A Significant Inhibitory Effect on Advanced Glycation End Product Formation by Catechin as the Major Metabolite of Lotus Seedpod Oligomeric Procyanidins

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Materials
2.3. Preparation of LSOPC
2.4. Butanol-HCl Assay
2.5. Experimental Animals and Diets
2.6. Analysis of B-type Procyanidins and Their Metabolites in Urine
2.7. Inhibition of AGE Formation
2.8. Methylglyoxal Scavenging
2.9. 1,1-Diphenyl-2-picrylhydrazyl (DPPH) Radical Scavenging Activity of LSOPC and Its Metabolites
2.10. Total Antioxidant Capability (T-AOC) of LSOPC and Its Metabolites
2.11. Statistical Analyses and Graph Drawing
3. Results and Discussion
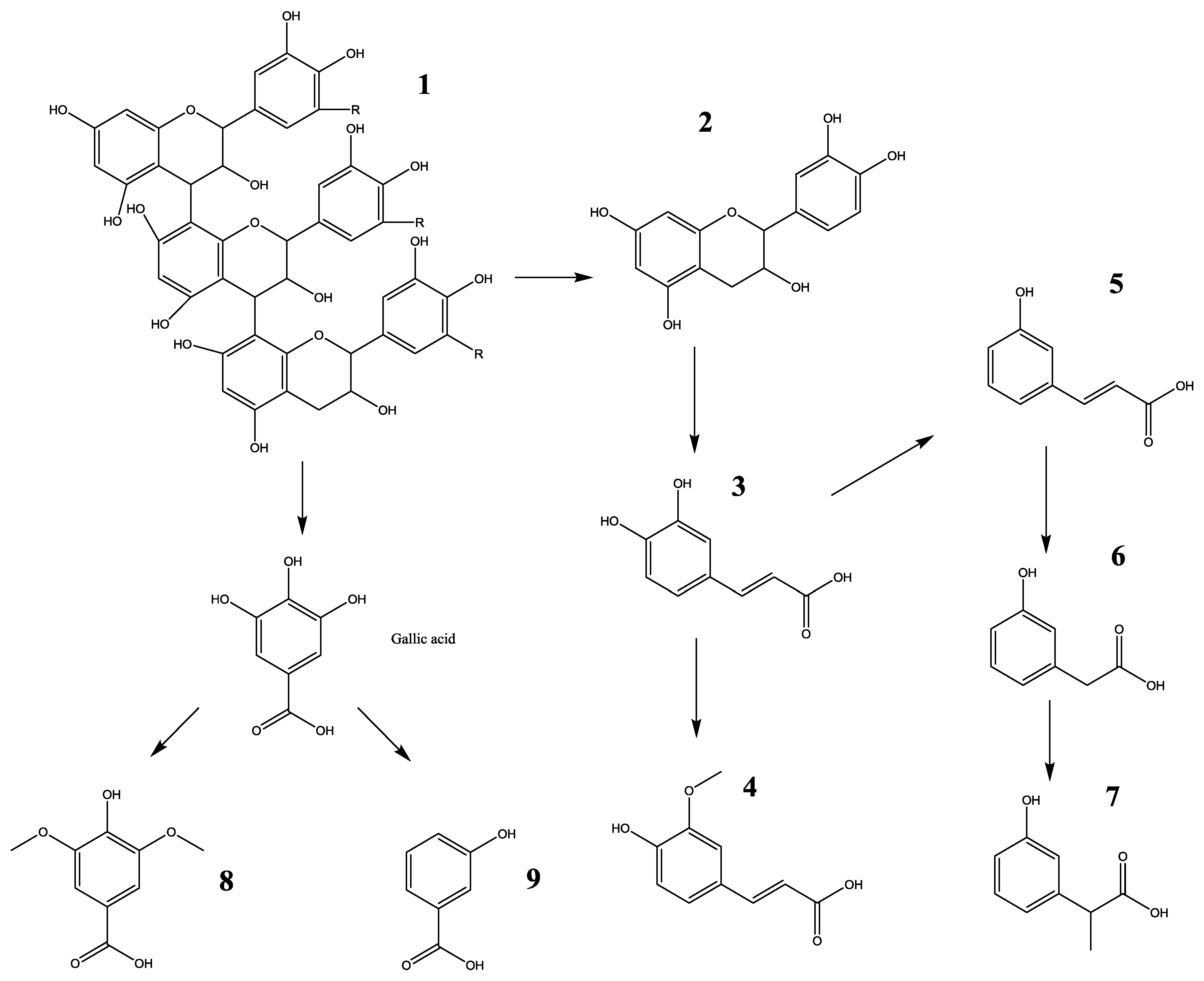
3.1. Analysis of Procyanidin Content in the Oligomeric Procyanidins of Lotus Seedpod (LSOPC)
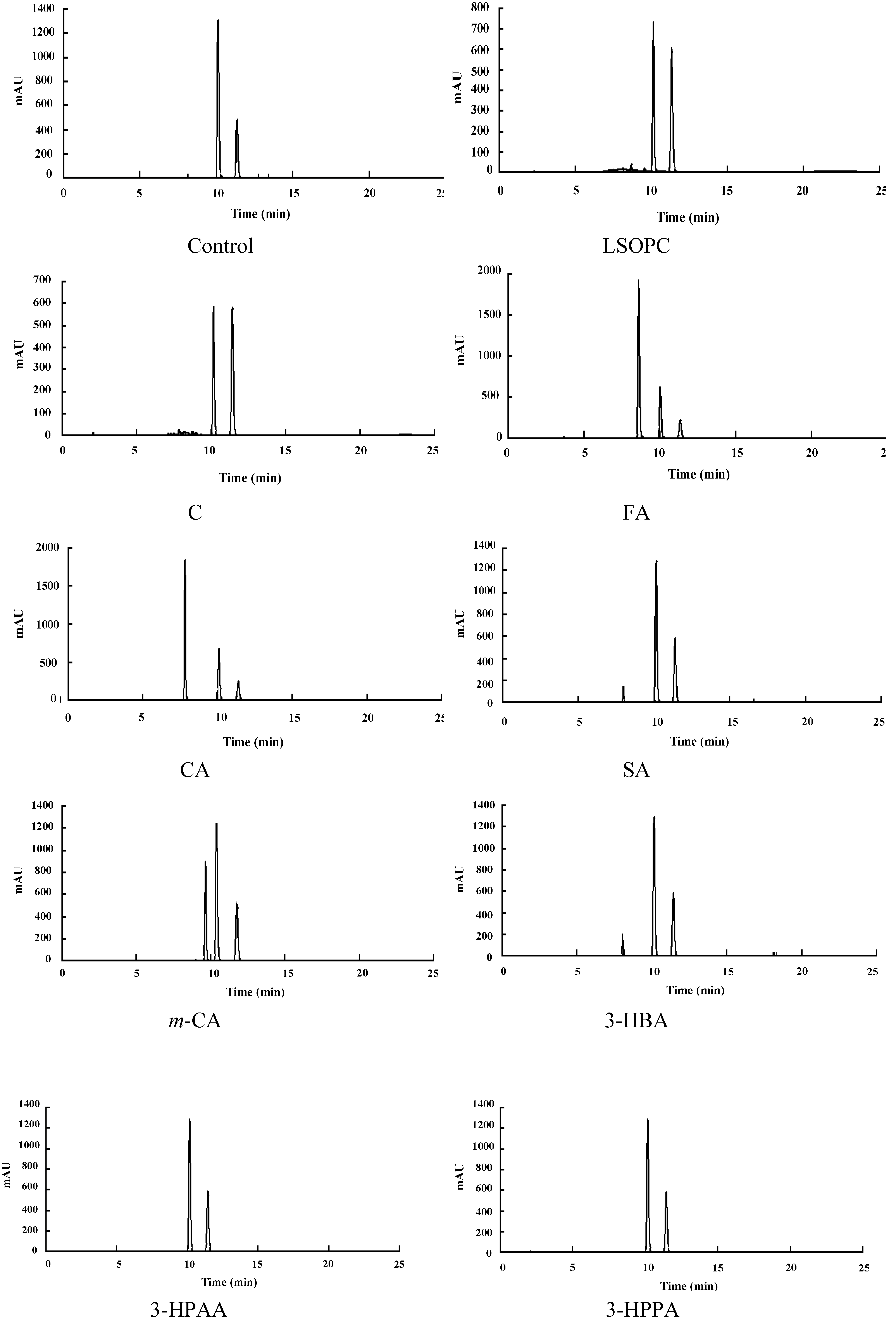
3.2. Determination of Metabolites in Rat Urine

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds (nmol) | Retention time (min) | Parent ion (m/z) | Product ion (m/z) | Urinary excretion within 24 h post-administration | |
|---|---|---|---|---|---|
| Control | 300 mg/kg body weight | ||||
| (+)-Catechin | 12.7 | 289.5 | 244.8 (CO2 loss), 124.8 (HRF, 2 H2O loss) | ND c | 202.76 ± 20.33 * |
| Caffeic acid | 14.8 | 179.3 | 134.7 (CO2 loss) | ND c | 22.46 ± 2.01 * |
| Syringic acid | 15.2 | 197.2 | 152.9 (CO2 loss) | ND c | 113.64 ± 12.14 * |
| 3-Hydroxybenzoic acid | 15.7 | 136.9 | 92.9 (CO2 loss) | ND c | 695.65 ± 59.47 * |
