Hepcidin and Iron Homeostasis during Pregnancy

Abstract
:1. Introduction



2. Methods
3. Results and Discussion

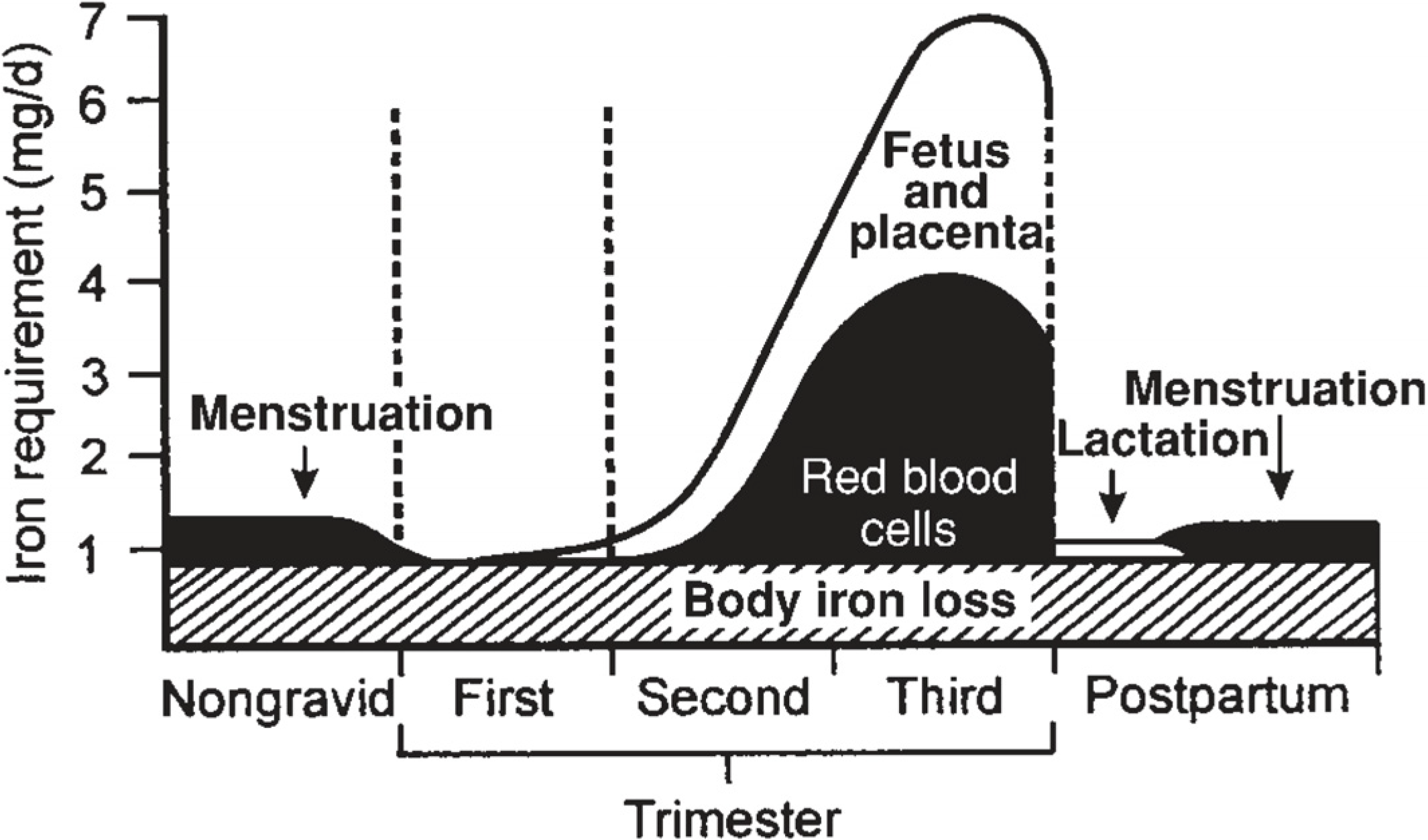{kind=link}
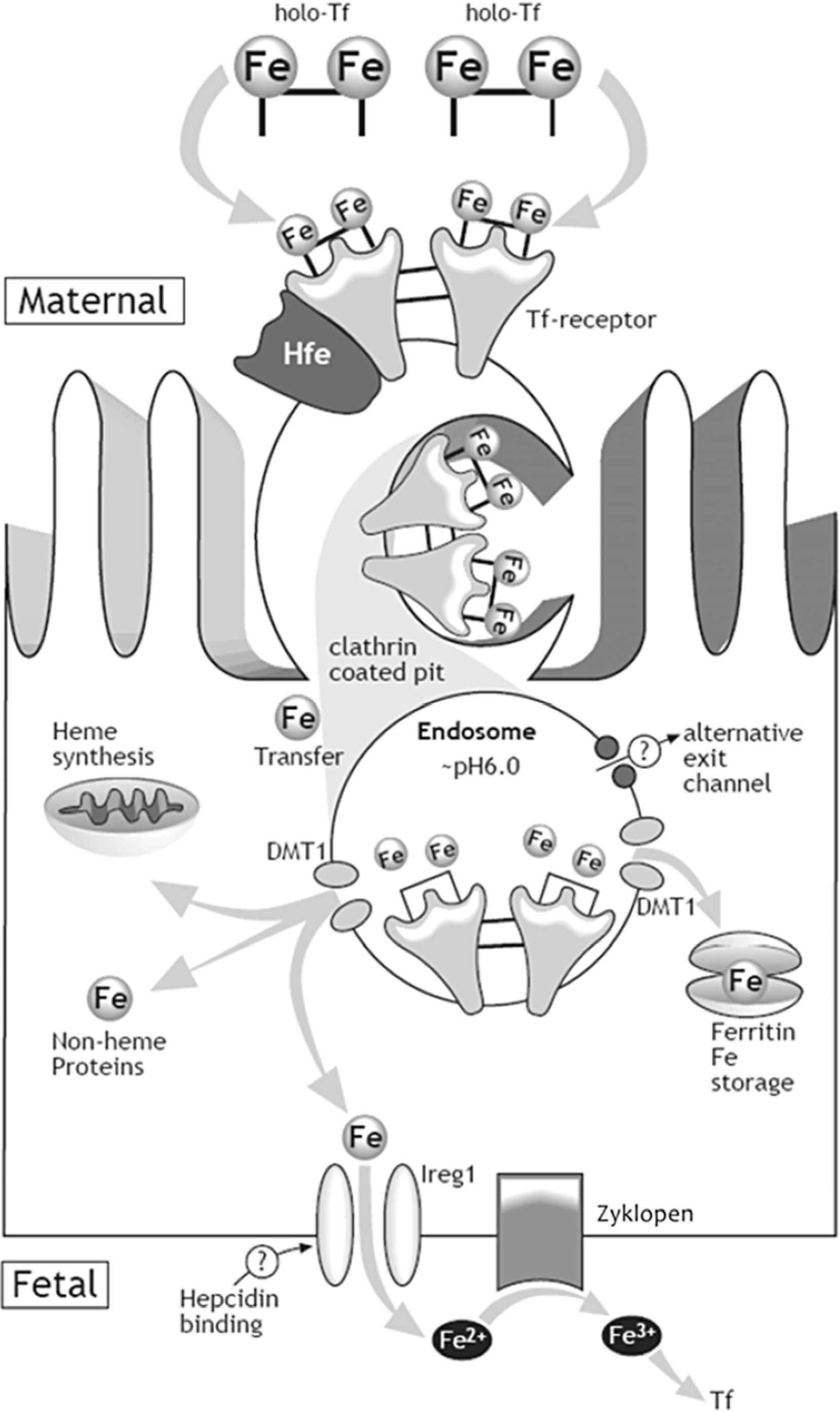{kind=link}
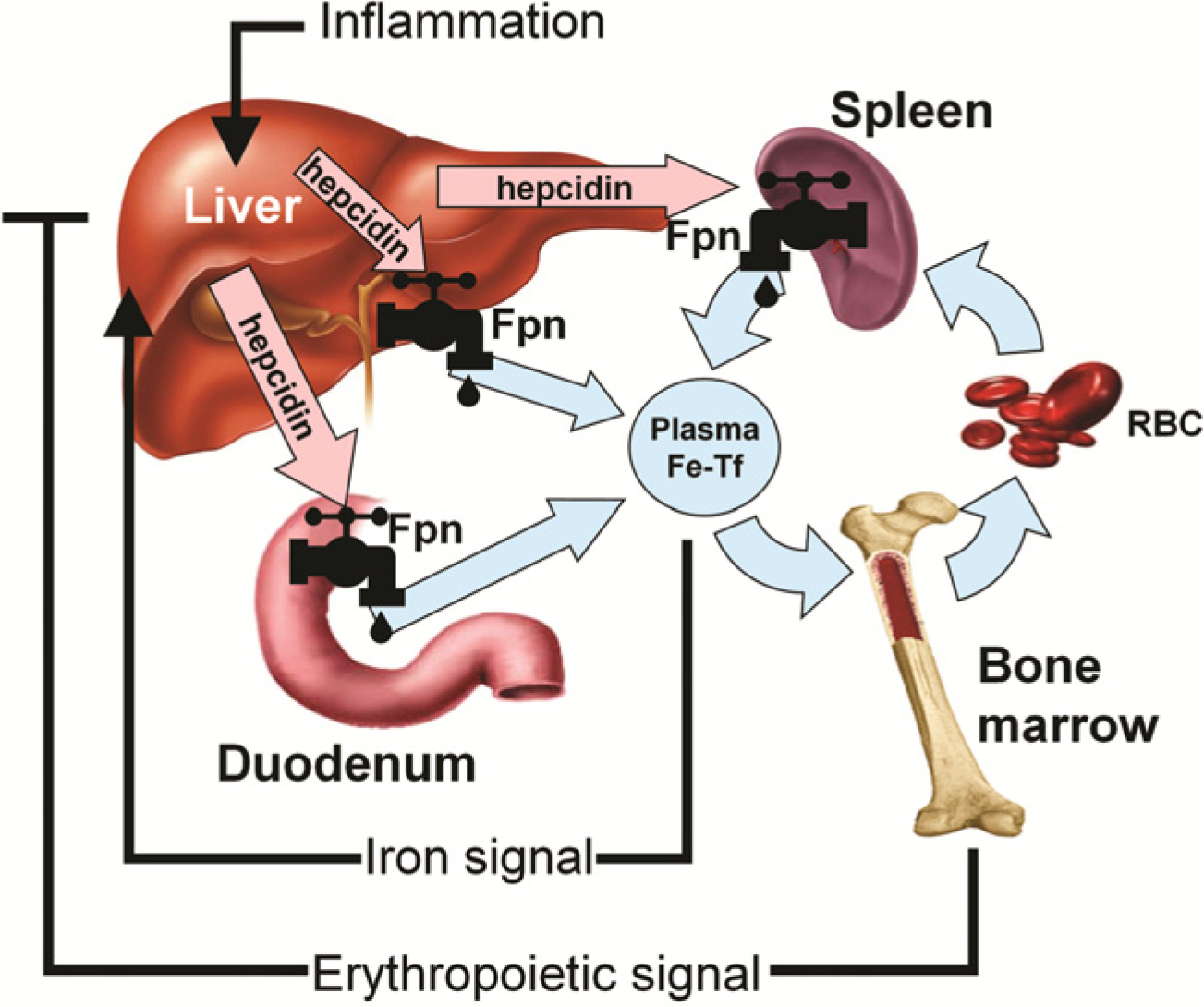{kind=link}
| Author | Design | Sample | Hepcidin | Main Study Findings |
|---|---|---|---|---|
| Human Studies | ||||
| Finkenstedt et al. (2012) [54] | Longitudinal, prospective | n = 42 Health status: 38 healthy and 4 with complications (preeclampsia: n = 1, GDM: n = 1, C/S: n = 1, preeclampsia and GDM: n = 1). Location: NDR Maternal age (years): 26.4 (range 17–40) BMI (kg/m2): NDR Parity: primigravida: n = 7, previous pregnancy mean = 1.6 Race/Ethnicity: NDR Inclusion/Exclusion criteria: NDR | Sample: Maternal serum Timing: 1st and 3rd trimester (n = 42); 2nd trimester (n = 12) Method: SELDI time-of-flight mass spectrometry [63] Hepcidin reference range: average for healthy women 50 ng/mL (range < 10–200 ng/mL) | Maternal serum hepcidin: 1st trimester: median = 16 ng/mL (4–97); 2nd trimester: median = 11 ng/mL (6–36); 3rd trimester: median = 9.5 ng/mL (1–43), p < 0.001 Correlations of maternal hepcidin with iron and inflammatory parameters: Serum iron (r = 0.391, p < 0.001), ferritin (r = 0.573, p < 0.001), Tsat (r = 0.457; p < 0.001), sHJV (r = −0.231; p = 0.025), EPO (r = −0.308; p = 0.002). |
| van Santen et al. (2013) [55] | Longitudinal, prospective | n = 31 Health status: healthy except for complications in 3 (10%) Location: The Netherlands Maternal age (years): 33.1 ± 5.4 BMI (kg/m2): NDR Parity 0 = 13 (42%), Parity 1 = 11 (36%), Parity ≥ 2 = 7 (23%) Race/Ethnicity: Northern European (n = 29) or Southeast Asian (n = 2) Inclusion criteria: Normal hematologic blood count, renal function and liver enzymes at first visit | Sample: Maternal serum Timing: 9–15, 19–25 and 29–35 weeks gestation; within 24 h postpartum; 6 weeks post-delivery Method: combination of weak cation exchange chromatography and time-of-flight mass spectrometry [64] lower limit of detection: 0.25 nmol/L Hepcidin reference range: median 2.0 nmol/L (range < 0.5–12.3 nmol/L) | Maternal serum hepcidin: 15–19 weeks gestation: 1.85 nmol/L (1.10–4.10); 20–25 weeks gestation: 0.25 nmol/L (0.25–1.20); 29–35 weeks gestation: 0.25 (0.25–0.25, undetectable); 24 h postpartum: 3.0 nmol/L (0.66–9.22); 6 weeks post-delivery: 1.35 nmol/L (0.73–2.40) Correlations of maternal hepcidin with iron and inflammatory parameters: Serum ferritin (r2 = 0.516, p < 0.0001), serum iron (r2 = 0.1, p = ns), Hgb (r2 = 0.275, p = 0.015), TIBC (r2 = −0.483, p < 0.0001), sTfR (r2 = −0.293, p = 0.008), sTfR-index (r2 = −0.452, p < 0.001), and Tsat (r2 = 0.243, p = 0.029), CRP (r2 = 0.322, p = ns) |
| Dao et al. (2013) [56] | Longitudinal, prospective | n = 30 Health status: (n = 15 Lean; n = 15 Obese) GDM (Obese, n = 1; Lean, n = 1) OB complication: C/S (Obese, n = 7; Lean, n = 4) Location: Boston, MA, USA Maternal age (years): Obese: 30.0 ± 3.9; lean 32.1 ± 5.8 BMI (kg/m2): Obese = 38.6 ± 7.0; Lean = 22.8 ± 1.5 Parity: NDR Race/Ethnicity: Obese: Caucasian n = 6; AA n = 6; Hispanic n = 2; Asian n = 1; Lean: Caucasian n = 9; AA n = 0; Hispanic n = 5; Asian n = 1 Exclusion Criteria: type 2 diabetes, preeclampsia, autoimmune disease, acute infection, premature membrane rupture and chorioamnionitis | Sample: Maternal serum and cord blood Timing: 24–28 weeks gestation (maternal sample); childbirth (cord blood) Method: ELISA (Bachem Group, Torrance, CA, USA) Hepcidin reference range: NDR | Maternal serum hepcidin: (24–28 weeks gestation): Obese: 13.5 ± 9.0 ng/mL; Lean: 5.1 ± 2.7 ng/mL (p < 0.01). Correlated with maternal BMI: r = 0.4, p = 0.04 Maternal inflammatory status (24–28 weeks gestation): CRP: Obese: 14.3 (IQR: 11.5) mg/L; Lean: 5.0 (IQR: 4.4) mg/L (p < 0.01) CRP: Obese: 14.3 (IQR: 11.5) mg/L; Lean: 5.0 (IQR: 4.4) mg/L (p < 0.01) Maternal iron status (24–28 weeks