Chicken Essence Improves Exercise Performance and Ameliorates Physical Fatigue


Abstract
:1. Introduction
2. Results
2.1. Effect of CE Supplementation on Forelimb Grip Strength

2.2. Effect of CE Supplementation on Exercise Performance
2.3. Effect of CE Supplementation on Fatigue-Related Biochemical Variables after Acute Exercise Challenge


2.4. Effect of CE Supplementation on Tissue Glycogen Level

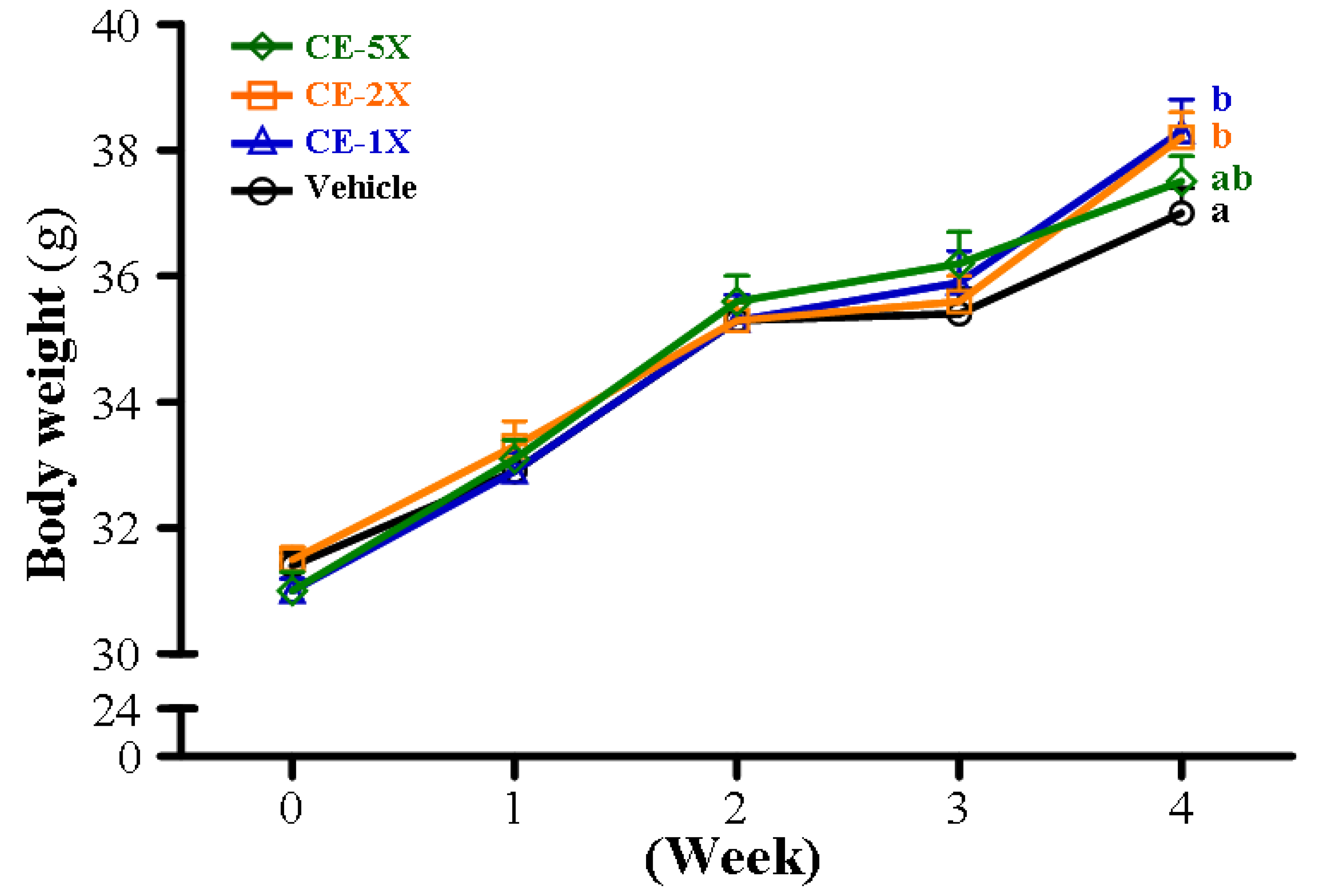
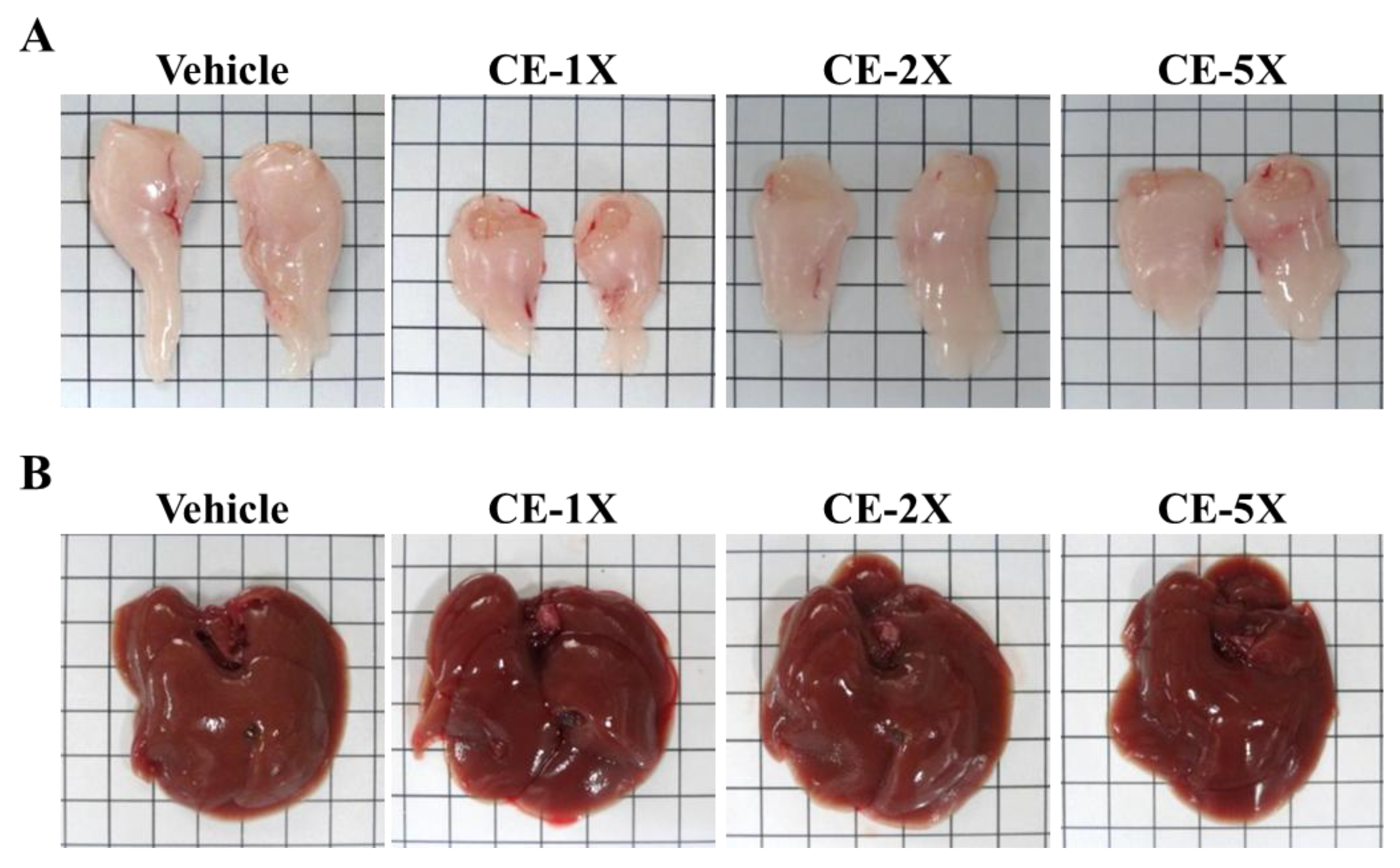
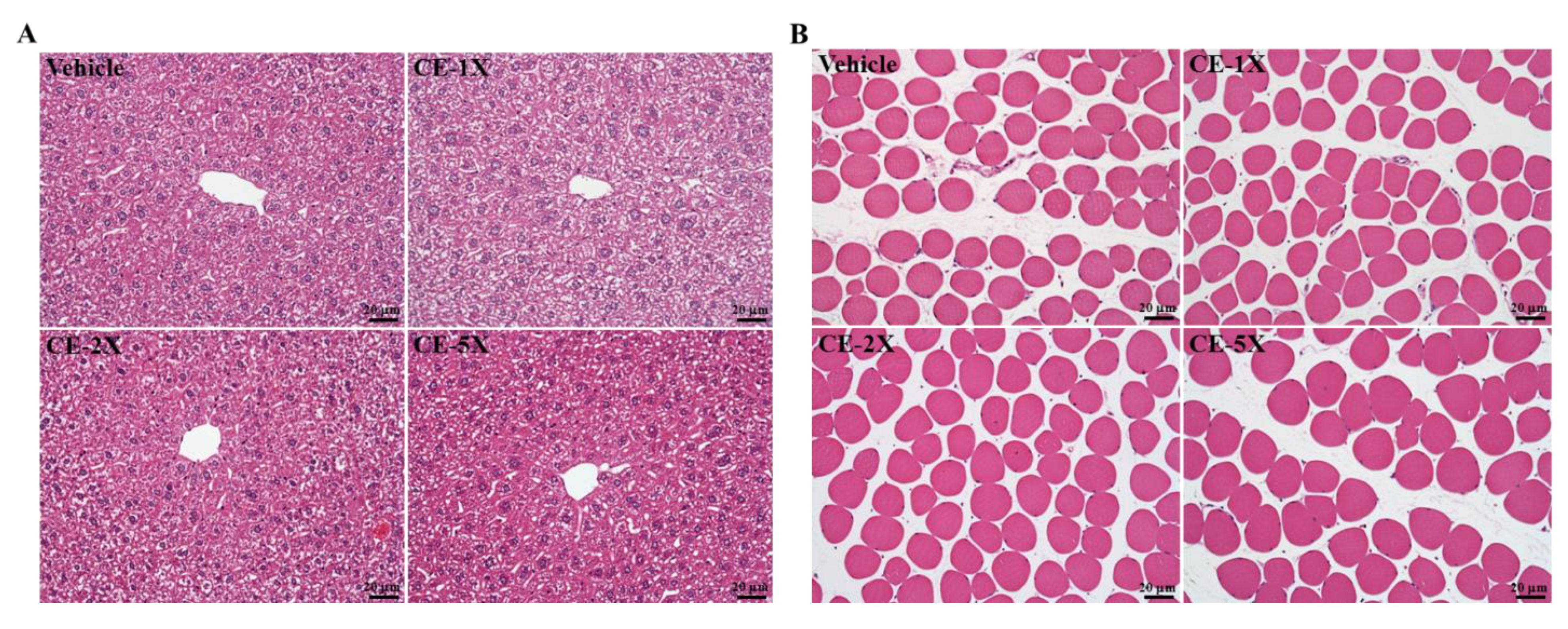
2.5. Subchronic Toxicity Evaluation of CE Supplementation


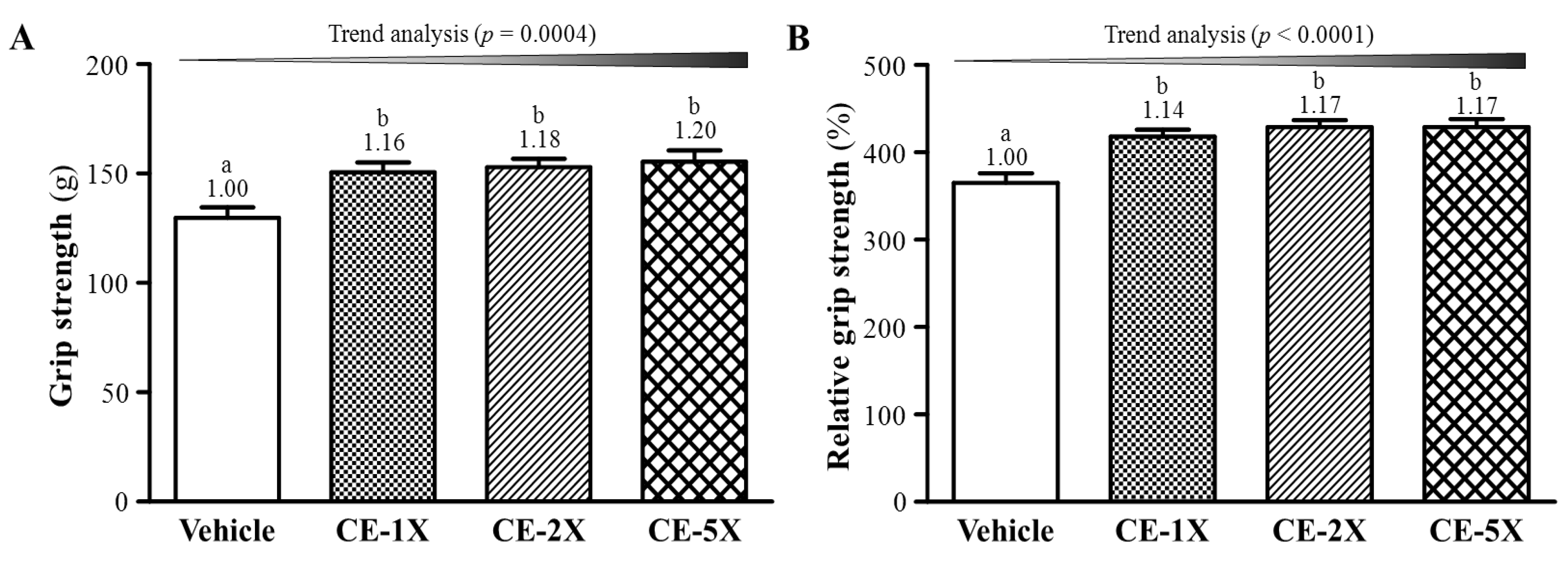{kind=link}
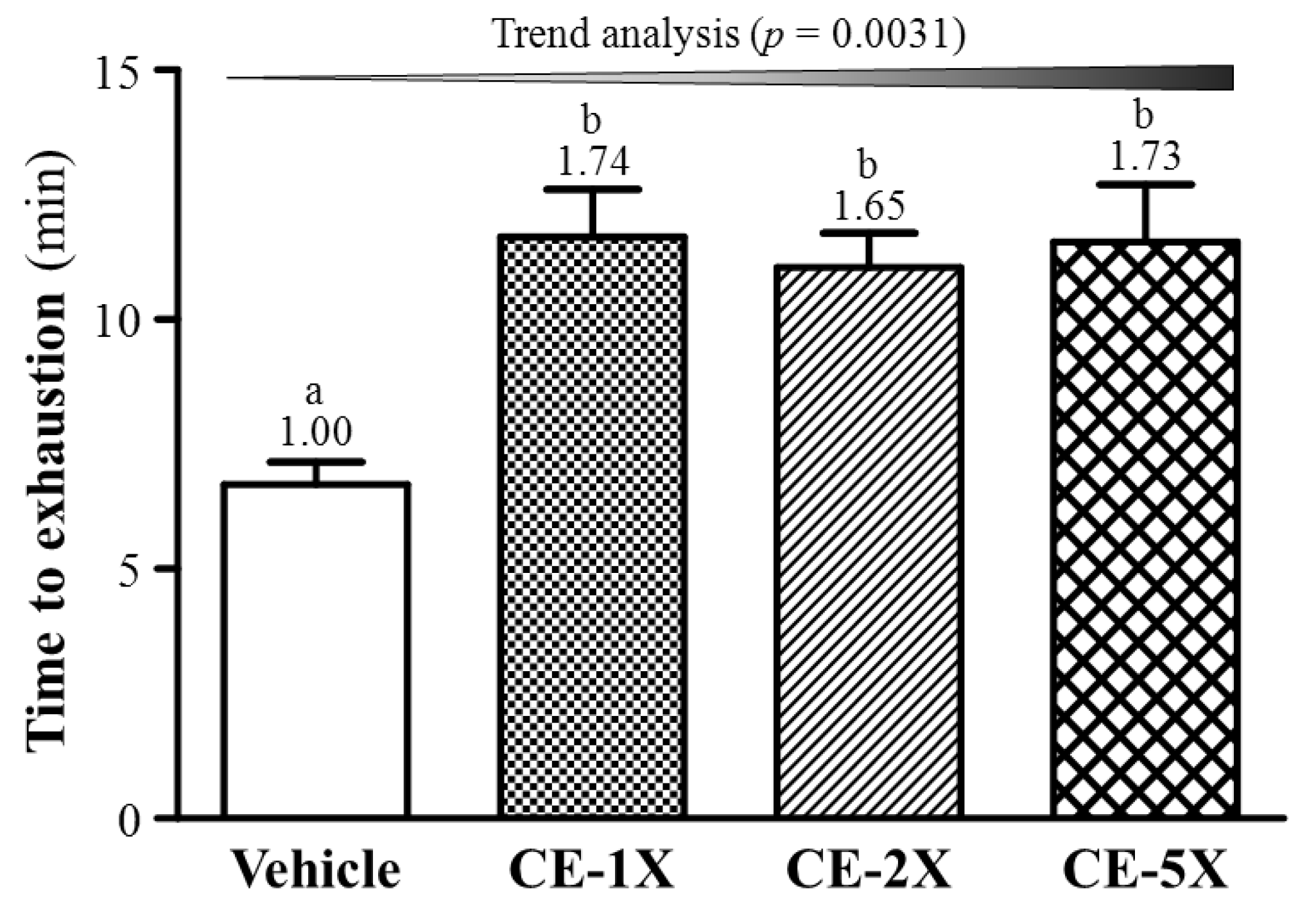{kind=link}
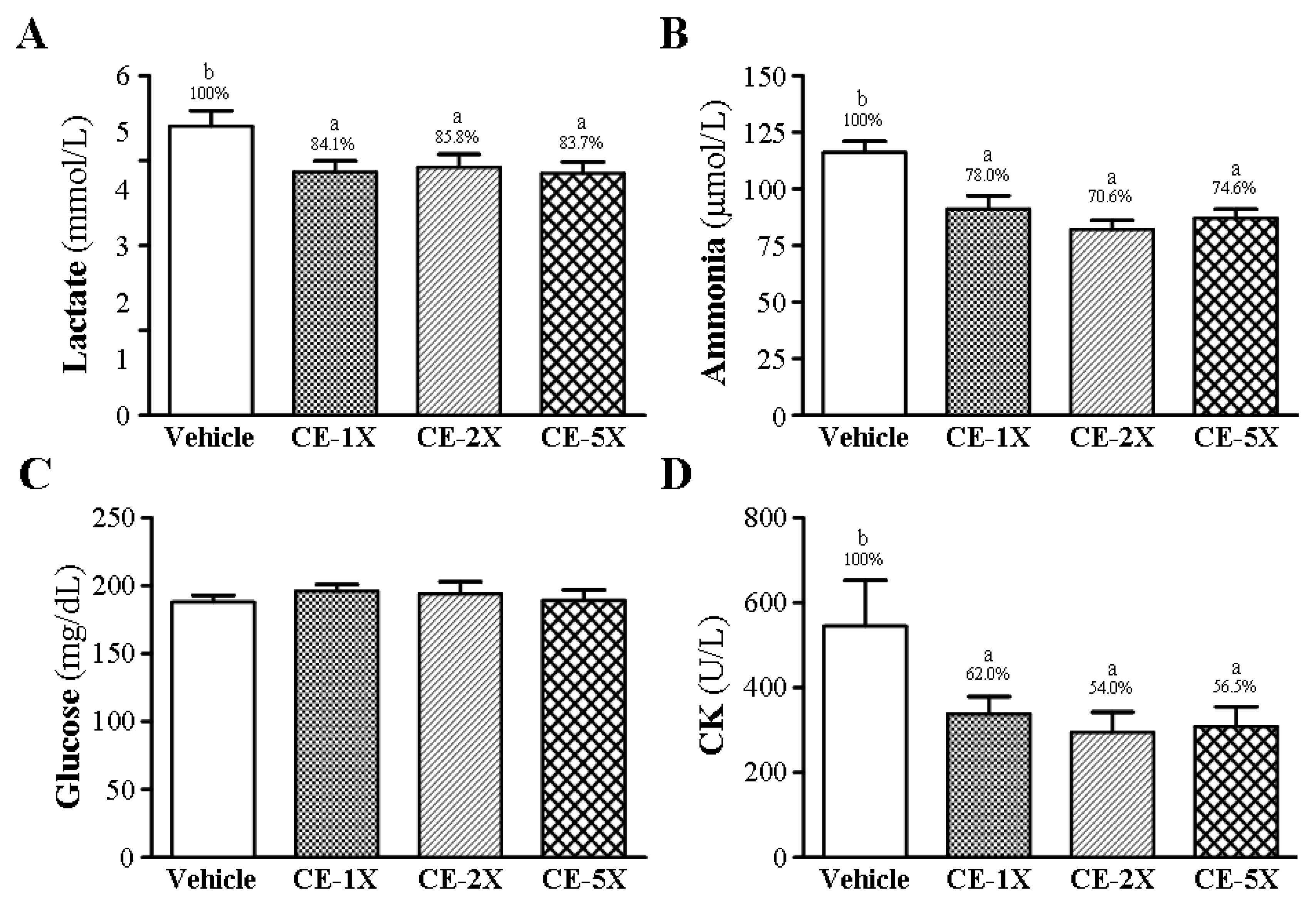{kind=link}
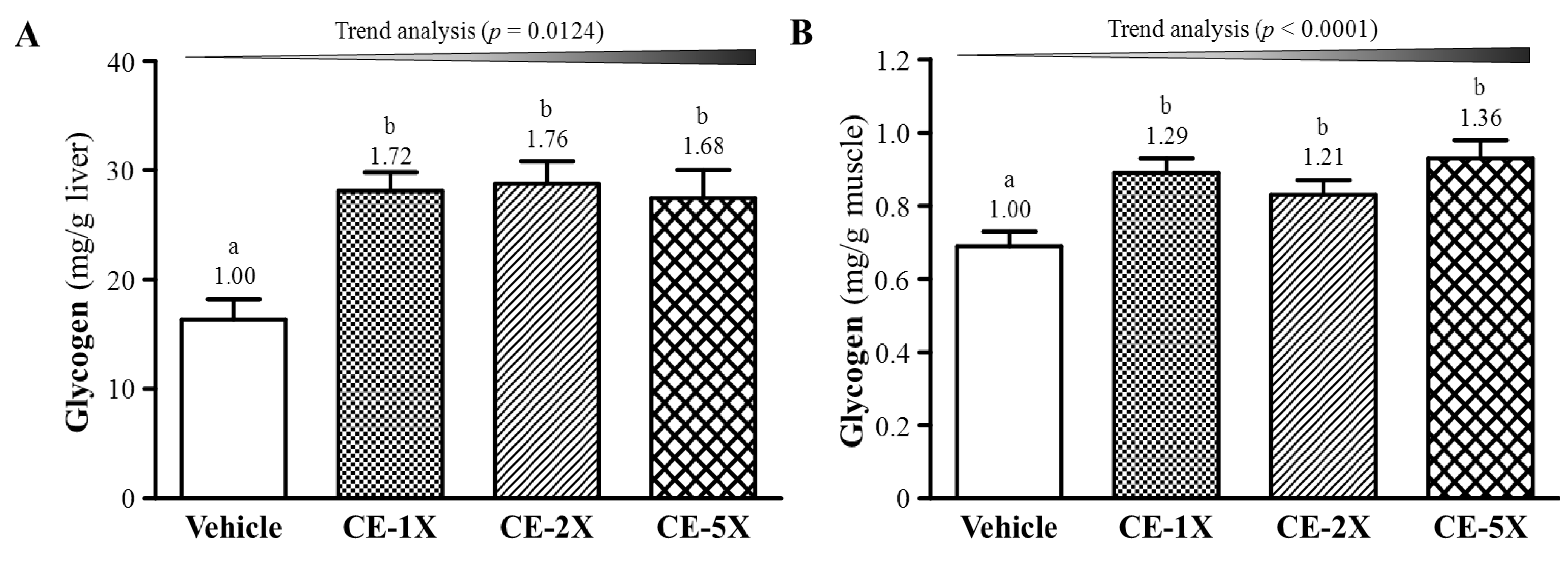{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Vehicle | CE-1X | CE-2X | CE-5X | Trend Analysis p Value |
|---|---|---|---|---|---|
| Food intake (g/mouse/day) | 6.3 ± 0.1 b | 6.4 ± 0.0 bc | 6.5 ± 0.1 c | 5.9 ± 0.1 a | 0.0256 (↓) |
| Water intake (mL/mouse/day) | 7.7 ± 0.2 a | 8.1 ± 0.2 ab | 8.2 ± 0.2 b | 8.2 ± 0.2 b | 0.0133 (↑) |
| Initial BW (g) | 31.4 ± 0.2 | 31.0 ± 0.2 | 31.5 ± 0.2 | 31.0 ± 0.3 | 0.6678 |
| Final BW (g) | 37.0 ± 0.4 a | 38.3 ± 0.5 b | 38.2 ± 0.4 b | 37.5 ± 0.4 ab | 0.3178 |
| Liver (g) | 2.16 ± 0.05 | 2.17 ± 0.05 | 2.17 ± 0.04 | 2.11 ± 0.05 | 0.7186 |
| Muscle (g) | 0.38 ± 0.01 | 0.39 ± 0.01 | 0.40 ± 0.01 | 0.39 ± 0.01 | 0.2168 |
| Kidney (g) | 0.68 ± 0.01 ab | 0.67 ± 0.01 a | 0.69 ± 0.01 ab | 0.72 ± 0.02 b | 0.0603 |
| Testis (g) | 0.25 ± 0.01 | 0.24 ± 0.01 | 0.24 ± 0.01 | 0.24 ± 0.00 | 0.2719 |
| EFP (g) | 0.54 ± 0.03 | 0.54 ± 0.02 | 0.52 ± 0.04 | 0.60 ± 0.03 | 0.3662 |
