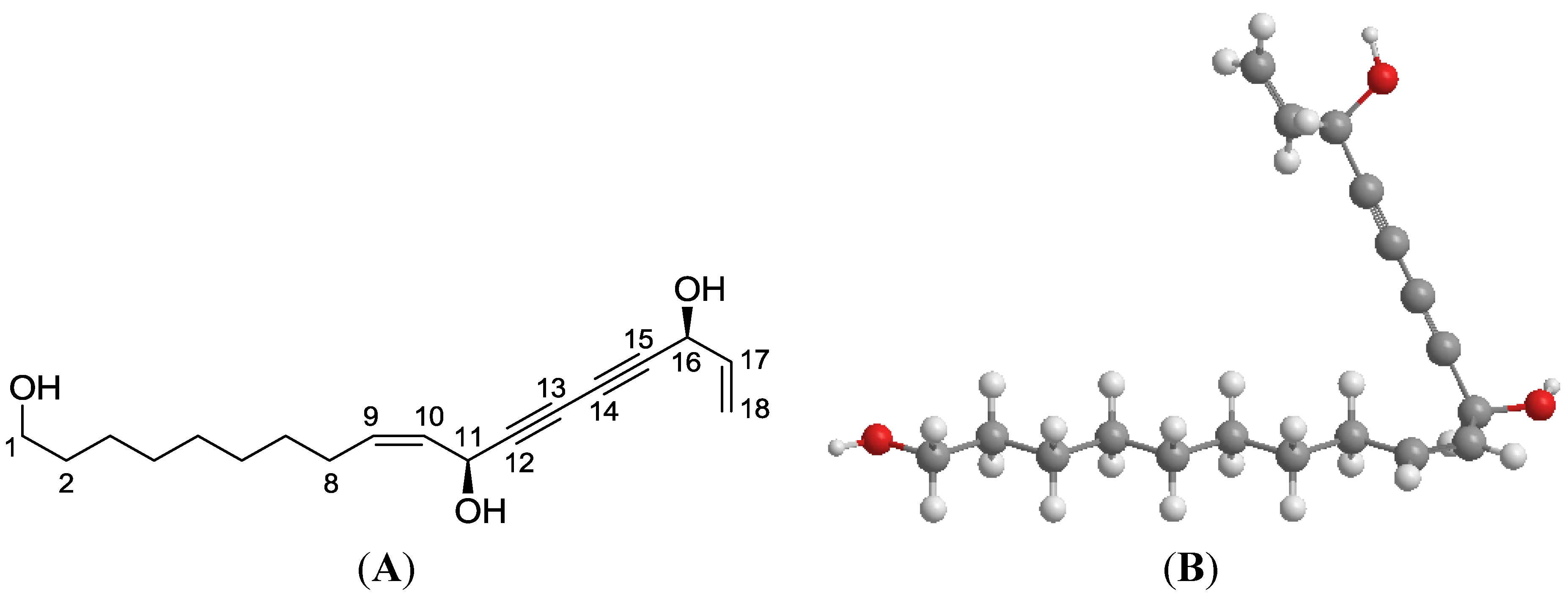
2.2. Plant Material, Isolation and Structural Elucidation
2.3. Human Cancer Cell Lines and Cell Culture
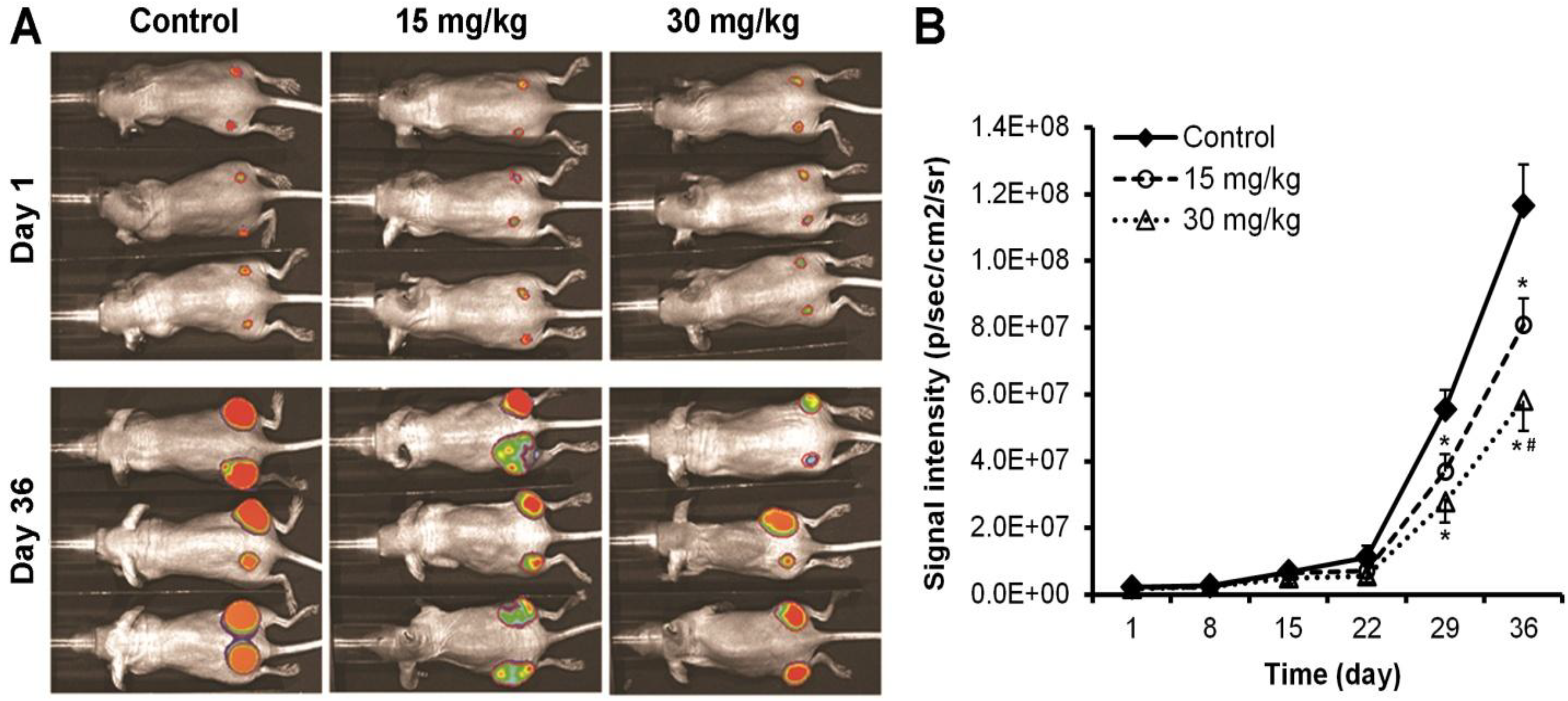
2.4. Xenograft Tumor Xenogen Bioluminescence Imaging
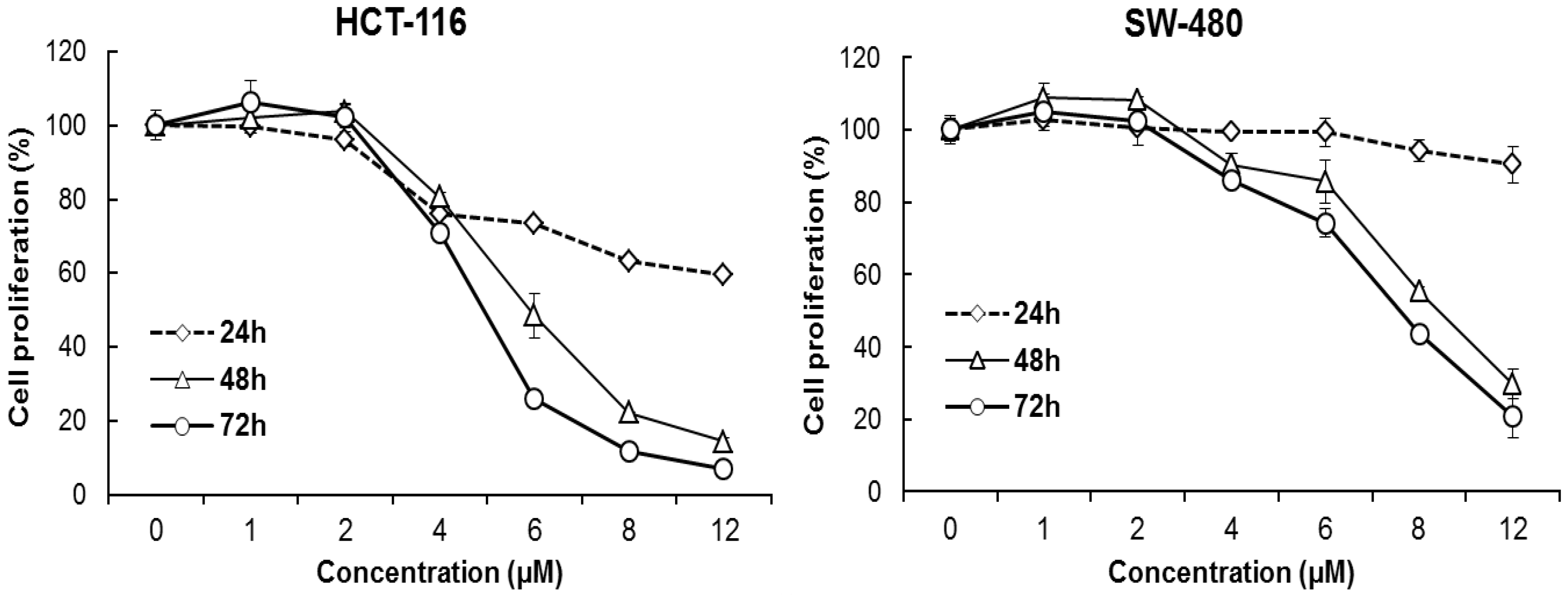
2.5. MTS Assays
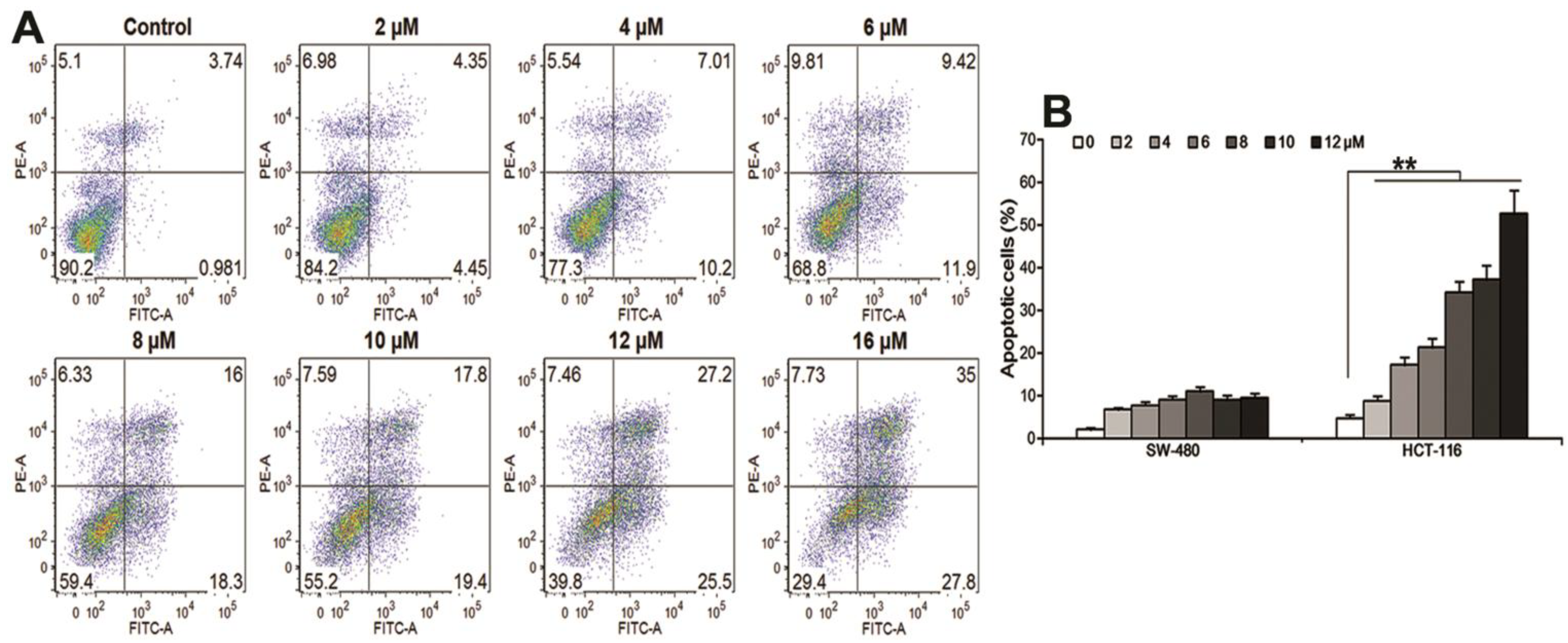
2.6. Apoptosis Assay
2.7. Cell Cycle Assay
2.8. Real-Time PCR Array of Apoptosis Screening Analysis
2.9. Statistical Analysis
3. Results
3.1. Oplopantriol A Inhibited Colon Tumor Growth

3.2. Oplopantriol A Inhibited HCT-116 and SW-480 Cell Proliferation
3.3. Oplopantriol A Induced Apoptosis


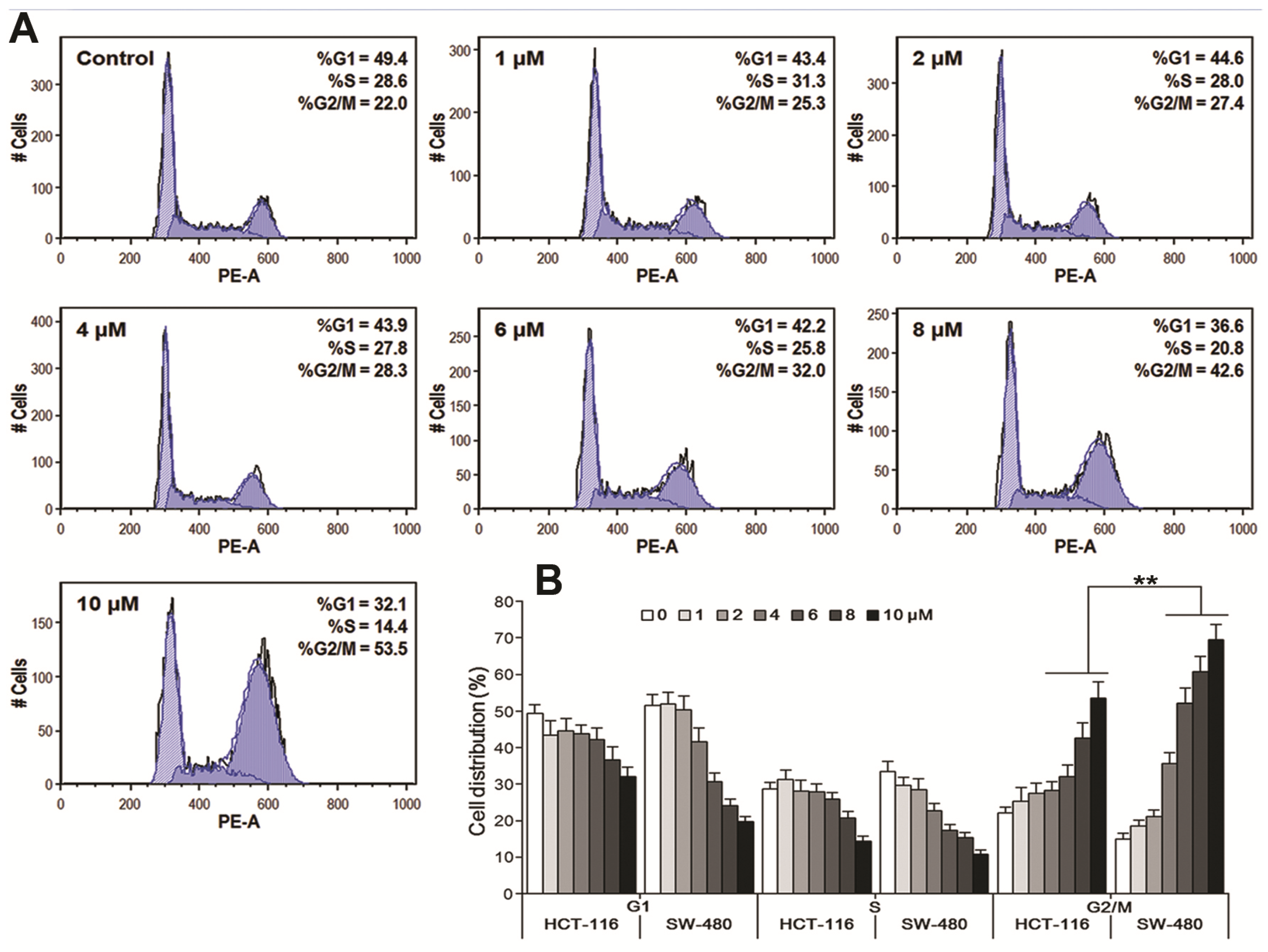
3.4. Oplopantriol A Arrested Cell Cycle at the G2/M Phase

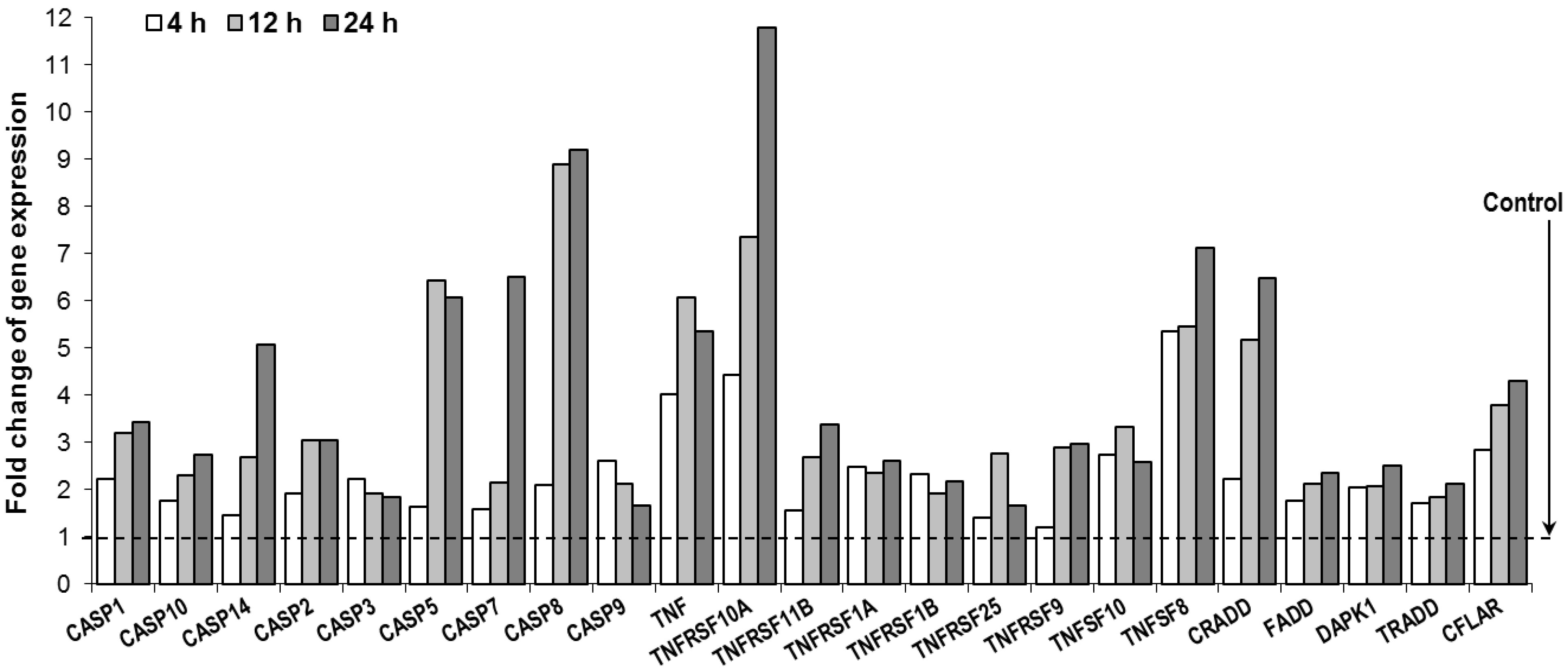
3.5. Oplopantriol A Induced Apoptosis Related Gene Expression and TNF-Cell Death Pathways

4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Calway, T.; Du, G.J.; Wang, C.Z.; Huang, W.H.; Zhao, J.; Li, S.P.; Yuan, C.S. Chemical and pharmacological studies of Oplopanax horridus, a North American botanical. J. Nat. Med. 2012, 66, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Lantz, T.C.; Swerhun, K.; Turner, N.J. Devil’s club (Oplopanax horridus): An ethnobotanical review. HerbalGram 2004, 62, 33–48. [Google Scholar]