gestation) (extrapolated from Figure 1): Serum iron: Obese: 60 μg/dL; Lean: 78 μg/dL Tsat: Obese: 15% Lean: 19% Cord blood hepcidin (Childbirth) (extrapolated from Figure 1): Obese: 125 ng/mL; Lean: 120 ng/mL Cord blood iron status parameters Serum iron: Obese: 97.3 ± 29.9 μg/dL; Lean: 147.7 ± 21.7 μg/dL (p < 0.01); Tsat: Obese: 39.6%; Lean: 63.5% (p = 0.01) Correlations of maternal parameters and cord blood iron status markers: Maternal BMI and cord blood iron: r = −0.8, p = 0.002; Maternal BMI and cord blood Tsat: r = −0.7, p = 0.009; Log maternal hepcidin and cord blood iron: r = −0.6, p = 0.02; Log maternal hepcidin and cord blood Tsat: r = −0.6, p = 0.02 |
| Gyarmati et al. (2011) [53] | Longitudinal, prospective | n = 38 Health status: healthy; elective C/S n = 13 Location: Hungary Maternal age: NDR BMI: NDR Parity: NDR Race/ethnicity: NDR Exclusion criteria: presence of infection, multiple birth, and any known OB complication | Sample: Maternal serum Timing: median 40 (39–41) weeks gestation; at first contraction for vaginal delivery, or before anesthesia for elective C/S, and then 3 days PP for all Method: mass spectrometry [50] Hepcidin reference range: NDR | Maternal serum hepcidin: Vaginal: before birth 2.52 ng/mL (2.07–3.1); 3 days PP 7.36 ng/mL(6.34–8.91) (p < 0.001); Elective caesarian delivery: before birth 2.83 ng/mL (2.53–3.87); 3 days PP 17.5 ng/mL (13.5–18.9) (p < 0.001) Correlations of maternal hepcidin with iron and inflammatory parameters: NDR; no significant correlation with iron, ferritin and IL-6. |
| van Santen et al. (2011) [51] | Cross-sectional, retrospective | n = 69 Health status: stratified by group:
Maternal age (years): range 16–19 BMI (kg/m2): NDR Parity: primigravid Race/Ethnicity: NDR Exclusion criteria: Primigravid without peripheral parasitemia, live singleton birth, no signs or symptoms of systemic infection | Sample: Maternal plasma and cord blood Timing: childbirth Method: combination of weak cation-exchange chromatography and time-of-flight mass spectrometry Hepcidin reference range: 4.2 nmol/L (0.5–13.9) nmol/L [65] | Maternal plasma hepcidin (Childbirth):
Correlations of maternal hepcidin with iron and inflammatory parameters: NDR |
| Young et al. (2012) [52] | Correlational, cross-sectional, prospective | n = 19 Health status: healthy Location: Rochester, NY, USA Maternal age (years): mean = 19.0 ± 2.9 (range 16–32) BMI (kg/m2): pre-pregnancy 24.7 [7.0] (20.7–43.6); delivery BMI: 30.4 [7.1] (24.4–47.5) Parity: mean = 0 ± 0.9 (0–3) Race/Ethnicity: African American 53%, Caucasian 47% Inclusion criteria: Healthy, nonsmoking, uncomplicated pregnancy Exclusion criteria: gestational diabetes, underlying malabsorption, or medical problems that affect Fe homeostasis | Sample: Maternal serum and cord blood Timing: mean = 39.9 ± 1.6 weeks (36–41.6) gestation; childbirth (maternal sample collected at admission) Method: ELISA Hepcidin reference range: median = 65 ng/mL (5%–95% range 17 to 286) [66] | Maternal serum hepcidin (Childbirth): Median [SD] = 9.30 μg/L [50.1] Cord blood hepcidin (Childbirth): Median [SD] = 61.7 μg/L [77.0], p < 0.05 Correlations of maternal hepcidin with iron and inflammatory parameters: ferritin (μg/L) (r2 = 0.59, p = 0.0001), TBI (mg/kg) (r2 = 0.59, p = 0.0001), Hgb (g/L) (r2 = 0.31, p = 0.01) and TfR (mg/L) (r2 = 0.31, p = 0.01)] Correlations of cord blood hepcidin with neonatal iron and inflammatory parameters: ferritin (r2 = 0.60, p < 0.0001) and TBI (r2 = 0.60, p = 0.0001) Maternal serum hepcidin did not significantly correlate with neonatal serum hepcidin. Pregnant women with undetectable levels of serum hepcidin transferred a greater quantity of the maternally ingested 57Fe-nonheme or 58Fe-heme iron to their fetus compared to women with detectable levels of serum hepcidin (p = 0.003 and 0.002) |