| BAT (g) | 0.126 ± 0.007 | 0.124 ± 0.004 | 0.131 ± 0.005 | 0.120 ± 0.004 | 0.5540 |
| Parameters | Vehicle | CE-1X | CE-2X | CE-5X | Trend Analysis p Value |
|---|---|---|---|---|---|
| AST (U/L) | 84 ± 12 | 78 ± 4 | 76 ± 3 | 79 ± 5 | 0.3850 |
| ALT (U/L) | 67 ± 10 b | 47 ± 3 a | 56 ± 4 ab | 54 ± 6 ab | 0.8386 |
| ALP (U/L) | 82 ± 5 | 76 ± 6 | 73 ± 5 | 78 ± 4 | 0.5927 |
| LDH (U/L) | 383 ± 58 | 336 ± 33 | 375 ± 29 | 361 ± 37 | 0.5300 |
| Albumin (g/dL) | 3.6 ± 0.1 | 3.7 ± 0.1 | 3.6 ± 0.1 | 3.7 ± 0.1 | 0.2990 |
| TP (g/dL) | 4.9 ± 0.0 | 5.0 ± 0.1 | 5.0 ± 0.0 | 5.0 ± 0.1 | 0.4529 |
| BUN (mg/dL) | 20.8 ± 0.9 a | 20.2 ± 0.3 a | 23.3 ± 0.8 b | 25.5 ± 0.4 c | <0.0001 (↑) |
| Creatinine (mg/dL) | 0.16 ± 0.01 a | 0.28 ± 0.01 b | 0.35 ± 0.02 c | 0.51 ± 0.03 d | <0.0001 (↑) |
| UA (mg/dL) | 1.9 ± 0.1 b | 1.4 ± 0.1 a | 1.5 ± 0.1 a | 1.3 ± 0.1 a | <0.0001 (↓) |
| TC (mg/dL) | 109 ± 5 | 105 ± 4 | 117 ± 6 | 108 ± 5 | 0.2260 |
| TG (mg/dL) | 110 ±6 | 118 ± 9 | 119 ± 8 | 112 ± 5 | 0.7562 |
| Glucose (mg/dL) | 175 ± 5 | 187 ± 6 | 185 ± 5 | 179 ± 3 | 0.6398 |
| CK (U/L) | 369 ± 171 | 363 ± 96 | 302 ± 65 | 376 ± 56 | 0.0989 |


3. Discussion
4. Experimental
4.1. Materials, Animals, and Experiment Design
| Nutrition Facts | Content |
|---|---|
| Nutrition Facts | /60 mL CE/Serving |
| Protein | 3.9 g |
| Fat | 0 |
| Saturated fat | 0 |
| Trans fat | 0 |
| Carbohydrate | 0 |
| Sodium | 0.0364 g |
| Total calories | 15.6 Kcal |
| Hydrolyzed amino acid profiles | mg/100 g |
| Aspartic Acid | 322.9 |
| Threonine | 127.4 |
| Serine | 164.9 |
| Glutamic acid | 654.0 |
| Glycine | 1105.0 |
| Alanine | 464.8 |
| Cystine | 8.6 |
| Valine | 123.9 |
| Methionine | 49.4 |
| Isoleucine | 79.1 |