- Lantz, T.C.; Antos, J.A. Clonal expansion in the deciduous understory shrub, devil’s club (Oplopanax horridus; Araliaceae). Can. J. Bot. 2002, 80, 1052–1062. [Google Scholar] [CrossRef]
- Huang, W.H.; Zhang, Q.W.; Yuan, C.S.; Wang, C.Z.; Li, S.P.; Zhou, H.H. Chemical constituents of the plants from the genus oplopanax. Chem. Biodivers. 2014, 11, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Zhang, Z.; Huang, W.H.; Du, G.J.; Wen, X.D.; Calway, T.; Yu, C.; Nass, R.; Zhao, J.; Du, W. Identification of potential anticancer compounds from Oplopanax horridus. Phytomedicine 2013, 20, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Inui, T.; Wang, Y.; Nikolic, D.; Smith, D.C.; Franzblau, S.G.; Pauli, G.F. Sesquiterpenes from Oplopanax horridus. J. Nat. Prod. 2010, 73, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.H.; Zhang, Q.W.; Wang, C.Z.; Yuan, C.S.; Li, S.P. Isolation and identification of two new polyynes from a North American ethnic medicinal plant—Oplopanax horridus (Smith) Miq. Molecules 2010, 15, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Rajpal, S.; Venook, A.P. Targeted therapy in colorectal cancer. Clin. Adv. Hematol. Oncol. 2006, 4, 124–132. [Google Scholar] [PubMed]