| Rehu et al. (2010) [48] | Cross-sectional, prospective, correlational | n = 191 Health status: vaginal delivery: n = 147; elective C/S: n = 24; emergency C/S: n = 20; gestational diabetes mellitus: n = 23 gestational hypertension: n = 6; preeclampsia: n = 5; liver dysfunction: n = 2; three groups: 1. iron-restricted erythropoiesis; 2. sufficient iron for erythropoiesis, but low iron stores; 3. normal iron stores and sufficient iron for erythropoiesis Location: Finland Maternal age (years): median = 28 (range 17–41) BMI (kg/m2): NDR Parity: NDR Race/Ethnicity: NDR Inclusion/Exclusion criteria: NDR | Sample: Maternal serum and cord blood Timing: median = 40 + 1 weeks (37–42) gestation; selected at time of delivery <24 h before delivery Method: ELISA Hepcidin reference range: median 65 ng/mL 5%–95% range 17 to 286 [66] | Maternal serum hepcidin (Childbirth): Overall geometric mean = 12.4 ng/mL (95% CI = 10.5–14.6); vaginal geometric mean = 13.6 (95% CI = 11.0–17.0); emergency CS geometric mean = 15.9 (95% CI = 8.8–28.7); elective CS geometric mean = 5.5 (95% CI = 3.2–9.5), significantly lower than vaginal or emergency CS; iron-restricted erythropoiesis = 3.8 ng/mL (1.5–9.6); low iron stores = 6.7 ng/mL (4.2–10.6); normal iron stores = 15.2 ng/mL (11.1–20.7), p = 0.002 Correlations of maternal hepcidin with iron and inflammatory parameters: CRP (r = 0.285, p < 0.001) Cord blood hepcidin (Childbirth): geometric mean = 71.6 ng/mL (95% CI = 60.8–84.4), p < 0.001; None of the maternal measurements were associated with the cord blood hepcidin |
| Toldi et al. (2010) [50] | Cross-sectional, descriptive | n = 67 Health status: 30 preeclamptic and 37 healthy Location: Hungary Maternal age (years): median = 30 BMI (kg/m2): NDR Parity: NDR Race/Ethnicity: NDR Inclusion/Exclusion criteria: not reported; routine perinatal oral iron supplementation: all participants. | Sample: Maternal plasma Timing: 24–40 weeks gestation; preeclamptic median = 36.5 (24–40) weeks gestation; healthy median = 36 (28–39) weeks gestation Method: mass spectrometry (LC-MS/MS) [36] | Maternal plasma hepcidin 24–40 weeks gestation:Preeclampsia: 5.68 ng/mL (0.72–9.25) Healthy: 3.74 ng/mL (0.73–8.14) (p = 0.003) Correlations of maternal hepcidin with iron and inflammatory parameters: NDR |
| Schulze et al. (2008) [49] | Correlational, cross sectional prospective. | n = 190 Health status: purposive sampling of women with iron deficiency Location: Bangladesh Maternal age: mean = 21.9 (17.5–25) years BMI (kg/m2): NDR Parity: 48% primigravida; 52% multigravida Race/Ethnicity: NDR Inclusion/Exclusion criteria: not reported | Sample: Maternal urine Timing: mean (SD) = 12 (8–14) weeks gestation; following pregnancy confirmation Method: surface-enhanced laser desorption/ionization time-of-flight mass spectrometry Hepcidin reference range: 0.09–2.97 [67] | Maternal urine hepcidin 8-14 weeks gestation: median = 2.35 (intensity/mmol creatinine) (0.51–7.22) Correlations of maternal hepcidin with iron and inflammatory parameters: ferritin (μg/L) (r = 0.33, p < 0.001) & TfR index (TfR/log ferritin) (r = −0.22, p = 0.007); no correlation with soluble TfR, hemoglobin, EPO; log AGP (mg/dL) (r = 0.20, p = 0.01); no correlation CRP |
| Howard et al. (2007) [47] | Correlational, cross-sectional, prospective | n = 31 Health status: + Plasmodium Falciparum malaria Location: Ghana Maternal age (years): mean = 24.4 BMI (kg/m2): NDR Parity: NDR Race/Ethnicity: NDR Inclusion criteria: diagnosed with P. Falciparum malaria, not admitted for transfusion, had a hemoglobin level >50 g/L, and no cerebral malaria | Sample: Maternal urine Timing: NDR Method: time-of-flight mass spectrometry Hepcidin reference range: 0.09–2.97 intensity/mmol creatinine [67] | Maternal urine hepcidin: mean = 0.7 (intensity/mmolcreatinine) (0.2, 5.9) Correlations of maternal hepcidin with iron and inflammatory parameters: not associated with hemoglobin or anemia, but 90% of women were anemic; log parasitemia positively associated (β = 0.218; CI = 0.043–0.392, p = 0.016) |