| Leucine | 180.2 |
| Tyrosine | 183.4 |
| Phenylalanine | 97.5 |
| Lysine | 248.9 |
| Histidine | 142.0 |
| Arginine | 395.6 |
| Proline | 613.7 |
| Total BCAA | mg/60 g CE |
| Valine, leucine and isoleucine | 443.0 |
4.2. Swimming Exercise Performance Test
4.3. Forelimb Grip Strength
4.4. Determination of Fatigue-Associated Biochemical Variables
4.5. Clinical Biochemical Profiles
4.6. Tissue Glycogen Determination and Visceral Organ Weight
4.7. Histology Staining of Tissues
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Supplementary Information

Conflicts of Interest
References
- Zain, A.M.; Syedsahiljamalulail, S. Effect of taking chicken essence on stress and cognition of human volunteers. Malays. J. Nutr. 2003, 9, 19–29. [Google Scholar]
- Nagai, H.; Harada, M.; Nakagawa, M.; Tanaka, T.; Gunadi, B.; Setiabudi, M.L.; Uktolseja, J.L.; Miyata, Y. Effects of chicken extract on the recovery from fatigue caused by mental workload. Appl. Hum. Sci. 1996, 15, 281–286. [Google Scholar] [CrossRef]
- Lv, Y.Q.; He, R.R.; Watanabe, H.; Abe, K.; Sakurai, E.; Yanai, K.; Kurihara, H. Effects of a chicken extract on food-deprived activity stress in rats. Biosci. Biotechnol. Biochem. 2010, 74, 1276–1278. [Google Scholar] [CrossRef]
- Kurihara, H.; Yao, X.; Nagai, H.; Tsuruoka, N.; Shibata, H.; Kiso, Y.; Fukami, H. Anti-stress effect of BRAND’s essence of chicken (BEC) on plasma glucose levels in mice loaded with restraint stress. J. Health Sci. 2006, 52, 252–258. [Google Scholar] [CrossRef]
- Man, Y.C.; Yee, C.W.; Shing, W.K.; Lai, T.P.; Ching, W.K.; Kei, K.K. The enhancing effects of a chicken-meat extract on serum Ig concentrations in normal and scalded animals. Br. J. Nutr. 2005, 94, 51–55. [Google Scholar] [CrossRef]