- Siegel, R.; Desantis, C.; Jemal, A. Colorectal cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Abdelfadil, E.; Cheng, Y.H.; Bau, D.T.; Ting, W.J.; Chen, L.M.; Hsu, H.H.; Lin, Y.M.; Chen, R.J.; Tsai, F.J.; Tsai, C.H.; et al. Thymoquinone induces apoptosis in oral cancer cells through p38β inhibition. Am. J. Chin. Med. 2013, 41, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Calway, T.; Yuan, C.S. Herbal medicines as adjuvants for cancer therapeutics. Am. J. Chin. Med. 2012, 40, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Du, G.J.; Zhang, Z.; Wen, X.D.; Yu, C.; Calway, T.; Yuan, C.S.; Wang, C.Z. Epigallocatechin gallate (EGCG) is the most effective cancer chemopreventive polyphenol in green tea. Nutrients 2012, 4, 1679–1691. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of natural products on developing new anti-cancer agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Wang, C.Z.; Chen, J.; Song, W.X.; Luo, J.; Tang, N.; He, B.C.; Kang, Q.; Wang, Y.; Du, W.; et al. Characterization of gene expression regulated by American ginseng and ginsenoside Rg3 in human colorectal cancer cells. Int. J. Oncol. 2008, 32, 975–983. [Google Scholar] [PubMed]
