Apoptosis Inducing Effects of Kuding Tea Polyphenols in Human Buccal Squamous Cell Carcinoma Cell Line BcaCD885

Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of Kuding Tea Polyphenols
2.2. Cancer Cell Preparation
2.3. Growth Inhibition Measurement
2.4. Flow Cytometry Analysis
2.5. mRNA Expression Measurement

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene name | Sequence |
|---|---|
| Procapase-3 | Forward: 5′-CTG GAA TAT CCC TGG ACA AC-3′ |
| Reverse: 5′-CAG GTC AAC AGG TCC ATT TG-3′ | |
| Procapase-8 | Forward: 5′-CCC CAC CCT CAC TTT GCT-3′ |
| Reverse: 5′-GGA GGA CCA GGC TCA CTT A-3′ | |
| Procapase-9 | Forward: 5′-GGC CCT TCC TCG CTT CAT CTC-3′ |
| Reverse: 5′-GGT CCT TGG GCC TTC CTG GTA T-3′ | |
| Bax | Forward: 5′-AAG CTG AGC GAG TGT CTC CGG CG-3′ |
| Reverse: 5′-CAG ATG CCG GTT CAG GTA CTC AGT C-3′ | |
| Bcl-2 | Forward: 5′-CTC GTC GCT ACC GTC GTG ACT TGG-3′ |
| Reverse: 5′-CAG ATG CCG GTT CAG GTA CTC AGT C-3′ | |
| Bcl-xL | Forward: 5′-CCC AGA AAG GAT ACA GCT GG-3′ |
| Reverse: 5′-GCG ATC CGA CTC ACC AAT AC-3′ | |
| HIAP-1 | Forward: 5′-GCC TGA TGC TGG ATA ACT GG-3′ |
| Reverse: 5′-GGC GAC AGA AAA GTC AAT GG-3′ | |
| HIAP-2 | Forward: 5′-GCC TGA TGC TGG ATA ACT GG-3′ |
| Reverse: 5′-GCT CTT GCC AAT TCT GAT GG-3′ | |