| Animal Studies | ||||
| Cornock et al. (2013) [61] | Prospective, cross-sectional | n = 40 Animal: virgin Wistar and Rowett Hooded Lister rats Age: NDR Diet: Dams fed either
| Sample: maternal and fetal liver Timing: day 21of gestation, placentas and livers dissected and frozen Method: mRNA by real-time RT-PCR | Maternal liver hepcidin: mRNA hepcidin expression decreased in rats fed low iron diet (p < 0.001), no difference by strain Fetal liver hepcidin: mRNA hepcidin expression higher in Wistar compared to Rowett Hooded Lister (43%, p < 0.001), and was decreased by exposure to low iron diet in both strains (61%, p < 0.001) |
| Sozo et al. (2013) [62] | n = 13 Animal: Border-Leicester × Merino ewes Groups:
| Sample: fetal liver Timing: gestational day 134 (term is ~day 145) Method: mRNA by real-time RT-PCR | Fetal liver hepcidin: Gene expression lower in ethanol-exposed liver (0.2 ± 0.1) compared to control (1 ± 0.2), p = 0.011 | |
| Neves et al. (2010) [58] | Prospective, cross sectional | n = 13 Animal: mice Groups:
Age: one year old nulliparous and pluriparous (mean = 29 weaned pups per female) Diet: fed standard diet with iron content of 312 mg/kg | Sample: female liver Timing: 1 year old. For pluriparous group, the sample was collected on average 11 weeks after last delivery Method: hepcidin 1 and 2 mRNA by real-time RT-PCR | Hepcidin 1 and 2: lower expression of hepatic hepcidin mRNA in pluriparous mice (of both genotypes) compared to nulliparous (p < 0.01) |
| Gambling et al. (2009) [59] | Experimental, correlational, prospective | n = 104 Animal: virgin Rowett Hooded Lister Rats Groups:
| Sample: maternal and fetal liver Timing: fetuses delivered by C/S on day 21.5 of gestation, Method: real time quantitative PCR from total RNA from liver | Maternal liver hepcidin: mRNA hepcidin expression decreased in iron deficient group (p < 0.001), hepcidin restored to near control levels by iron supplementation in the 2nd half but not 1st half of pregnancy (p < 0.05) Fetal liver hepcidin: Low in fetuses from iron deficient group (p < 0.001); fetal liver iron and maternal liver hepcidin (r2 = 0.59, p < 0.001); fetal liver iron and hepcidin expression in fetal liver (r2 = 0.47, p < 0.001) |
| Millard et al. (2004) [60] | Cross-sectional, Prospective | n = 4 per time point Animal: Sprague-Dawley rats Age: mated at 10–12 weeks, those not pregnant used as control Diet: All fed standard rodent pellet diet (370 mg iron/kg) | Sample: maternal liver Timing: 5 time points: 9, 15, 18, and 21 days gestation, and 24–48 h PP Method: hepatic samples by ribonuclease protection assay | Maternal hepatic hepcidin mRNA: Progressive decline through gestation down to 1.9% of the non-pregnant level on day 21 (p < 0.05); Postpartum levels normalized |
3.1. Maternal Hepcidin Levels through Pregnancy
3.2. Maternal Hepcidin and Iron Status
3.3. Maternal Hepcidin and Inflammation
3.4. Hepcidin and Delivery Type
3.5. Relationship between Maternal and Neonate Cord Blood Hepcidin and Iron Status
4. Limitations
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Andrews, N.C. Disorders of iron metabolism. N. Engl. J. Med. 1999, 341, 1986–1995. [Google Scholar] [CrossRef]
- National Research Council. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; National Academy Press: Washington, DC, USA, 2001; p. 772.
- Greer, J.P.; Ovid Technologies Inc. Wintrobe’s Clinical Hematology, 12th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2009. [Google Scholar]
- Corbett, J.V. Accidental poisoning with iron supplements. MCN Am. J. Matern. Child Nurs. 1995, 20, 234. [Google Scholar] [CrossRef]
- Bothwell, T.H. Iron requirements in pregnancy and strategies to meet them. Am. J. Clin. Nutr. 2000, 72, 257S–264S. [Google Scholar]
- Allen, L.H. Biological mechanisms that might underlie iron’s effects on fetal growth and preterm birth. J. Nutr. 2001, 131, 581S–589S. [Google Scholar]