| 3-Hydroxyphenylacetic acid | 16.1 | 151.3 | 106.9 (CO2 loss) | 204.7 | 1046.05 ± 99.84 * |
| 3-Hydroxyphenylpropionic acid | 18.0 | 164.8 | 120.8 (CO2 loss) | ND c | 903.61 ± 89.05 * |
| Ferulic acid | 20.5 | 192.8 | 148.8 (CO2 loss) | 17.05 | 711.34 ± 70.86 * |
| m-Coumaric acid | 22.2 | 163.1 | 118.8 (CO2 loss) | 22.41 | 182.93 ± 16.58 * |


3.3. Inhibition of AGE Formation
| Compounds | IC50 (mg/ mL) | MGO Scavenging |
|---|---|---|
| LSOPC | 0.035 ± 0.004 | 81.24% ± 1.15% |
| (+)-Catechin | 0.049 ± 0.019 | 78.25% ± 2.99% |
| Ferulic acid | 0.741 ± 0.013 | 12.65% ± 0.53% |
| Caffeic acid | 0.683 ± 0.012 | 14.80% ± 0.29% |
| Syringic acid | 0.720 ± 0.017 | 20.61% ± 0.01% |
| m-Coumaric acid | 4.150 ± 0.012 | 16.69% ± 0.37% |
| 3-Hydroxybenzoic acid | 2.318 ± 0.030 | 19.30% ± 0.13% |
| 3-Hydroxyphenylacetic acid | 1.126 ± 0.023 | 20.16% ± 0.25% |
| 3-Hydroxyphenylpropionic acid | 1.899 ± 0.014 | 20.72% ± 0.42% |
3.4. Methylglyoxal Scavenging

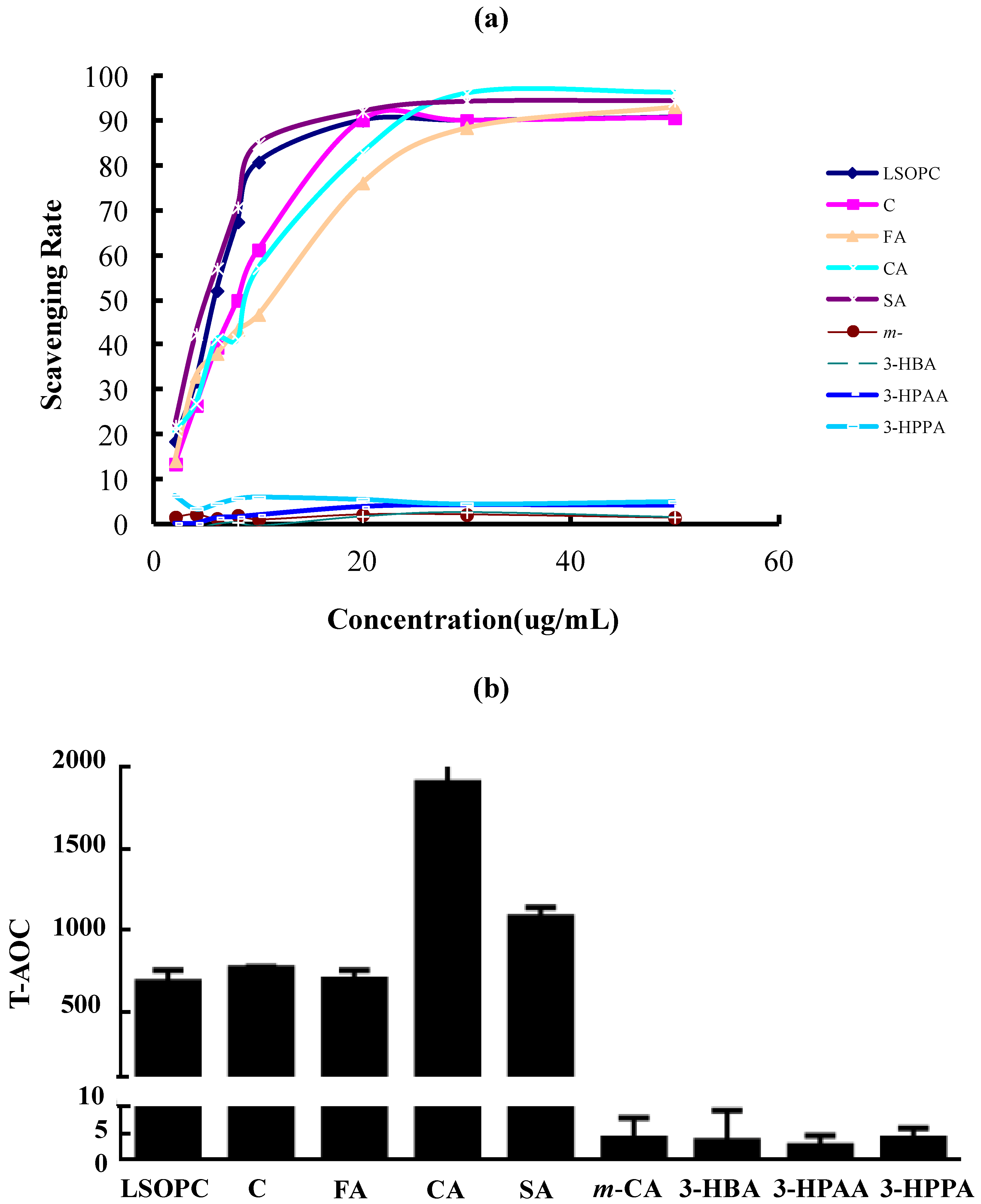
3.5. (DPPH) Radical Scavenging Activity and T-AOC of LSOPC and Its Metabolites

4. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Baynes, J.W.; Monnier, V.M. The Maillard reaction in aging diabetes and nutrition. Clin. Gerontol. 1991, 31, 420–422. [Google Scholar]
- Kanska, U.; Boratynski, J. Thermal glycation of proteins by d-glucose and d-fructose. Arch. Immunol. Ther. 2002, 50, 61–66. [Google Scholar]
