Long-Term Vitamin D3 Supplementation Does Not Prevent Colonic Inflammation or Modulate Bone Health in IL-10 Knockout Mice at Young Adulthood


Abstract
:1. Introduction
2. Experimental Section
2.1. Animals and Diet
2.2. Litter Characteristics and Body Weight

{kind=link}
{kind=link}
| Ingredient | Unit | Value |
|---|---|---|
| Macronutrients | ||
| Crude Protein | % | 17.9 |
| Crude Fat | % | 7.0 |
| Crude Fibre | % | 4.8 |
| Moisture | % | 7.0 |
| Ash | % | 4.2 |
| Amino Acids | ||
| Arginine | % | 0.70 |
| Lysine | % | 1.48 |
| Methionine | % | 0.56 |
| Cystine | % | 0.30 |
| Tryptophan | % | 0.21 |
| Histidine | % | 0.51 |
| Leucine | % | 1.76 |
| Isoleucine | % | 1.14 |
| Phenylalanine | % | 0.96 |
| Tyrosine | % | 0.98 |
| Threonine | % | 0.76 |
| Valine | % | 1.00 |
| Minerals | ||
| Calcium | mg/kg | 5000 |
| Phosphorus | mg/kg | 1561 |
| Potassium | mg/kg | 3600 |
| Sodium | mg/kg | 1019 |
| Magnesium | mg/kg | 507 |
| Iron | mg/kg | 35 |
| Zinc | mg/kg | 30 |
| Manganese | mg/kg | 10 |
| Copper | mg/kg | 6.0 |
| Iodine | mg/kg | 0.2 |
| Vitamins | ||
| Vitamin A | IU/g | 4.00 |
| Vitamin D3 | IU/g | 0.025 † or 5.00 †† |
| Alpha-Tocopherol | IU/g | 75.00 |
| Thiamine | mg | 5.0 |
| Riboflavin | mg | 6.0 |
| Niacin | mg | 30 |
| Pantothenic Acid | mg | 15.0 |
| Pyridoxine | mg | 6.0 |
| Folic Acid | mg | 2.0 |
| Biotin | mcg | 200 |
| Vitamin B12 | mcg | 25.0 |
| Vitamin K | mcg | 750 |
| Gross Energy | kcal/g | 3.80 |
2.3. Serum 25(OH)D
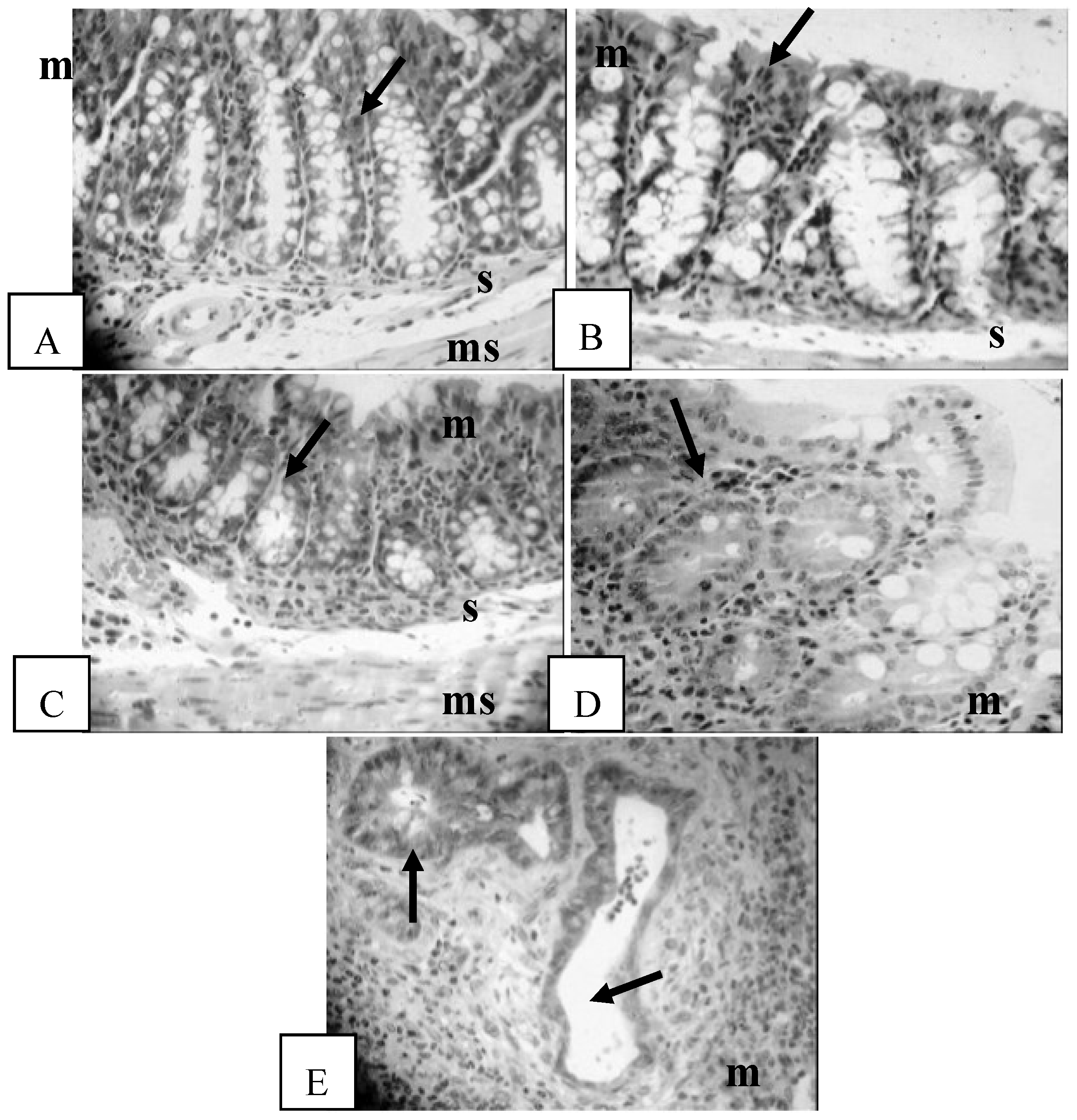
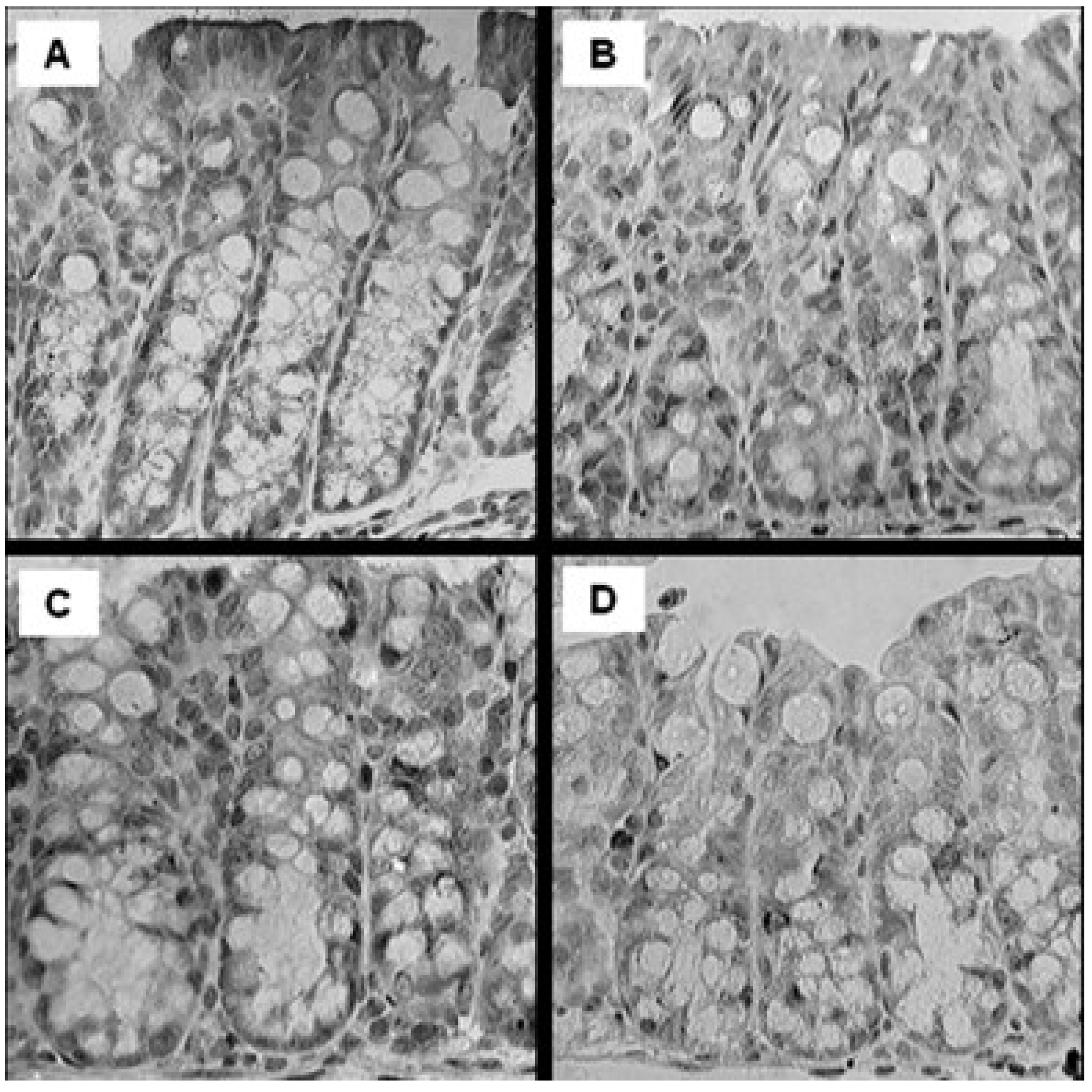
2.4. Histological Assessment
2.5. RNA Extraction
2.6. Microarray Analysis
2.7. Colonic VDR Expression
2.8. Cytokine and Bone Marker Analysis
2.9. Bone Morphometry, Bone Mineral Content (BMC), BMD and Biomechanical Strength Testing
2.10. Statistical Analyses
3. Results
3.1. Litter Characteristics and Body Weight
| Age | HH | HL | LH | LL | p Value |
|---|---|---|---|---|---|
| Females | |||||
| 1 month (g) | 12.1 ± 0.1 | 12.3 ± 1.1 | 12.4 ± 0.8 | 12.7 ± 0.6 | 0.96 |