- Scott, D.E. Anemia in pregnancy. Obstet. Gynecol. Annu. 1972, 1, 219–244. [Google Scholar]
- Tapiero, H.; Gate, L.; Tew, K.D. Iron: Deficiencies and requirements. Biomed. Pharmacother. 2001, 55, 324–332. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Recommendations to prevent and control iron deficiency in the United States. MMWR Recomm. Rep. 1998, 47, 1–29. [Google Scholar]
- Reshetnikova, O.S.; Burton, G.J.; Teleshova, O.V. Placental histomorphometry and morphometric diffusing capacity of the villous membrane in pregnancies complicated by maternal iron-deficiency anemia. Am. J. Obstet. Gynecol. 1995, 173, 724–727. [Google Scholar] [CrossRef]
- Kadyrov, M.; Kosanke, G.; Kingdom, J.; Kaufmann, P. Increased fetoplacental angiogenesis during first trimester in anaemic women. Lancet 1998, 352, 1747–1749. [Google Scholar] [CrossRef]
- Konijn, A.M. Iron metabolism in inflammation. Baillieres Clin. Haematol. 1994, 7, 829–849. [Google Scholar] [CrossRef]
- Godfrey, K.M.; Redman, C.W.; Barker, D.J.; Osmond, C. The effect of maternal anaemia and iron deficiency on the ratio of fetal weight to placental weight. Br. J. Obstet. Gynecol. 1991, 98, 886–891. [Google Scholar] [CrossRef]
- Collins, J.F.; Wessling-Resnick, M.; Knutson, M.D. Hepcidin regulation of iron transport. J. Nutr. 2008, 138, 2284–2288. [Google Scholar] [CrossRef]
- Cogswell, M.E.; Parvanta, I.; Ickes, L.; Yip, R.; Brittenham, G.M. Iron supplementation during pregnancy, anemia, and birth weight: A randomized controlled trial. Am. J. Clin. Nutr. 2003, 78, 773–781. [Google Scholar]
- Blot, I.; Diallo, D.; Tchernia, G. Iron deficiency in pregnancy: Effects on the newborn. Curr. Opin. Hematol. 1999, 6, 65–70. [Google Scholar] [CrossRef]
- Earl, R.O.; Woteki, C.E.; Institute of Medicine (U.S.); Committee on the Prevention Detection and Management of Iron Deficiency Anemia among U.S.; Children and Women of Childbearing Age; Institute of Medicine (U.S.); Food and Nutrition Board. Iron Deficiency Anemia: Recommended Guidelines for the Prevention, Detection, and Management among U.S. Children and Women of Childbearing Age; National Academy Press: Washington, DC, USA, 1993; p. 6. [Google Scholar]
- Allen, L.H. Pregnancy and iron deficiency: Unresolved issues. Nutr. Rev. 1997, 55, 91–101. [Google Scholar] [CrossRef]
- Malhotra, M.; Sharma, J.B.; Batra, S.; Sharma, S.; Murthy, N.S.; Arora, R. Maternal and perinatal outcome in varying degrees of anemia. Int. J. Gynaecol. Obstet. 2002, 79, 93–100. [Google Scholar] [CrossRef]
- Allen, L.H. Anemia and iron deficiency: Effects on pregnancy outcome. Am. J. Clin. Nutr. 2000, 71, 1280S–1284S. [Google Scholar]
- Gulmezoglu, A.M.; Mahomed, K.; Hofmeyr, G.J.; Nikodem, V.C.; Kramer, T. Fetal and maternal catecholamine levels at delivery. J. Perinat. Med. 1996, 24, 687–691. [Google Scholar] [CrossRef]
- Campos, M.S.; Barrionuevo, M.; Alferez, M.J.; Gomez-Ayala, A.E.; Rodriguez-Matas, M.C.; Lopez Aliaga, I.; Lisbona, F. Interactions among iron, calcium, phosphorus and magnesium in the nutritionally iron-deficient rat. Exp. Physiol. 1998, 83, 771–781. [Google Scholar]
- Coghetto Baccin, A.; Lauerman Lazzaretti, L.; Duarte Martins Brandao, V.; Manfredini, V.; Peralba, M.C.; Silveira Benfato, M. Oxidative stress in older patients with iron deficiency anaemia. J. Nutr. Health Aging 2009, 13, 666–670. [Google Scholar] [CrossRef]
- Casanueva, E.; Viteri, F.E. Iron and oxidative stress in pregnancy. J. Nutr. 2003, 133, 1700S–1708S. [Google Scholar]
- Knutson, M.D.; Handelman, G.J.; Viteri, F.E. Methods for measuring ethane and pentane in expired air from rats and humans. Free Radic. Biol. Med. 2000, 28, 514–519. [Google Scholar] [CrossRef]
- Walter, P.B.; Knutson, M.D.; Paler-Martinez, A.; Lee, S.; Xu, Y.; Viteri, F.E.; Ames, B.N. Iron deficiency and iron excess damage mitochondria and mitochondrial DNA in rats. Proc. Natl. Acad. Sci. USA 2002, 99, 2264–2269. [Google Scholar]