- Miller, A.G.; Hegge, S.; Uhlmann, A.; Gerrard, J.A. A continuous enzyme assay and characterisation of fructosyl amine oxidase enzymes (EC 1.5.3). Arch. Biochem. Biophys. 2005, 434, 60–66. [Google Scholar] [CrossRef]
- Wu, C.H.; Yen, G.C. Inhibitory effect of naturally occurring flavonoids on the formation of advanced glycation endproducts. J. Agric. Food Chem. 2005, 53, 3167–3173. [Google Scholar] [CrossRef]
- Lin, Z.Q. Study on Procyanidins and Its Biological, Pharmacological Activities from the Seedpod of Nelumbo nucifera Gaertn. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, June 2001. [Google Scholar]
- Wang, Z.H.; Sun, Z.D.; Xie, B.J. Stability and Thermal Degradation Kinetics of rocyanidins from Lotus Seed Pods. J. Food Sci. 2011, 32, 77–82. [Google Scholar]
- Gong, Y.S.; Tang, Y.; Xiao, J.S.; Xie, B.J. Effects of procyanidins extracted from lotus seedpod on learning and memory impairment in mice. Acta Nutrimenta Sinia 2006, 28, 318–321. [Google Scholar]
- Wu, Q.; Chen, H.Y.; Lv, Z.J.; Li, S.Y.; Hu, B.; Guan, Y.F.; Xie, B.J.; Sun, Z.D. Oligomeric procyanidins of lotus seedpod inhibits the formation of advanced glycation end-products by scavenging reactive carbonyls. Food Chem. 2013, 138, 1493–1502. [Google Scholar]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar]
- Rios, L.Y.; Gonthier, M.P.; Rémésy, C.; Mila, I.; Lapierre, C.; Lazarus, S.A.; Williamson, G.; Scalbert, A. Chocolate intake increases urinary excretion of polyphenol-derived phenolic acids in healthy human subjects. Am. J. Clin. Nutr. 2003, 77, 912–918. [Google Scholar]