- He, B.C.; Gao, J.L.; Luo, X.; Luo, J.; Shen, J.; Wang, L.; Zhou, Q.; Wang, Y.T.; Luu, H.H.; Haydon, R.C.; et al. Ginsenoside Rg3 inhibits colorectal tumor growth through the down-regulation of Wnt/ss-catenin signaling. Int. J. Oncol. 2011, 38, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Wang, C.Z.; Mehendale, S.R.; Sun, S.; Wang, Q.; Yuan, C.S. Panaxadiol, a purified ginseng component, enhances the anti-cancer effects of 5-fluorouracil in human colorectal cancer cells. Cancer Chemother. Pharmacol. 2009, 64, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wu, G.; Wei, X.; Chen, X.; Wang, Y.; Chen, L. Celastrol induced DNA damage, cell cycle arrest, and apoptosis in human rheumatoid fibroblast-like synovial cells. Am. J. Chin. Med. 2013, 41, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Li, X.L.; Wang, Q.F.; Mehendale, S.R.; Yuan, C.S. Selective fraction of Scutellaria baicalensis and its chemopreventive effects on MCF-7 human breast cancer cells. Phytomedicine 2010, 17, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Calway, T.D.; Wen, X.D.; Smith, J.; Yu, C.; Wang, Y.; Mehendale, S.R.; Yuan, C.S. Hydrophobic flavonoids from Scutellaria baicalensis induce colorectal cancer cell apoptosis through a mitochondrial-mediated pathway. Int. J. Oncol. 2013, 42, 1018–1026. [Google Scholar] [PubMed]