- Bhaskaram, P. Immunobiology of mild micronutrient deficiencies. Br. J. Nutr. 2001, 85 (Suppl. 2), S75–S80. [Google Scholar]
- McArdle, H.J.; Lang, C.; Hayes, H.; Gambling, L. Role of the placenta in regulation of fetal iron status. Nutr. Rev. 2011, 69 (Suppl. 1), S17–S22. [Google Scholar] [CrossRef]
- Paterson, S.; Armstrong, N.J.; Iacopetta, B.J.; McArdle, H.J.; Morgan, E.H. Intravesicular pH and iron uptake by immature erythroid cells. J. Cell. Physiol. 1984, 120, 225–232. [Google Scholar] [CrossRef]
- Harris, E.D. New insights into placental iron transport. Nutr. Rev. 1992, 50, 329–331. [Google Scholar]
- Donovan, A.; Lima, C.A.; Pinkus, J.L.; Pinkus, G.S.; Zon, L.I.; Robine, S.; Andrews, N.C. The iron exporter ferroportin/Slc40a1 is essential for iron homeostasis. Cell Metab. 2005, 1, 191–200. [Google Scholar]
- Chen, H.; Attieh, Z.K.; Syed, B.A.; Kuo, Y.M.; Stevens, V.; Fuqua, B.K.; Andersen, H.S.; Naylor, C.E.; Evans, R.W.; Gambling, L.; et al. Identification of zyklopen, a new member of the vertebrate multicopper ferroxidase family, and characterization in rodents and human cells. J. Nutr. 2010, 140, 1728–1735. [Google Scholar] [CrossRef]
- Ganz, T. Hepcidin—A regulator of intestinal iron absorption and iron recycling by macrophages. Best Pract. Res. Clin. Haematol. 2005, 18, 171–182. [Google Scholar] [CrossRef]
- Ganz, T. Hepcidin and its role in regulating systemic iron metabolism. Hematol. Am. Soc. Hematol. Educ. Progr. 2006. [Google Scholar] [CrossRef]
- Nemeth, E.; Ganz, T. Regulation of iron metabolism by hepcidin. Annu. Rev. Nutr. 2006, 26, 323–342. [Google Scholar] [CrossRef]
- Murphy, A.T.; Witcher, D.R.; Luan, P.; Wroblewski, V.J. Quantitation of hepcidin from human and mouse serum using liquid chromatography tandem mass spectrometry. Blood 2007, 110, 1048–1054. [Google Scholar] [CrossRef]
- Ganz, T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood 2003, 102, 783–788. [Google Scholar] [CrossRef]
- Darshan, D.; Anderson, G.J. Interacting signals in the control of hepcidin expression. Biometals 2009, 22, 77–87. [Google Scholar] [CrossRef]
- Hamlin, F.; Latunde-Dada, G.O. Iron bioavailibity from a tropical leafy vegetable in anaemic mice. Nutr. Metab. 2011, 8, 9. [Google Scholar] [CrossRef]
- Goodnough, L.T.; Nemeth, E.; Ganz, T. Detection, evaluation, and management of iron-restricted erythropoiesis. Blood 2010, 116, 4754–4761. [Google Scholar] [CrossRef]
- Gagliardo, B.; Kubat, N.; Faye, A.; Jaouen, M.; Durel, B.; Deschemin, J.C.; Canonne-Hergaux, F.; Sari, M.A.; Vaulont, S. Pro-hepcidin is unable to degrade the iron exporter ferroportin unless maturated by a furin-dependent process. J. Hepatol. 2009, 50, 394–401. [Google Scholar]
- Sasu, B.J.; Li, H.; Rose, M.J.; Arvedson, T.L.; Doellgast, G.; Molineux, G. Serum hepcidin but not prohepcidin may be an effective marker for anemia of inflammation (AI). Blood Cells Mol. Dis. 2010, 45, 238–245. [Google Scholar] [CrossRef]
- Roe, M.A.; Spinks, C.; Heath, A.L.; Harvey, L.J.; Foxall, R.; Wimperis, J.; Wolf, C.; Fairweather-Tait, S.J. Serum prohepcidin concentration: No association with iron absorption in healthy men; and no relationship with iron status in men carrying HFE mutations, hereditary haemochromatosis patients undergoing phlebotomy treatment, or pregnant women. Br. J. Nutr. 2007, 97, 544–549. [Google Scholar] [CrossRef]
- Briana, D.D.; Boutsikou, T.; Baka, S.; Boutsikou, M.; Stamati, L.; Hassiakos, D.; Gourgiotis, D.; Malamitsi-Puchner, A. Perinatal role of hepcidin and iron homeostasis in full-term intrauterine growth-restricted infants. Eur. J. Haematol. 2012, 90, 37–44. [Google Scholar]
- Simavli, S.; Derbent, A.U.; Uysal, S.; Turhan, N.O. Hepcidin, iron status, and inflammation variables among healthy pregnant women in the turkish population. J. Matern. Fetal Neonatal Med. 2014, 27, 75–79. [Google Scholar] [CrossRef]