- Déprez, S.; Brezillon, C.; Rabot, S.; Philippe, C.; Mila, I.; Lapierre, C.; Scalbert, A. Polymeric proanthocyanidins are catabolized by human colonic microflora into low-molecular-weight phenolic acids. J. Nutr. 2000, 130, 2733–2738. [Google Scholar]
- Nagasawa, T.; Tabata, N.; Ito, Y.; Nishizawa, N.; Aiba, Y.; Kitts, D.D. Inhibition of glycation reaction in tissue protein incubations by water soluble rutin derivative. Mol. Cell. Biochem. 2003, 249, 3–10. [Google Scholar] [CrossRef]
- Lean, M.E.; Noroozi, M.; Kelly, I.; Burns, J.; Talwar, D.; Sattar, N. Dietary flavonols protect diabetic human lymphocytes against oxidative damage to DNA. Diabetes 1999, 48, 176–181. [Google Scholar] [CrossRef]
- Xiao, J.S.; Xie, B.J.; Gao, Y.P.; Wu, H.; Sun, Z.D.; Xiao, D. Characterization of oligomeric procyanidins and identification of quercetin glucuronide from lotus (Nelumbo nucifera Gaertn.) seedpod. J. Agric. Food Chem. 2012, 60, 2825–2829. [Google Scholar]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry 1985, 25, 223–230. [Google Scholar] [CrossRef]
- Gonthier, M.P.; Cheynier, V.; Donovan, J.L.; Manach, C.; Morand, C.; Mila, I.; Lapierre, C.; Rémésy, C.; Scalbert, A. Microbial aromatic acid metabolites formed in the gut account for a major fraction of the polyphenols excreted in urine of rats fed red wine polyphenols. J. Nutr. 2003, 133, 461–467. [Google Scholar]