| 3 months (g) | 20.0 ± 1.0 | 20.3 ± 1.1 | 19.8 ± 1.2 | 20.2 ± 1.3 | 0.95 |
| Males | |||||
| 1 month (g) | 13.7 ± 0.8 | 12.9 ± 1.1 | 14.5 ± 0.8 | 14.1 ± 0.0 | 0.10 |
| 3 months (g) | 23.5 ± 0.7 | 23.7 ± 1.1 | 23.8 ± 1.8 | 23.5 ± 0.8 | 0.94 |
3.2. Serum 25(OH)D
3.3. Histological Assessment
| Outcomes | HH | HL | LH | LL | Mother’s Diet | Pup’s Diet | Interaction |
|---|---|---|---|---|---|---|---|
| Females | |||||||
| Colon severity score | 1.4 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.3 | 1.4 ± 0.3 | 0.57 | 0.56 | 0.07 |
| Serum 25(OH)D (nmol/L) | 99.1 ± 4.1 a | 22.7 ± 1.5 b | 93.6 ± 7.5 a | 20.6 ± 0.6 b | 0.39 | <0.001 | 0.70 |
| IL-1α (pg/mL) | 153.5 ± 47.9 | 77.8 ± 55.3 | 175.2 ± 47.9 | 125.9 ± 60.6 | 0.51 | 0.25 | 0.81 |
| IL-6 (pg/mL) | 7.6 ± 7.6 | 12.9 ± 9.8 | 17.8 ± 7.6 | 28.8 ± 7.6 | 0.13 | 0.33 | 0.73 |
| IL-17 (pg/mL) | 17.0 ± 4.8 | 8.9 ± 4.8 | 14.8 ± 5.4 | 14.2 ± 4.8 | 0.75 | 0.39 | 0.45 |
| TNF-α (pg/mL) | 10.7 ± 2.6 | 6.1 ± 2.6 | 12.8 ± 2.8 | 7.4 ± 2.6 | 0.52 | 0.07 | 0.88 |
| Males | |||||||
| Colon severity score | 1.5 ± 0.2 | 1.5 ± 0.2 | 1.4 ± 0.2 | 1.7 ± 0.2 | 0.93 | 0.26 | 0.84 |
| Serum 25(OH)D (nmol/L) | 93.8 ± 2.7 a | 26.0 ± 1.6 b | 101.6 ± 10.3 a | 23.0 ± 2.2 b | 0.66 | <0.001 | 0.34 |
| IL-1α (pg/mL) | 60.6 ± 13.9 | 62.9 ± 18.1 | 42.2 ± 18.1 | 42.2 ± 18.0 | 0.28 | 0.94 | 0.96 |
| IL-6 (pg/mL) | 15.6 ± 7.1 | 13.9 ± 7.6 | 30.7 ± 7.1 | 37.3 ± 8.7 | 0.01 | 0.75 | 0.60 |
| IL-17 (pg/mL) | 26.4 ± 4.9 | 21.9 ± 5.2 | 20.8 ± 4.9 | 25.3 ± 5.6 | 0.83 | 0.99 | 0.39 |
| TNF-α (pg/mL) | 12.3 ± 2.1 | 10.9 ± 2.3 | 10.2 ± 1.9 | 11.3 ± 2.3 | 0.68 | 0.94 | 0.57 |

3.4. Colonic Gene Expression
3.5. Colonic VDR Expression
3.6. Cytokine Analysis

3.7. Serum Bone Markers and Femur and Lumbar Vertebrae Outcomes
| Bone Outcomes | HH | HL | LH | LL | Mother’s Diet | Pup’s Diet | Interaction |
|---|---|---|---|---|---|---|---|
| Females | |||||||
| RANKL (pg/mL) | 115 ± 10 a,b | 87 ± 10 a | 115 ± 9 a,b | 139 ± 10 b | 0.009 | 0.85 | 0.01 |
| OPG (pg/mL) | 1795 ± 182 | 1829 ± 182 | 1501 ± 172 | 1681 ± 193 | 0.23 | 0.56 | 0.68 |