- Itkonen, O.; Parkkinen, J.; Stenman, U.H.; Hamalainen, E. Preanalytical factors and reference intervals for serum hepcidin LC-MS/MS method. Clin. Chim. Acta 2012, 413, 696–701. [Google Scholar] [CrossRef]
- Howard, C.T.; McKakpo, U.S.; Quakyi, I.A.; Bosompem, K.M.; Addison, E.A.; Sun, K.; Sullivan, D.; Semba, R.D. Relationship of hepcidin with parasitemia and anemia among patients with uncomplicated plasmodium falciparum malaria in ghana. Am. J. Trop. Med. Hyg. 2007, 77, 623–626. [Google Scholar]
- Rehu, M.; Punnonen, K.; Ostland, V.; Heinonen, S.; Westerman, M.; Pulkki, K.; Sankilampi, U. Maternal serum hepcidin is low at term and independent of cord blood iron status. Eur. J. Haematol. 2010, 85, 345–352. [Google Scholar] [CrossRef]
- Schulze, K.J.; Christian, P.; Ruczinski, I.; Ray, A.L.; Nath, A.; Wu, L.S.; Semba, R.D. Hepcidin and iron status among pregnant women in bangladesh. Asia Pac. J. Clin. Nutr. 2008, 17, 451–456. [Google Scholar]
- Toldi, G.; Stenczer, B.; Molvarec, A.; Takats, Z.; Beko, G.; Rigo, J., Jr.; Vasarhelyi, B. Hepcidin concentrations and iron homeostasis in preeclampsia. Clin. Chem. Lab. Med. 2010, 48, 1423–1426. [Google Scholar]
- Van Santen, S.; de Mast, Q.; Luty, A.J.; Wiegerinck, E.T.; van der Ven, A.J.; Swinkels, D.W. Iron homeostasis in mother and child during placental malaria infection. Am. J. Trop. Med. Hyg. 2011, 84, 148–151. [Google Scholar] [CrossRef]
- Merhi, Z.O.; Seifer, D.B.; Weedon, J.; Adeyemi, O.; Holman, S.; Anastos, K.; Golub, E.T.; Young, M.; Karim, R.; Greenblatt, R.; et al. Circulating vitamin D correlates with serum antimullerian hormone levels in late-reproductive-aged women: Women’s interagency HIV study. Fertil. Steril. 2012, 98, 228–234. [Google Scholar] [CrossRef]
- Gyarmati, B.; Szabo, E.; Szalay, B.; Czuczy, N.; Toldi, G.; Cseh, A.; Vasarhelyi, B.; Takats, Z. Serum maternal hepcidin levels 3 days after delivery are higher compared to those measured at parturition. J. Obstet. Gynaecol. Res. 2011, 37, 1620–1624. [Google Scholar] [CrossRef]
- Finkenstedt, A.; Widschwendter, A.; Brasse-Lagnel, C.G.; Theurl, I.; Hubalek, M.; Dieplinger, H.; Tselepis, C.; Ward, D.G.; Vogel, W.; Zoller, H. Hepcidin is correlated to soluble hemojuvelin but not to increased GDF15 during pregnancy. Blood Cells Mol. Dis. 2012, 48, 233–237. [Google Scholar] [CrossRef]
- Van Santen, S.; Kroot, J.J.; Zijderveld, G.; Wiegerinck, E.T.; Spaanderman, M.E.; Swinkels, D.W. The iron regulatory hormone hepcidin is decreased in pregnancy: A prospective longitudinal study. Clin. Chem. Lab. Med. 2013, 51, 1395–1401. [Google Scholar]
- Dao, M.C.; Sen, S.; Iyer, C.; Klebenov, D.; Meydani, S.N. Obesity during pregnancy and fetal iron status: Is hepcidin the link? J. Perinatol. 2013, 33, 177–181. [Google Scholar] [CrossRef]
- Nicolas, G.; Bennoun, M.; Porteu, A.; Mativet, S.; Beaumont, C.; Grandchamp, B.; Sirito, M.; Sawadogo, M.; Kahn, A.; Vaulont, S. Severe iron deficiency anemia in transgenic mice expressing liver hepcidin. Proc. Natl. Acad. Sci. USA 2002, 99, 4596–4601. [Google Scholar] [CrossRef]
- Neves, J.V.; Olsson, I.A.; Porto, G.; Rodrigues, P.N. Hemochromatosis and pregnancy: Iron stores in the Hfe−/− mouse are not reduced by multiple pregnancies. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G525–G529. [Google Scholar] [CrossRef]
- Gambling, L.; Czopek, A.; Andersen, H.S.; Holtrop, G.; Srai, S.K.; Krejpcio, Z.; McArdle, H.J. Fetal iron status regulates maternal iron metabolism during pregnancy in the rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1063–R1070. [Google Scholar] [CrossRef]
- Millard, K.N.; Frazer, D.M.; Wilkins, S.J.; Anderson, G.J. Changes in the expression of intestinal iron transport and hepatic regulatory molecules explain the enhanced iron absorption associated with pregnancy in the rat. Gut 2004, 53, 655–660. [Google Scholar] [CrossRef]