- Brownlee, M.; Vlassara, H.; Kooney, A.; Ulrich, P.; Cerami, A. Aminoguanidine prevents diabetes-induced arterial wall protein cross-linking. Science 1986, 232, 1629–1632. [Google Scholar]
- Peng, X.; Zheng, Z.; Cheng, K.W.; Shan, F.; Ren, G.; Chen, F.; Wang, M. Inhibitory effect of mung bean extract and its constituents vitexin and isovitexin on the formation of advanced glycation endproducts. Food Chem. 2008, 106, 475–481. [Google Scholar] [CrossRef]
- Yang, B.; Zhou, M.M.; Shi, J.; Yang, N.; Jiang, Y.M. Effect of ultrasonic treatment on the recovery and DPPH radical scavenging activity of polysaccharides from longan fruit pericarp. Food Chem. 2008, 106, 685–690. [Google Scholar] [CrossRef]
- Gonthier, M.P.; Donovan, J.L.; Texier, O.; Felgines, C.; Remesy, C.; Scalbert, A. Metabolism of dietary procyanidins in rats. Free Radic. Biol. Med. 2003, 35, 837–844. [Google Scholar] [CrossRef]
- Natella, F.; Nardini, M.; Felice, D.M.; Scaccini, C. Benzoic and cinnamic acid derivatives as antioxidants: Structure-activity relation. J. Agric. Food Chem. 1999, 47, 1453–1459. [Google Scholar] [CrossRef]
- Rice, E.C.; Spencer, J.; Schroeter, H.; Rechner, A.R. Bioavailability of flavonoids and potential bioactive forms in vivo. Drug Metab. Drug Interact. 2011, 17, 291–310. [Google Scholar]
- Edelstein, D.; Brownlee, M. Mechanistic studies of advanced glycosylation end product inhibition by aminoguanidine. Diabetes 1992, 41, 26–29. [Google Scholar] [CrossRef]