| p53 | Forward: 5′-CTG AGG TTG GCT CTG ACT GTA CCA CCA TCC-3′ |
| Reverse: 5′-CTC ATT CAG CTC TCG GAA CAT CTC GAA GCG-3′ | |
| p21 | Forward: 5′-GCA GAC CAG CAT GAC AGA TTT-3′ |
| Reverse: 5′-GGA TTA GGG CTT CCT CTT GGA-3′ | |
| E2F1 | Forward: 5′-GGG GAG AAG TCA CGC TAT GA-3′ |
| Reverse: 5′-CTC AGG GCA CAG GAA AAC AT-3′ | |
| p73 | Forward: 5′-GAC GGA ATT CAC CAC CAT CCT-3′ |
| Reverse: 5′-CCA GGC TCT CTT TCA GCT TCA-3′ | |
| Fas | Forward: 5′-GAA ATG AAA TCC AAA GCT-3′ |
| Reverse: 5′-TAA TTT AGA GGC AAA GTG GC-3′ | |
| FasL | Forward: 5′-GGA TTG GGC CTG GGG ATG TTT CA-3′ |
| Reverse: 5′-TTG TGG CTC AGG GGC AGG TTG TTG-3′ | |
| GAPDH | Forward: 5′-CGG AGT CAA CGG ATT TGG TC-3′ |
| Reverse: 5′-AGC CTT CTC CAT GGT CGT GA-3′ |
2.6. Protein Expression Measurement
2.7. Statistical Analysis
3. Results
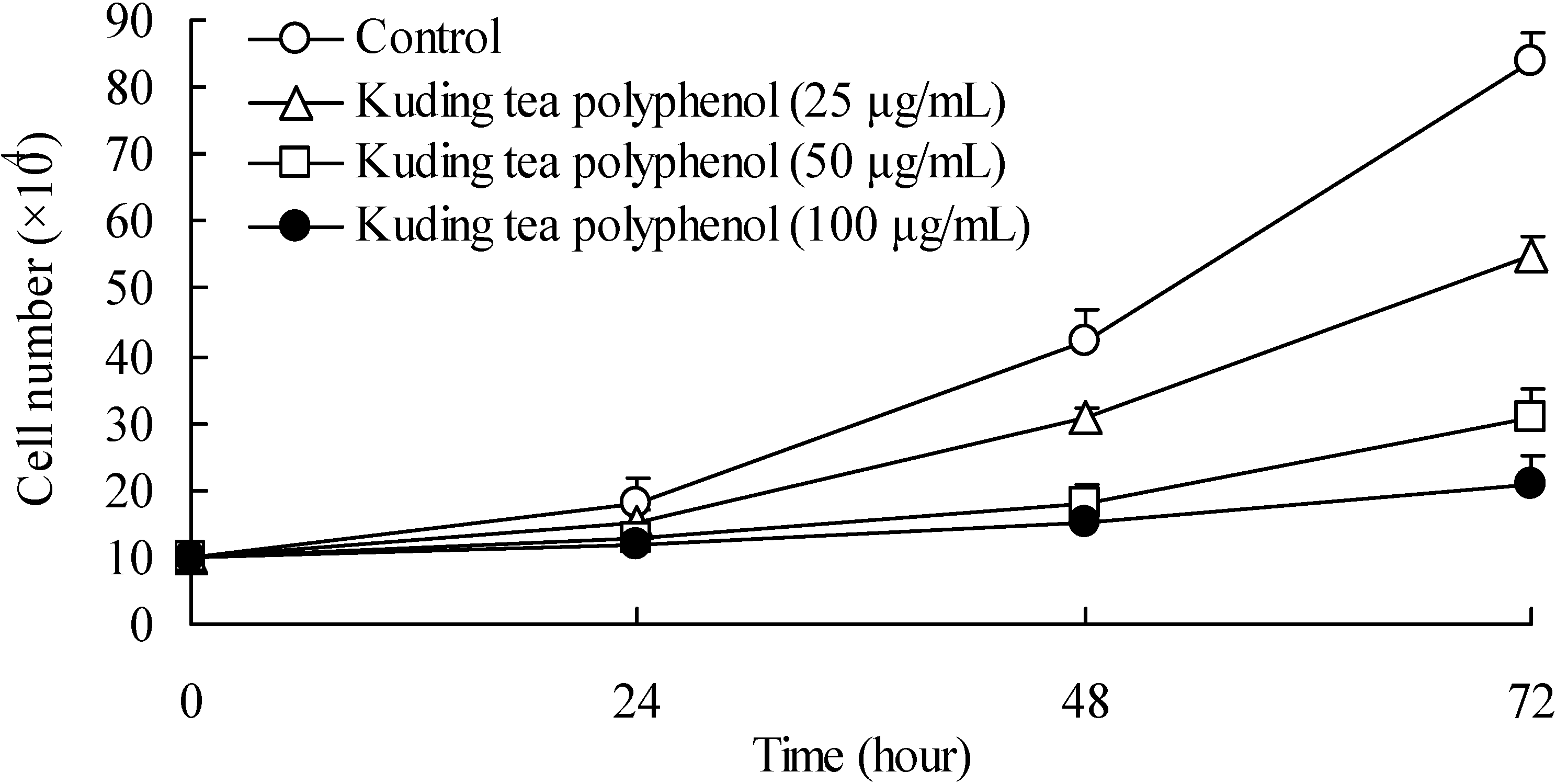
3.1. Growth Inhibitory Effects of Kuding Tea Polyphenol against BcaCD885 Cells

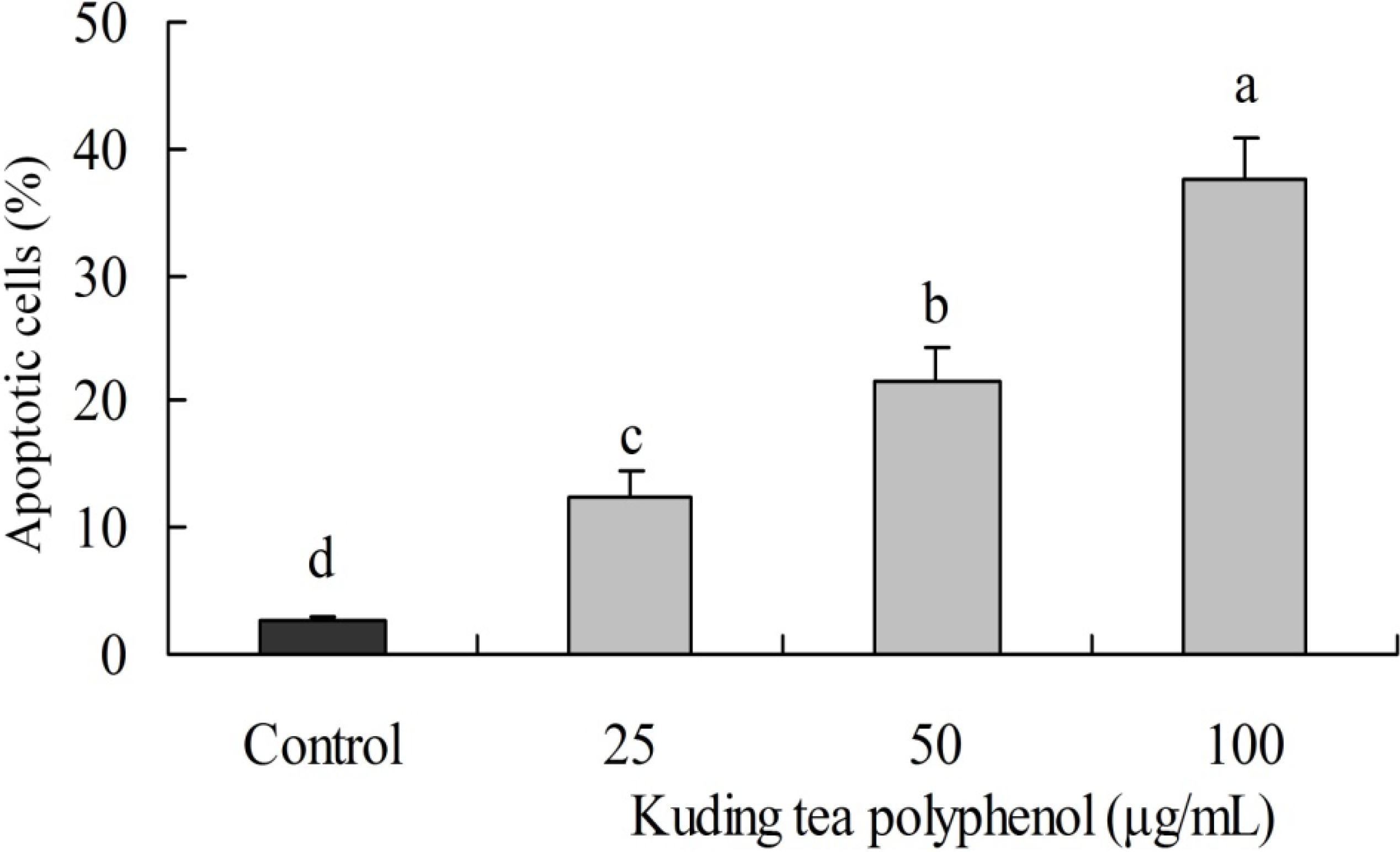
3.2. Induction of Apoptosis by Kuding Tea Polyphenol

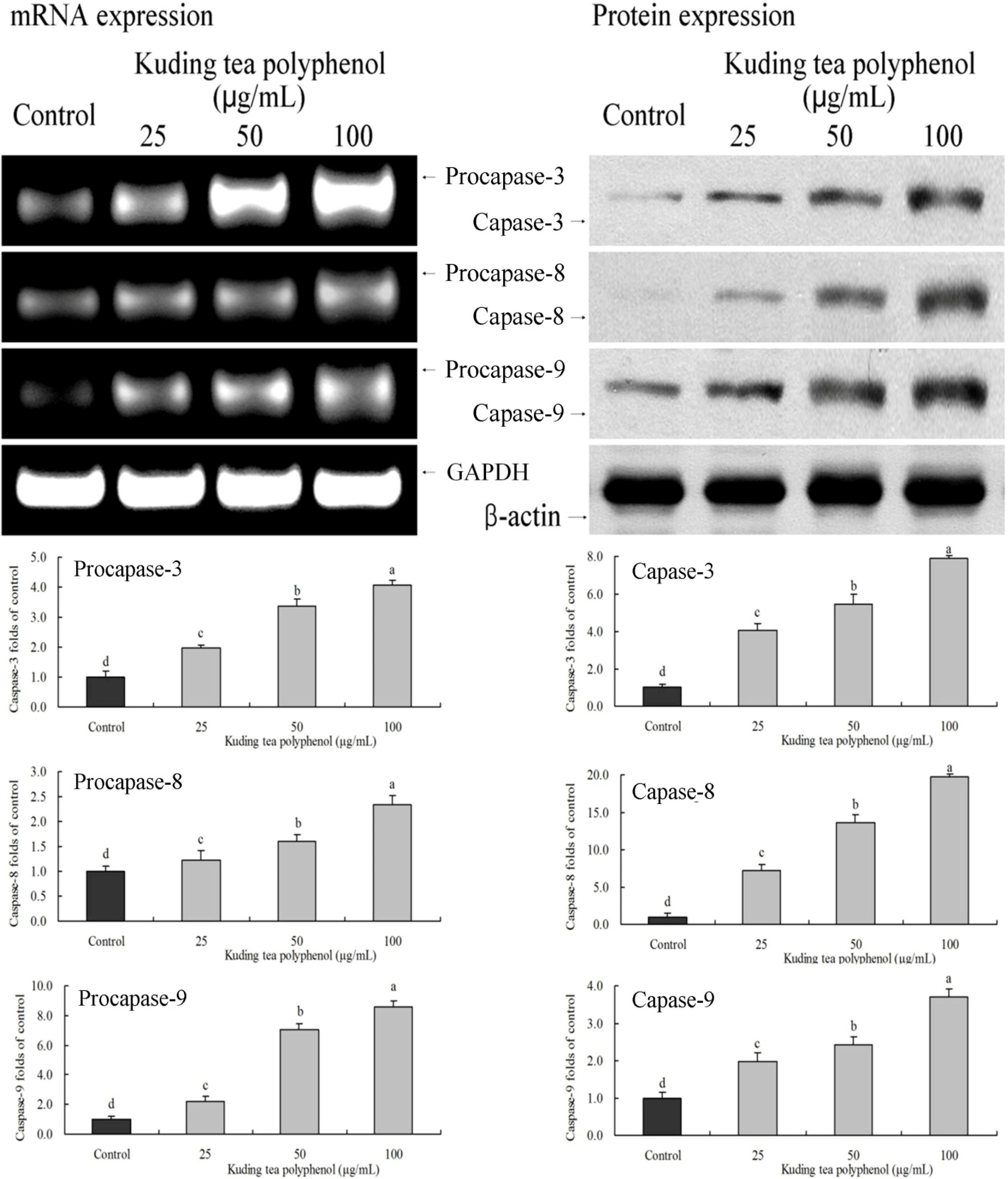
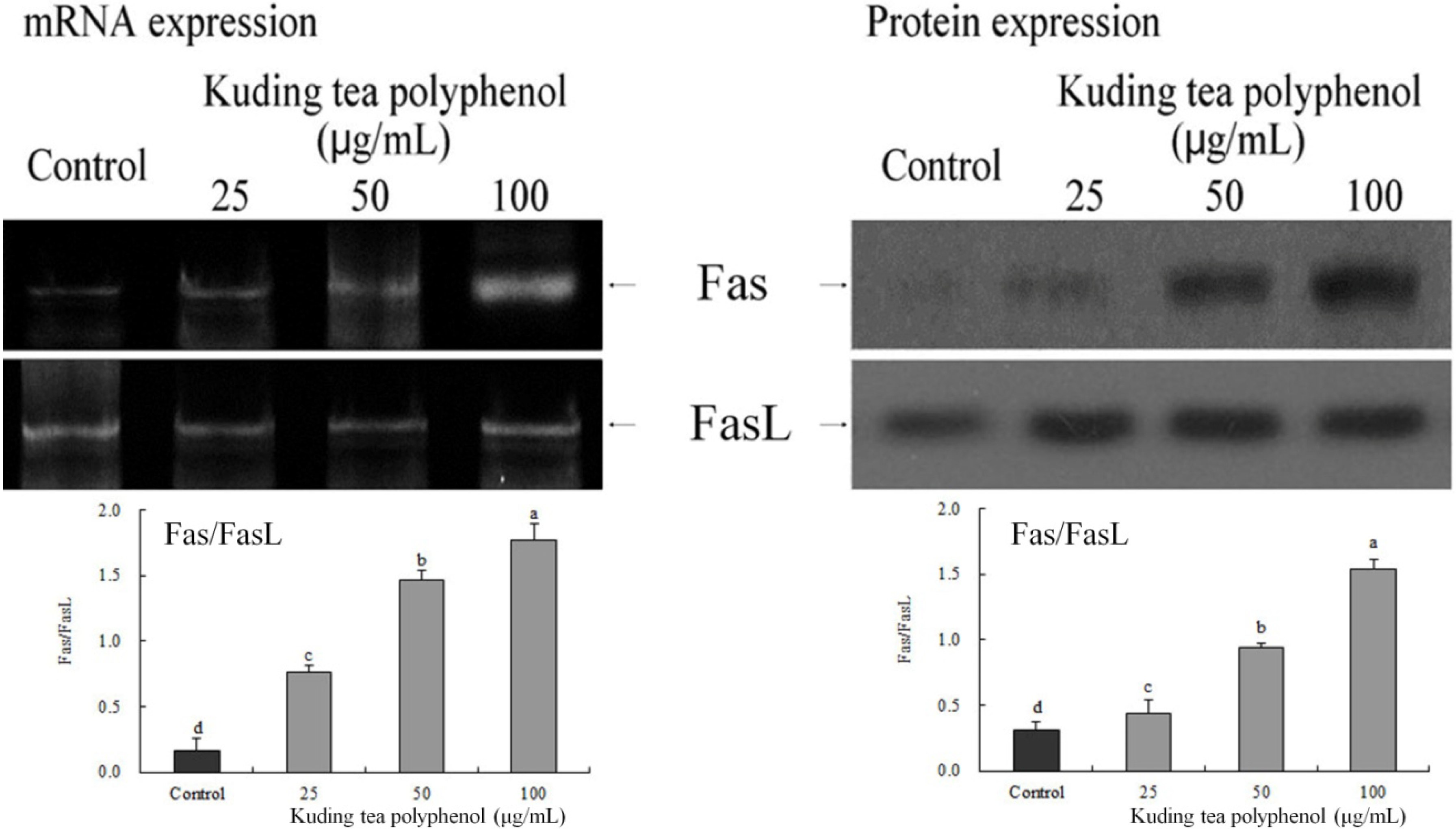
3.3. Gene Expression of the Apoptosis-Related Procapases and Fas/FasL


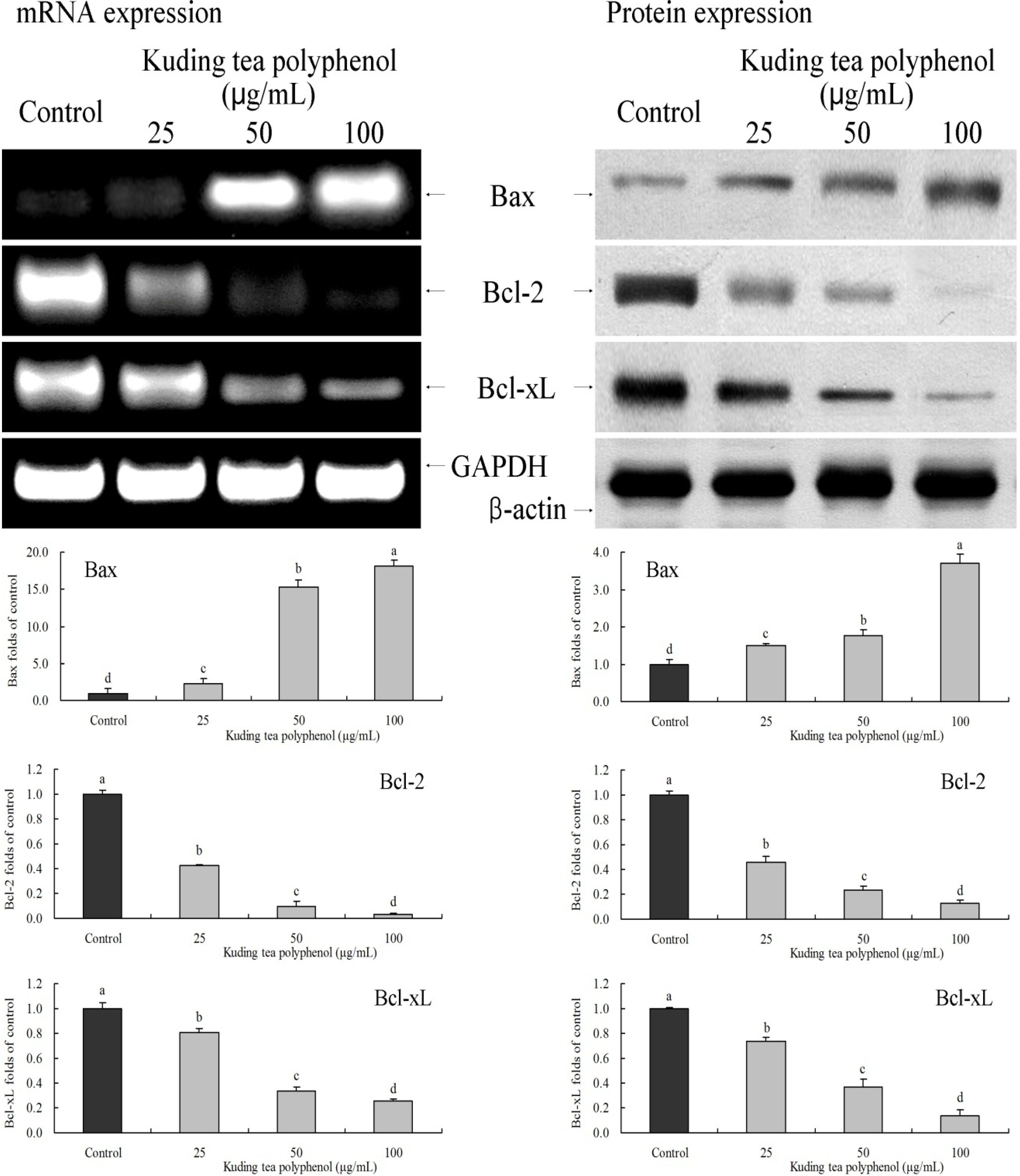
3.4. Gene Expression of the Apoptosis-Related Bcl-2 Family