- Chao, J.; Tseng, H.; Chang, C.; Chien, Y.; Au, H.; Chen, J.; Chen, C. Chicken extract affects colostrum protein compositions in lactating women. J. Nutr. Biochem. 2004, 15, 37–44. [Google Scholar] [CrossRef]
- Matsumura, Y.; Kita, S.; Ono, H.; Kiso, Y.; Tanaka, T. Preventive effect of a chicken extract on the development of hypertension in stroke-prone spontaneously hypertensive rats. Biosci. Biotechnol. Biochem. 2002, 66, 1108–1110. [Google Scholar] [CrossRef]
- Geissler, C.; Boroumand-Naini, M.; Harada, M.; Iino, T.; Hirai, K.; Suwa, Y.; Tanaka, T.; Iwata, S. Chicken extract stimulates haemoglobin restoration in iron deficient rats. Int. J. Food Sci. Nutr. 1996, 47, 351–360. [Google Scholar]
- Suzuki, Y.; Nakao, T.; Maemura, H.; Sato, M.; Kamahara, K.; Morimatsu, F.; Takamatsu, K. Carnosine and anserine ingestion enhances contribution of nonbicarbonate buffering. Med. Sci. Sports Exerc. 2006, 38, 334–338. [Google Scholar]
- Wu, T.; Watanabe, H.; Hong, L.K.; Abe, K.; Ni, Y.; Fu, Z. Effect of BRAND’s essence of chicken on the resetting process of circadian clocks in rats subjected to experimental jet lag. Mol. Biol. Rep. 2011, 38, 1533–1540. [Google Scholar] [CrossRef]
- Xu, M.; He, R.R.; Zhai, Y.J.; Abe, K.; Kurihara, H. Effects of carnosine on cyclophosphamide-induced hematopoietic suppression in mice. Am. J. Chin. Med. 2014, 42, 131–142. [Google Scholar] [CrossRef]
- Li, Y.F.; He, R.R.; Tsoi, B.; Kurihara, H. Bioactivities of chicken essence. J. Food Sci. 2012, 77, 105–110. [Google Scholar]
- Zwarts, M.J.; Bleijenberg, G.; van Engelen, B.G. Clinical neurophysiology of fatigue. Clin. Neurophysiol. 2008, 119, 2–10. [Google Scholar] [CrossRef]