| OPG/RANKL ratio | 16 ± 2 | 25 ± 5 | 12 ± 2 | 13 ± 2 | 0.004 | 0.06 | 0.16 |
| Femur midpoint yield load (N) | 10.1 ± 0.2 | 8.7 ± 0.4 | 9.4 ± 0.3 | 9.1 ± 0.4 | 0.66 | 0.005 | 0.11 |
| Femur midpoint peak load (N) | 19.8 ± 0.6 | 18.4 ± 0.4 | 18.6 ± 0.6 | 18.2 ± 0.4 | 0.17 | 0.09 | 0.34 |
| Femur neck peak load (N) | 10.0 ± 0.4 | 9.6 ± 0.4 | 10.1 ± 0.4 | 9.8 ± 0.3 | 0.71 | 0.33 | 0.84 |
| LV2 peak load (N) | 45.4 ± 1.9 | 44.1 ± 1.5 | 47.3 ± 1.5 | 46.2 ± 1.4 | 0.21 | 0.45 | 0.95 |
| Whole femur BMC (mg) | 20.7 ± 0.74 | 19.8 ± 0.89 | 19.1 ± 0.73 | 19.1 ± 0.94 | 0.20 | 0.92 | 0.34 |
| Whole femur BMD (mg/mm2) | 6.49 ± 0.16 | 6.41 ± 0.19 | 6.23 ± 0.18 | 6.18 ± 0.18 | 0.15 | 0.67 | 0.92 |
| 1/3 prox. femur BMC (mg) | 7.92 ± 0.24 | 7.62 ± 0.33 | 7.27 ± 0.25 | 7.52 ± 0.32 | 0.20 | 0.92 | 0.34 |
| 1/3 prox. femur BMD (mg/mm2) | 6.95 ± 0.15 | 6.87 ± 0.18 | 6.72 ± 0.19 | 6.62 ± 0.16 | 0.17 | 0.62 | 0.96 |
| LV4 BMC (mg) | 6.20 ± 0.39 | 6.49 ± 0.21 | 5.95 ± 0.38 | 5.63 ± 0.35 | 0.10 | 0.96 | 0.36 |
| LV4 BMD (mg/mm2) | 5.48 ± 0.01 | 5.73 ± 0.16 | 5.25 ± 0.29 | 5.29 ± 0.26 | 0.26 | 0.62 | 0.72 |
| Males | |||||||
| RANKL (pg/mL) | 116 ± 10 | 119 ± 10 | 119 ± 10 | 109 ± 10 | 0.74 | 0.72 | 0.49 |
| OPG (pg/mL) | 2016 ± 214 | 1992 ± 226 | 2027 ± 240 | 2154 ± 226 | 0.70 | 0.82 | 0.74 |
| OPG/RANKL ratio | 19 ± 2 | 18 ± 4 | 20 ± 5 | 22 ± 4 | 0.52 | 0.89 | 0.77 |
| Femur midpoint yield load (N) | 10.5 ± 0.26 | 9.95 ± 0.28 | 1.50 ± 0.02 | 10.36 ± 0.36 | 0.22 | 0.11 | 0.91 |
| Femur midpoint peak load (N) | 21.84 ± 0.60 | 20.45 ± 0.50 | 10.83 ± 0.35 | 21.41 ± 0.76 | 0.91 | 0.43 | 0.18 |
| Femur neck peak load (N) | 9.82 ± 0.38 | 9.30 ± 0.43 | 10.82 ± 0.54 | 10.08 ± 0.58 | 0.07 | 0.20 | 0.82 |
| LV2 peak load (N) | 51.6 ± 1.3 | 47.4 ± 1.3 | 51.7 ± 1.5 | 49.9 ± 2.2 | 0.43 | 0.07 | 0.44 |
| Whole femur BMC (mg) | 20.4 ± 0.49 | 19.8 ± 1.02 | 21.8 ± 0.76 | 19.5 ± 0.66 | 0.56 | 0.07 | 0.22 |
| Whole femur BMD (mg/mm2) | 6.18 ± 0.12 | 6.11 ± 0.18 | 6.46 ± 0.17 | 5.96 ± 0.12 | 0.65 | 0.07 | 0.16 |