3.5. Gene Expression of the Apoptosis-Related HIAP-1 and HIAP-2
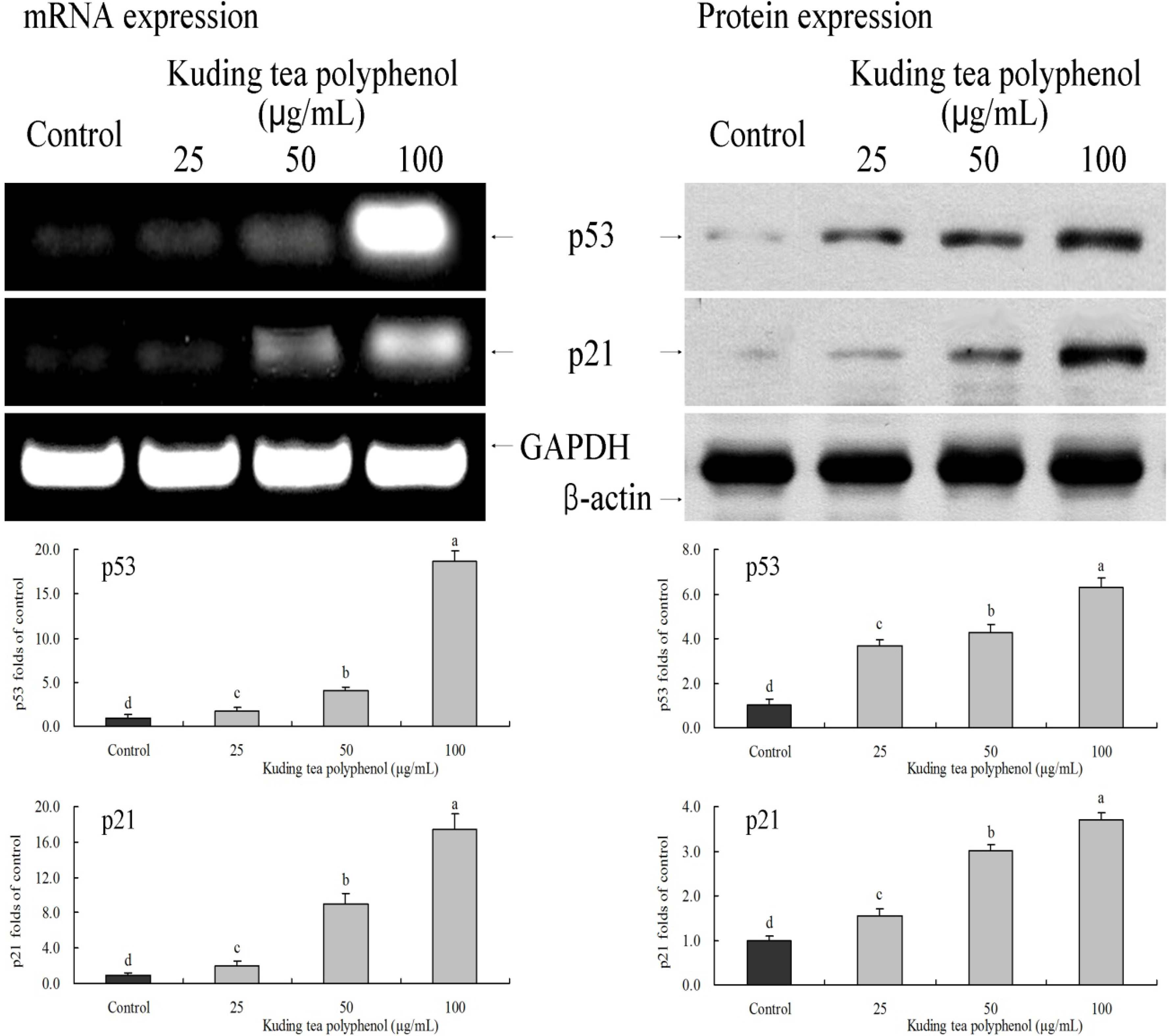
3.6. Gene Expression of the Apoptosis-Related p53 and p21



3.7. Gene Expression of the Apoptosis-Related E2F1 and p73

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhu, F.; Cai, Y.Z.; Sun, M.; Ke, J.; Lu, D.; Corke, H. Comparison of major phenolic constituents and in vitro antioxidant activity of diverse Kudingcha genotypes from Ilex Kudingcha, Ilex cornuta, and Ligustrum robustum. J. Agric. Food Chem. 2009, 57, 6082–6089. [Google Scholar] [CrossRef]
- Ding, B.; Wang, D.L.; Yin, J.Q.; Ma, W.; Zhou, G.Y.; Long, W.N. Effects of manganese on chemical qualities in Kuding tea. J. Jiangsu Forest. Sci. Technol. 2009, 36, 19–21. [Google Scholar]
- Song, J.L.; Qian, Y.; Li, G.J.; Zhao, X. Anti-inflammatory effects of Kudingcha methanol extract (Ilex Kudingcha C.J. Tseng) in dextran sulfate sodium-induced ulcerative colitis. Mol. Med. Rep. 2013, 8, 1256–1262. [Google Scholar]
- Gao, B.B.; She, G.M.; She, D.M. Chemical constituents and biological activities of plants from the genus ligustrum. Chem. Biodivers. 2013, 10, 96–128. [Google Scholar] [CrossRef]