- Fukuda, K.; Straus, S.E.; Hickie, I.; Sharpe, M.C.; Dobbins, J.G.; Komaroff, A.; International Chronic Fatigue Syndrome Study Group. The chronic fatigue syndrome: A comprehensive approach to its definition and study. Ann. Intern. Med. 1994, 121, 953–959. [Google Scholar] [CrossRef]
- Xu, C.; Sim, M. Effect of oral feeding of essence of chicken on the level of 5-hydroxyindoie acetic acid in the cerebrospinal fluid of the rat. Int. J. Food Sci. Nutr. 1997, 48, 113–117. [Google Scholar] [CrossRef]
- Lo, H.I.; Tsi, D.; Tan, A.C.; Wang, S.W.; Hsu, M.C. Effects of postexercise supplementation of chicken essence on the elimination of exercise-induced plasma lactate and ammonia. Chin. J. Physiol. 2005, 48, 187–192. [Google Scholar]
- Chen, C.C.; Ringenbach, D.R.; Snow, M. Treadmill walking effects on grip strength in young men with Down syndrome. Res. Dev. Disabil. 2014, 35, 288–293. [Google Scholar]
- Yamano, E.; Tanaka, M.; Ishii, A.; Tsuruoka, N.; Abe, K.; Watanabe, Y. Effects of chicken essence on recovery from mental fatigue in healthy males. Med. Sci. Monit. 2013, 19, 540–547. [Google Scholar] [CrossRef]
- Huang, C.C.; Hsu, M.C.; Huang, W.C.; Yang, H.R.; Hou, C.C. Triterpenoid-rich extract from Antrodia camphorata improves physical fatigue and exercise performance in mice. Evid. Based Complement. Alternat. Med. 2012. [Google Scholar] [CrossRef]
- Wang, S.Y.; Huang, W.C.; Liu, C.C.; Wang, M.F.; Ho, C.S.; Huang, W.P.; Hou, C.C.; Chuang, H.L.; Huang, C.C. Pumpkin (Cucurbita moschata) fruit extract improves physical fatigue and exercise performance in mice. Molecules 2012, 17, 11864–11876. [Google Scholar]