| 1/3 prox. femur BMC (mg) | 8.00 ± 0.22 | 7.71 ± 0.37 | 8.38 ± 0.31 | 7.64 ± 0.34 | 0.61 | 0.10 | 0.48 |
| 1/3 prox. femur BMD (mg/mm2) | 6.63 ± 0.13 | 6.53 ± 0.19 | 6.88 ± 0.18 | 6.47 ± 0.13 | 0.54 | 0.12 | 0.33 |
| LV4 BMC (mg) | 6.30 ± 0.27 | 5.60 ± 0.21 | 6.39 ± 0.39 | 5.98 ± 0.25 | 0.17 | 0.55 | 0.56 |
| LV4 BMD (mg/mm2) | 5.12 ± 0.24 | 5.04 ± 0.18 | 5.29 ± 0.28 | 4.91 ± 0.10 | 0.93 | 0.27 | 0.46 |
4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sartor, R.B. Mechanisms of disease: Pathogenesis of Crohn’s disease and ulcerative colitis. Nat. Clin. Pract. Gastroenterol. Hepatol. 2006, 3, 390–407. [Google Scholar] [CrossRef]
- Sanchez-Munoz, F.; Dominguez-Lopez, A.; Yamamoto-Furusho, J.K. Role of cytokines in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 4280–4288. [Google Scholar] [CrossRef]
- Kwan Tat, S.; Padrines, M.; Theoleyre, S.; Heymann, D.; Fortun, Y. IL-6, RANKL, TNF-alpha/IL-1: Interrelations in bone resorption pathophysiology. Cytokine Growth Factor Rev. 2004, 15, 49–60. [Google Scholar]
- Van Hogezand, R.A.; Hamdy, N.A. Skeletal morbidity in inflammatory bowel disease. Scand. J. Gastroenterol. Suppl. 2006, 41, 59–64. [Google Scholar] [CrossRef]
- Wong, S.C.; Catto-Smith, A.G.; Zacharin, M. Pathological fractures in paediatric patients with inflammatory bowel disease. Eur. J. Pediatr. 2014, 173, 141–151. [Google Scholar] [CrossRef]
- Madsen, K.L. Inflammatory bowel disease: Lessons from the IL-10 gene-deficient mouse. Clin. Investig. Med. 2001, 24, 250–257. [Google Scholar]
- Kuhn, R.; Lohler, J.; Rennick, D.; Rajewsky, K.; Muller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 1993, 75, 263–274. [Google Scholar] [CrossRef]
- Cohen, S.L.; Moore, A.M.; Ward, W.E. Interleukin-10 knockout mouse: A model for studying bone metabolism during intestinal inflammation. Inflamm. Bowel Dis. 2004, 10, 557–563. [Google Scholar] [CrossRef]
- Ardizzone, S.; Cassinotti, A.; Bevilacqua, M.; Clerici, M.; Porro, G.B. Vitamin D and inflammatory bowel disease. Vitam. Horm. 2011, 86, 367–377. [Google Scholar]