- Chen, C.; Yu, R.; Owuor, E.D.; Kong, A.N. Activation of antioxidant-response element (ARE), mitogen-activated protein kinases (MAPKs) and caspases by major green tea polyphenol components during cell survival and death. Arch. Pharm. Res. 2000, 23, 605–612. [Google Scholar] [CrossRef]
- Mukhtar, H.; Ahmad, N. Tea polyphenols: Prevention of cancer and optimizing health. Am. J. Clin. Nutr. 2000, 71, 1698–1702. [Google Scholar]
- Ahmad, N.; Mukhtar, H. Green tea polyphenols and cancer: Biologic mechanisms and practical implications. Nutr. Rev. 1999, 57, 78–83. [Google Scholar] [CrossRef]
- Liu, L.X.; Tanguy, L.; Liang, X.F.; Sun, Y.; Ye, H.; Zeng, X.X. Dtermination of the total content of polyphenols in Kudingcha by Folin-Ciocalteu colorimetry. J. Tea Sci. 2008, 28, 101–106. [Google Scholar]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Pan, M.H.; Ghai, G.; Ho, C.T. Food bioactives, apoptosis, and cancer. Mol. Nutr. Food Res. 2008, 52, 43–52. [Google Scholar] [CrossRef]
- Fan, J.; Wu, Z.; Zhao, T.; Sun, Y.; Ye, H.; Xu, R.; Zeng, X. Characterization, antioxidant and hepatoprotective activities of polysaccharides from Ilex latifolia Thunb. Carbohydr. Polym. 2014, 101, 990–997. [Google Scholar] [CrossRef]
- Seo, H.R.; Kim, J.Y.; Kim, J.H.; Park, K.Y. Identification of Bacillus cereus in a chungkukjang that showed high anticancer effects against AGS human gastric adenocarcinoma cells. J. Med. Food 2009, 12, 1274–1280. [Google Scholar] [CrossRef]