- Peterson, D.G.; Totlani, V.M. Reactivity of epicatechin in aqueous glycine and glucose Maillard reaction models: Quenching of C2, C3, and C4 sugar fragments. J. Agric. Food Chem. 2005, 53, 4130–4135. [Google Scholar] [CrossRef]
- Baynes, J.W. Role of oxidative stress in development of complications in diabetes. Diabetes 1991, 40, 405–412. [Google Scholar] [CrossRef]
- Brownlee, M. Advanced protein glycosylation in diabetes and aging. Annu. Rev. Med. 1995, 46, 223–234. [Google Scholar] [CrossRef]
- Manske, C.L. Coronary artery disease in diabetic patients with nephropathy. Am. J. Hypertens 1993, 6, 367S–374S. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, Q.; Li, S.; Li, X.; Fu, X.; Sui, Y.; Guo, T.; Xie, B.; Sun, Z. A Significant Inhibitory Effect on Advanced Glycation End Product Formation by Catechin as the Major Metabolite of Lotus Seedpod Oligomeric Procyanidins. Nutrients 2014, 6, 3230-3244. https://doi.org/10.3390/nu6083230
Wu Q, Li S, Li X, Fu X, Sui Y, Guo T, Xie B, Sun Z. A Significant Inhibitory Effect on Advanced Glycation End Product Formation by Catechin as the Major Metabolite of Lotus Seedpod Oligomeric Procyanidins. Nutrients. 2014; 6(8):3230-3244. https://doi.org/10.3390/nu6083230
Chicago/Turabian StyleWu, Qian, Shuyi Li, Xiaopeng Li, Xiaoyan Fu, Yong Sui, Tingting Guo, Bijun Xie, and Zhida Sun. 2014. "A Significant Inhibitory Effect on Advanced Glycation End Product Formation by Catechin as the Major Metabolite of Lotus Seedpod Oligomeric Procyanidins" Nutrients 6, no. 8: 3230-3244. https://doi.org/10.3390/nu6083230