- Leenders, M.; Verdijk, L.B.; van der Hoeven, L.; van Kranenburg, J.; Nilwik, R.; Wodzig, W.K.; Senden, J.M.; Keizer, H.A.; van Loon, L.J. Protein supplementation during resistance-type exercise training in the elderly. Med. Sci. Sports. Exerc. 2013, 45, 542–552. [Google Scholar] [CrossRef]
- Bandy, W.D.; Lovelace-Chandler, V.; McKitrick-Bandy, B. Adaptation of skeletal muscle to resistance training. J. Orthop. Sports Phys. Ther. 1990, 12, 248–255. [Google Scholar] [CrossRef]
- Funes, L.; Carrera-Quintanar, L.; Cerdán-Calero, M.; Ferrer, M.D.; Drobnic, F.; Pons, A.; Roche, E.; Micol, V. Effect of lemon verbena supplementation on muscular damage markers, proinflammatory cytokines release and neutrophils’ oxidative stress in chronic exercise. Eur. J. Appl. Physiol. 2011, 111, 695–705. [Google Scholar] [CrossRef]
- Jackman, S.R.; Witard, O.C.; Jeukendrup, A.E.; Tipton, K.D. Branched-chain amino acid ingestion can ameliorate soreness from eccentric exercise. Med. Sci. Sports. Exerc. 2010, 42, 962–970. [Google Scholar] [CrossRef]
- Kohen, R.; Yamamoto, Y.; Cundy, K.C.; Ames, B.N. Antioxidant activity of carnosine, homocarnosine, and anserine present in muscle and brain. Proc. Natl. Acad. Sci. USA 1988, 85, 3175–3179. [Google Scholar]
- Wu, R.E.; Huang, W.C.; Liao, C.C.; Chang, Y.K.; Kan, N.W.; Huang, C.C. Resveratrol protects against physical fatigue and improves exercise performance in mice. Molecules 2013, 18, 4689–4702. [Google Scholar] [CrossRef]
- Cairns, S.P. Lactic acid and exercise performance: Culprit or friend? Sports Med. 2006, 36, 279–291. [Google Scholar] [CrossRef]