- Zhao, X.; Ju, J.; Kim, H.M.; Park, K.Y. Antimutagenic activity and in vitro anticancer effects of bamboo salt on HepG2 human hepatoma cells. J. Environ. Pathol. Toxicol. Oncol. 2013, 32, 9–20. [Google Scholar] [CrossRef]
- Zhao, X.; Kim, S.Y.; Park, K.Y. Bamboo salt has in vitro anticancer activity in HCT-116 cells and exerts anti-metastatic effects in vivo. J. Med. Food 2013, 16, 9–19. [Google Scholar]
- Jiang, X.; Zhao, X.; Luo, H.; Zhu, K. Therapeutic effect of polysaccharide of large yellow croaker swim bladder on lupus nephritis of mice. Nutrients 2014, 6, 1223–1235. [Google Scholar] [CrossRef]
- Majors, B.S.; Betenbaugh, M.J.; Chiang, G.G. Links between metabolism and apoptosis in mammalian cells: Applications for anti-apoptosis engineering. Metab. Eng. 2007, 9, 317–326. [Google Scholar] [CrossRef]
- Jin, K.; Mao, X.O.; Batteur, S.P.; McEachron, E.; Leahy, A.; Greenberg, D.A. Caspase-3 and the regulation of hypoxic neuronal death by vascular endothelial growth factor. Neuroscience 2001, 108, 351–358. [Google Scholar] [CrossRef]