- Chaiyakunapruk, N.; Kitikannakorn, N.; Nathisuwan, S.; Leeprakobboon, K.; Leelasettagool, C. The efficacy of ginger for the prevention of postoperative nausea and vomiting: A meta-analysis. Am. J. Obstet. Gynecol. 2006, 194, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Zhang, H.Y. Western-medicine-validated anti-tumor agents and traditional Chinese medicine. Trends Mol. Med. 2008, 14, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Morton, C.L.; Houghton, P.J. Establishment of human tumor xenografts in immunodeficient mice. Nat. Protoc. 2007, 2, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kohno, H.; Suzuki, R.; Yamada, Y.; Sugie, S.; Mori, H. A novel inflammation-related mouse colon carcinogenesis model induced by azoxymethane and dextran sodium sulfate. Cancer Sci. 2003, 94, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Swamy, M.V.; Patlolla, J.M.; Steele, V.E.; Kopelovich, L.; Reddy, B.S.; Rao, C.V. Chemoprevention of familial adenomatous polyposis by low doses of atorvastatin and celecoxib given individually and in combination to APCMin mice. Cancer Res. 2006, 66, 7370–7377. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Mori, H. Multistep carcinogenesis of the colon in Apc(Min/+) mouse. Cancer Sci. 2007, 98, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Neufert, C.; Becker, C.; Neurath, M.F. An inducible mouse model of colon carcinogenesis for the analysis of sporadic and inflammation-driven tumor progression. Nat. Protoc. 2007, 2, 1998–2004. [Google Scholar] [CrossRef] [PubMed]
- Din, F.V.; Dunlop, M.G.; Stark, L.A. Evidence for colorectal cancer cell specificity of aspirin effects on NF kappa B signalling and apoptosis. Br. J. Cancer 2004, 91, 381–388. [Google Scholar] [PubMed]
- Buchynska, L.G.; Nesina, I.P.; Yurchenko, N.P.; Bilyk, O.O.; Grinkevych, V.N.; Svintitsky, V.S. Expression of p53, p21WAF1/CIP1, p16INK4A and Ki-67 proteins in serous ovarian tumors. Exp. Oncol. 2007, 29, 49–53. [Google Scholar] [PubMed]
- Cimoli, G.; Malacarne, D.; Ponassi, R.; Valenti, M.; Alberti, S.; Parodi, S. Meta-analysis of the role of p53 status in isogenic systems tested for sensitivity to cytotoxic antineoplastic drugs. Biochim. Biophys. Acta 2004, 1705, 103–120. [Google Scholar] [PubMed]