- Prado, E.S.; de Rezende Neto, J.M.; de Almeida, R.D.; Dória de Melo, M.G.; Cameron, L.C. Keto analogue and amino acid supplementation affects the ammonaemia response during exercise under ketogenic conditions. Br. J. Nutr. 2011, 105, 1729–1733. [Google Scholar] [CrossRef]
- Wilkinson, D.J.; Smeeton, N.J.; Watt, P.W. Ammonia metabolism, the brain and fatigue; revisiting the link. Prog. Neurobiol. 2010, 91, 200–219. [Google Scholar] [CrossRef]
- Krook, A.; Wallberg-Henriksson, H.; Zierath, J.R. Sending the signal: Molecular mechanisms regulating glucose uptake. Med. Sci. Sports Exerc. 2004, 36, 1212–1217. [Google Scholar] [CrossRef]
- Fujii, N.; Jessen, N.; Goodyear, L.J. AMP-activated protein kinase and the regulation of glucose transport. Am. J. Physiol. Endocrinol. Metab. 2006, 291, 867–877. [Google Scholar] [CrossRef]
- Manabe, Y.; Miyatake, S.; Takagi, M.; Nakamura, M.; Okeda, A.; Nakano, T.; Hirshman, M.F.; Goodyear, L.J.; Fujii, N.L. Characterization of an acute muscle contraction model using cultured C2C12 myotubes. PLoS One 2012, 7, e52592. [Google Scholar]
- Smith, A.E.; Stout, J.R.; Kendall, K.L.; Fukuda, D.H.; Cramer, J.T. Exercise-induced oxidative stress: The effects of β-alanine supplementation in women. Amino Acids 2012, 43, 77–90. [Google Scholar] [CrossRef]
- Kan, N.W.; Huang, W.C.; Lin, W.T.; Huang, C.Y.; Wen, K.C.; Chiang, H.M.; Huang, C.C.; Hsu, M.C. Hepatoprotective effects of Ixora parviflora extract against exhaustive exercise-induced oxidative stress in mice. Molecules 2013, 18, 10721–10732. [Google Scholar] [CrossRef]
- Skenderi, K.P.; Kavouras, S.A.; Anastasiou, C.A.; Yiannakouris, N.; Matalas, A.L. Exertional Rhabdomyolysis during a 246-km continuous running race. Med. Sci. Sports Exerc. 2006, 38, 1054–1057. [Google Scholar]