- Kajita, M.; Katayama, H.; Murata, T.; Kai, C.; Hori, M.; Ozaki, H. Canine distemper virus induces apoptosis through caspase-3 and -8 activation in vero cells. J. Vet. Med. B Infect. Dis. Vet. Public Health 2006, 53, 273–277. [Google Scholar] [CrossRef]
- Lawen, A. Apoptosis—An introduction. BioEssays 2003, 25, 888–896. [Google Scholar] [CrossRef]
- Wajant, H. The Fas signaling pathway: More than a paradigm. Science 2001, 31, 1635–1636. [Google Scholar]
- Krammer, P.H. CD95’s deadly mission in the immune system. Nature 2000, 407, 789–795. [Google Scholar] [CrossRef]
- Nakajima-Shimada, J.; Zou, C.; Takagi, M.; Umeda, M.; Nara, T.; Aoki, T. Inhibition of Fas-mediated apoptosis by Trypanosoma cruzi infection. Biochim. Biophys. Acta 2000, 1475, 175–183. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. The Bcl-2 apoptotic switch in cancer development and therapy. Oncogene 2007, 26, 1324–1337. [Google Scholar] [CrossRef]
- Brunelle, J.K.; Letai, A. Control of mitochondrial apoptosis by the Bcl-2 family. J. Cell Sci. 2009, 122, 437–441. [Google Scholar] [CrossRef]
- Asselin, E.; Mills, G.B.; Tsang, B.K. XIAP regulates Akt activity and caspase-3-dependent cleavage during cisplatin-induced apoptosis in human ovarian epithelial cancer cells. Cancer Res. 2001, 61, 1862–1868. [Google Scholar]