- Pappa, H.M.; Grand, R.J.; Gordon, C.M. Report on the vitamin D status of adult and pediatric patients with inflammatory bowel disease and its significance for bone health and disease. Inflamm. Bowel Dis. 2006, 12, 1162–1174. [Google Scholar] [CrossRef]
- Dusso, A.S.; Brown, A.J.; Slatopolsky, E. Vitamin D. Am. J. Physiol. Ren. Physiol. 2005, 289, F8–F28. [Google Scholar] [CrossRef]
- Wang, T.T.; Tavera-Mendoza, L.E.; Laperriere, D.; Libby, E.; MacLeod, N.B.; Nagai, Y.; Bourdeau, V.; Konstorum, A.; Lallemant, B.; Zhang, R. Large-scale in silico and microarray-based identification of direct 1,25-dihydroxyvitamin D3 target genes. Mol. Endocrinol. 2005, 19, 2685–2695. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, H.; Wu, H.; Li, H.; Liu, L.; Guo, J.; Li, C; Shih, D.Q.; Zhang, X. Protective role of 1,25(OH)2 vitamin D3 in the mucosal injury and epithelial barrier disruption in DSS-induced acute colitis in mice. BMC Gastroenterol. 2012, 12, 57. [Google Scholar]
- Ryz, N.R.; Patterson, S.J.; Zhang, Y.; Ma, C.; Huang, T.; Bhinder, G.; Wu, X.; Chan, J.; Glesby, A.; Sham, H.P. Active vitamin D (1,25-dihydroxyvitamin D3) increases host susceptibility to Citrobacter rodentium by suppressing mucosal Th17 responses. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1299–G1311. [Google Scholar] [CrossRef]
- Bergstrand, O.; Hellers, G. Breast-feeding during infancy in patients who later develop Crohn’s disease. Scand. J. Gastroenterol. 1983, 18, 903–906. [Google Scholar] [CrossRef]
- Angelucci, E.; Cocco, A.; Cesarini, M.; Crudeli, A.; Necozione, S.; Caprilli, R.; Latella, G. Monthly and seasonal birth patterns and the occurrence of Crohn’s disease. Am. J. Gastroenterol. 2009, 104, 1608–1609. [Google Scholar] [CrossRef]
- Lagishetty, V.; Misharin, A.V.; Liu, N.Q.; Lisse, T.S.; Chun, R.F.; Ouyang, Y.; McLachlan, S.M.; Adams, J.S.; Hewison, M. Vitamin D deficiency in mice impairs colonic antibacterial activity and predisposes to colitis. Endocrinology 2010, 151, 2423–2432. [Google Scholar] [CrossRef]