- Cornock, R.; Gambling, L.; Langley-Evans, S.C.; McArdle, H.J.; McMullen, S. The effect of feeding a low iron diet prior to and during gestation on fetal and maternal iron homeostasis in two strains of rat. Reprod. Biol. Endocrinol. 2013, 11, 32. [Google Scholar] [CrossRef] [Green Version]
- Sozo, F.; Dick, A.M.; Bensley, J.G.; Kenna, K.; Brien, J.F.; Harding, R.; de Matteo, R. Alcohol exposure during late ovine gestation alters fetal liver iron homeostasis without apparent dysmorphology. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R1121–R1129. [Google Scholar] [CrossRef]
- Ward, D.G.; Roberts, K.; Stonelake, P.; Goon, P.; Zampronio, C.G.; Martin, A.; Johnson, P.J.; Iqbal, T.; Tselepis, C. SELDI-TOF-MS determination of hepcidin in clinical samples using stable isotope labelled hepcidin as an internal standard. Proteome Sci. 2008, 6, 28. [Google Scholar] [CrossRef]
- Kroot, J.J.; Laarakkers, C.M.; Geurts-Moespot, A.J.; Grebenchtchikov, N.; Pickkers, P.; van Ede, A.E.; Peters, H.P.; van Dongen-Lases, E.; Wetzels, J.F.; Sweep, F.C.; et al. Immunochemical and mass-spectrometry-based serum hepcidin assays for iron metabolism disorders. Clin. Chem. 2010, 56, 1570–1579. [Google Scholar] [CrossRef]
- Kroot, J.J.; Hendriks, J.C.; Laarakkers, C.M.; Klaver, S.M.; Kemna, E.H.; Tjalsma, H.; Swinkels, D.W. (Pre)analytical imprecision, between-subject variability, and daily variations in serum and urine hepcidin: Implications for clinical studies. Anal. Biochem. 2009, 389, 124–129. [Google Scholar] [CrossRef]
- Ganz, T.; Olbina, G.; Girelli, D.; Nemeth, E.; Westerman, M. Immunoassay for human serum hepcidin. Blood 2008, 112, 4292–4297. [Google Scholar]
- Kemna, E.H.; Tjalsma, H.; Podust, V.N.; Swinkels, D.W. Mass spectrometry-based hepcidin measurements in serum and urine: Analytical aspects and clinical implications. Clin. Chem. 2007, 53, 620–628. [Google Scholar] [CrossRef]
- Finkenstedt, A.; Bianchi, P.; Theurl, I.; Vogel, W.; Witcher, D.R.; Wroblewski, V.J.; Murphy, A.T.; Zanella, A.; Zoller, H. Regulation of iron metabolism through GDF15 and hepcidin in pyruvate kinase deficiency. Br. J. Haematol. 2009, 144, 789–793. [Google Scholar] [CrossRef]
- Blackburn, S.T. Maternal, Fetal & Neonatal Physiology: A Clinical Perspective, 2nd ed.; Saunders: St. Louis, MO, USA, 2003; p. 238. [Google Scholar]
- Weiss, G.; Goodnough, L.T. Anemia of chronic disease. N. Engl. J. Med. 2005, 352, 1011–1023. [Google Scholar] [CrossRef]
- Ganz, T.; Nemeth, E. Iron sequestration and anemia of inflammation. Semin. Hematol. 2009, 46, 387–393. [Google Scholar] [CrossRef]
- Park, K.H.; Sawada, T.; Kosuge, T.; Kita, J.; Shimoda, M.; Tomosugi, N.; Kubota, K. Surgical inflammation induces hepcidin production after abdominal surgery. World J. Surg. 2012, 36, 800–806. [Google Scholar] [CrossRef]
- Kroot, J.J.; van Herwaarden, A.E.; Tjalsma, H.; Jansen, R.T.; Hendriks, J.C.; Swinkels, D.W. Second round robin for plasma hepcidin methods: First steps toward harmonization. Am. J. Hematol. 2012, 87, 977–983. [Google Scholar] [CrossRef]
- Whittaker, P.G.; Lind, T. The intravascular mass of albumin during human pregnancy: A serial study in normal and diabetic women. Br. J. Obstet. Gynaecol. 1993, 100, 587–592. [Google Scholar] [CrossRef]
- McArdle, H.J.; Andersen, H.S.; Jones, H.; Gambling, L. Copper and iron transport across the placenta: Regulation and interactions. J. Neuroendocrinol. 2008, 20, 427–431. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Koenig, M.D.; Tussing-Humphreys, L.; Day, J.; Cadwell, B.; Nemeth, E. Hepcidin and Iron Homeostasis during Pregnancy. Nutrients 2014, 6, 3062-3083. https://doi.org/10.3390/nu6083062
Koenig MD, Tussing-Humphreys L, Day J, Cadwell B, Nemeth E. Hepcidin and Iron Homeostasis during Pregnancy. Nutrients. 2014; 6(8):3062-3083. https://doi.org/10.3390/nu6083062
Chicago/Turabian StyleKoenig, Mary Dawn, Lisa Tussing-Humphreys, Jessica Day, Brooke Cadwell, and Elizabeta Nemeth. 2014. "Hepcidin and Iron Homeostasis during Pregnancy" Nutrients 6, no. 8: 3062-3083. https://doi.org/10.3390/nu6083062