- Bilim, V.; Kasahara, T.; Hara, N.; Takahashi, K.; Tomita, Y. Role of XIAP in the malignant phenotype of transitional cell cancer (TCC) and therapeutic activity of XIAP antisense oligonucleotides against multidrug-resistant TCC in vitro. Int. J. Cancer 2003, 103, 29–37. [Google Scholar] [CrossRef]
- Sasaki, H.; Sheng, Y.; Kotsuji, F.; Tsang, B.K. Down-regulation of X-linked inhibitor of apoptosis protein induces apoptosis in chemoresistant human ovarian cancer cells. Cancer Res. 2000, 60, 5659–5666. [Google Scholar]
- Soussi, T. Focus on the p53 gene and cancer: Advances in TP53 mutation research. Hum. Mutat. 2003, 21, 173–175. [Google Scholar] [CrossRef]
- Chipuk, J.E.; Bouchier-Hayes, L.; Kuwana, T.; Newmeyer, D.D.; Green, D.R. PUMA couples the nuclear and cytoplasmic proapoptotic function of p53. Science 2005, 309, 1732–1735. [Google Scholar] [CrossRef]
- Wu, G.S.; Ding, Z.H. Caspase 9 is required for p53-dependent apoptosis and chemosensitivity in a human ovarian cancer cell line. Oncogene 2002, 21, 1–8. [Google Scholar] [CrossRef]
- Campomenosi, P.; Monti, P.; Aprile, A.; Abbondandolo, A.; Frebourg, T.; Gold, B.; Crook, T.; Inga, A.; Resnick, M.A.; Iggo, R.; Fronza, G. p53 mutants can often transactivate promoters containing a p21 but not Bax or PIG3 responsive elements. Oncogene 2001, 20, 3573–3579. [Google Scholar] [CrossRef]