- Ooi, J.H.; Li, Y.; Rogers, C.J.; Cantorna, M.T. Vitamin D regulates the gut microbiome and protects mice from dextran sodium sulfate-induced colitis. J. Nutr. 2013, 143, 1679–1686. [Google Scholar] [CrossRef]
- Froicu, M.; Zhu, Y.; Cantorna, M.T. Vitamin D receptor is required to control gastrointestinal immunity in IL-10 knockout mice. Immunology 2006, 117, 310–318. [Google Scholar] [CrossRef]
- Cantorna, M.T.; Munsick, C.; Bemiss, C.; Mahon, B.D. 1,25-Dihydroxycholecalciferol prevents and ameliorates symptoms of experimental murine inflammatory bowel disease. J. Nutr. 2000, 130, 2648–2652. [Google Scholar]
- Canadian Council on Animal Care. Guide to the Care and Use of Experimental Animals; Ernest, D., Olfert, D.V.M., Brenda, M., Cross, D.V.M., McWilliam, A., Eds.; Canadian Council on Animal Care: Ottawa, ON, Canada, 1993; Volume 1, pp. 1–212. [Google Scholar]
- Fleet, J.C.; Gliniak, C.; Zhang, Z.; Xue, Y.; Smith, K.B.; McCreedy, R.; Adedokun, S.A. Serum metabolite profiles and target tissue gene expression define the effect of cholecalciferol intake on calcium metabolism in rats and mice. J. Nutr. 2008, 138, 1114–1120. [Google Scholar]
- Allred, D.C.; Harvey, J.M.; Berardo, M.; Clark, G.M. Prognostic and predictive factors in breast cancer by immunohistochemical analysis. Mod. Pathol. 1998, 11, 155–168. [Google Scholar]
- Cohen, S.L.; Moore, A.M.; Ward, W.E. Flaxseed oil and inflammation-associated bone abnormalities in interleukin-10 knockout mice. J. Nutr. Biochem. 2005, 16, 368–374. [Google Scholar] [CrossRef]
- R Project for Statistical Computing. Version 2.14.1. Available online: http://www.r-project.org/ (accessed on 17 September 2014).
- Gene Expression Omnibus. Available online: http://www.ncbi.nlm.nih.gov/geo/ (accessed on 15 September 2014).
- Kaludjerovic, J.; Ward, W.E. Neonatal exposure to daidzein, genistein, or the combination modulates bone development in female CD-1 mice. J. Nutr. 2009, 139, 467–473. [Google Scholar] [CrossRef]