- Sullivan, M.A.; Aroney, S.T.; Li, S.; Warren, F.J.; Joo, J.S.; Mak, K.S.; Stapleton, D.I.; Bell-Anderson, K.S.; Gilbert, R.G. Changes in glycogen structure over feeding cycle sheds new light on blood-glucose control. Biomacromolecules 2014, 15, 660–665. [Google Scholar] [CrossRef]
- Gejl, K.D.; Hvid, L.G.; Frandsen, U.; Jensen, K.; Sahlin, K.; Ortenblad, N. Muscle glycogen content modifies SR Ca2+ release rate in elite endurance athletes. Med. Sci. Sports Exerc. 2014, 46, 496–505. [Google Scholar] [CrossRef]
- Iaia, F.; Perez-Gomez, J.; Nordsborg, N.; Bangsbo, J. Effect of previous exhaustive exercise on metabolism and fatigue development during intense exercise in humans. Scand. J. Med. Sci. Sports 2010, 20, 619–629. [Google Scholar]
- Chen, W.C.; Huang, W.C.; Chiu, C.C.; Chang, Y.K.; Huang, C.C. Whey protein improves exercise performance and biochemical profiles in trained mice. Med. Sci. Sports Exerc. 2014. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, W.-C.; Lin, C.-I.; Chiu, C.-C.; Lin, Y.-T.; Huang, W.-K.; Huang, H.-Y.; Huang, C.-C. Chicken Essence Improves Exercise Performance and Ameliorates Physical Fatigue. Nutrients 2014, 6, 2681-2696. https://doi.org/10.3390/nu6072681
Huang W-C, Lin C-I, Chiu C-C, Lin Y-T, Huang W-K, Huang H-Y, Huang C-C. Chicken Essence Improves Exercise Performance and Ameliorates Physical Fatigue. Nutrients. 2014; 6(7):2681-2696. https://doi.org/10.3390/nu6072681
Chicago/Turabian StyleHuang, Wen-Ching, Ching-I Lin, Chien-Chao Chiu, Yi-Ting Lin, Wei-Kai Huang, Hui-Yu Huang, and Chi-Chang Huang. 2014. "Chicken Essence Improves Exercise Performance and Ameliorates Physical Fatigue" Nutrients 6, no. 7: 2681-2696. https://doi.org/10.3390/nu6072681