- Rogoff, H.A.; Pickering, M.T.; Debatis, M.E.; Jones, S.; Kowalik, T.F. E2F1 induces phosphorylation of p53 that is coincident with p53 accumulation and apoptosis. Mol. Cell. Biol. 2002, 22, 5308–5318. [Google Scholar] [CrossRef]
- Irwin, M.; Marin, M.C.; Phillips, A.C.; Seelan, R.S.; Smith, D.I.; Liu, W.; Flores, E.R.; Tsai, K.Y.; Jacks, T.; Vousden, K.H.; Kaelin, W.G., Jr. Role for the p53 homologue p73 in E2F-1-induced apoptosis. Nature 2000, 407, 645–648. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhao, X.; Pang, L.; Li, J.; Song, J.-L.; Qiu, L.-H. Apoptosis Inducing Effects of Kuding Tea Polyphenols in Human Buccal Squamous Cell Carcinoma Cell Line BcaCD885. Nutrients 2014, 6, 3084-3100. https://doi.org/10.3390/nu6083084
Zhao X, Pang L, Li J, Song J-L, Qiu L-H. Apoptosis Inducing Effects of Kuding Tea Polyphenols in Human Buccal Squamous Cell Carcinoma Cell Line BcaCD885. Nutrients. 2014; 6(8):3084-3100. https://doi.org/10.3390/nu6083084
Chicago/Turabian StyleZhao, Xin, Liang Pang, Jing Li, Jia-Le Song, and Li-Hua Qiu. 2014. "Apoptosis Inducing Effects of Kuding Tea Polyphenols in Human Buccal Squamous Cell Carcinoma Cell Line BcaCD885" Nutrients 6, no. 8: 3084-3100. https://doi.org/10.3390/nu6083084