- Hartmann, A.; Troadec, J.D.; Hunot, S.; Kikly, K.; Faucheux, B.A.; Mouatt-Prigent, A.; Ruberg, M.; Agid, Y.; Hirsch, E.C. Caspase-8 is an effector in apoptotic death of dopaminergic neurons in Parkinson’s disease, but pathway inhibition results in neuronal necrosis. J. Neurosci. 2001, 21, 2247–2255. [Google Scholar] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, Z.; Yu, C.; Zhang, C.-F.; Wu, X.-H.; Wen, X.-D.; Anderson, S.; Du, W.; Huang, W.-H.; Li, S.-P.; Wang, C.-Z.; et al. Chemopreventive Effects of Oplopantriol A, a Novel Compound Isolated from Oplopanax horridus, on Colorectal Cancer. Nutrients 2014, 6, 2668-2680. https://doi.org/10.3390/nu6072668
Zhang Z, Yu C, Zhang C-F, Wu X-H, Wen X-D, Anderson S, Du W, Huang W-H, Li S-P, Wang C-Z, et al. Chemopreventive Effects of Oplopantriol A, a Novel Compound Isolated from Oplopanax horridus, on Colorectal Cancer. Nutrients. 2014; 6(7):2668-2680. https://doi.org/10.3390/nu6072668
Chicago/Turabian StyleZhang, Zhiyu, Chunhao Yu, Chun-Feng Zhang, Xiao-Hui Wu, Xiao-Dong Wen, Samantha Anderson, Wei Du, Wei-Hua Huang, Shao-Ping Li, Chong-Zhi Wang, and et al. 2014. "Chemopreventive Effects of Oplopantriol A, a Novel Compound Isolated from Oplopanax horridus, on Colorectal Cancer" Nutrients 6, no. 7: 2668-2680. https://doi.org/10.3390/nu6072668