- Lagunova, Z.; Porojnicu, A.C.; Vieth, R.; Lindberg, F.A.; Hexeberg, S.; Moan, J. Serum 25-hydroxyvitamin D is a predictor of serum 1,25-dihydroxyvitamin D in overweight and obese patients. J. Nutr. 2011, 141, 112–117. [Google Scholar] [CrossRef]
- Mathieu, C.; van Etten, E.; Gysemans, C.; Decallonne, B.; Kato, S.; Laureys, J.; Depovere, J.; Valckx, D.; Verstuyf, A.; Bouillon, R. In vitro and in vivo analysis of the immune system of vitamin D receptor knockout mice. J. Bone Miner. Res. 2001, 16, 2057–2065. [Google Scholar] [CrossRef]
- Reiff, C.; Delday, M.; Rucklidge, G.; Reid, M.; Duncan, G.; Wohlgemuth, S.; Hormannsperger, G.; Loh, G.; Blaut, M.; Collie-Duguid, E. Balancing inflammatory, lipid, and xenobiotic signaling pathways by VSL#3, a biotherapeutic agent, in the treatment of inflammatory bowel disease. Inflamm. Bowel Dis. 2009, 15, 1721–1736. [Google Scholar] [CrossRef]
- Dresner-Pollak, R.; Gelb, N.; Rachmilewitz, D.; Karmeli, F.; Weinreb, M. Interleukin 10-deficient mice develop osteopenia, decreased bone formation, and mechanical fragility of long bones. Gastroenterology 2004, 127, 792–801. [Google Scholar] [CrossRef]
- Reeves, P.G.; Rossow, K.L.; Lindlauf, J. Development and testing of the AIN-93 purified diets for rodents: Results on growth, kidney calcification and bone mineralization in rats and mice. J. Nutr. 1993, 123, 1923–1931. [Google Scholar]
- Larmonier, C.B.; McFadden, R.M.; Hill, F.M.; Schreiner, R.; Ramalingam, R.; Besselsen, D.G.; Ghishan, F.K.; Kiela, P.R. High vitamin D3 diet administered during active colitis negatively affects bone metabolism in an adoptive T cell transfer model. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G35–G46. [Google Scholar] [CrossRef]
- Spelling, P.; Bonfa, E.; Caparbo, V.F.; Pereira, R.M. Osteoprotegerin/RANKL system imbalance in active polyarticular-onset juvenile idiopathic arthritis: A bone damage biomarker? Scand. J. Rheumatol. 2008, 37, 439–444. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Glenn, A.J.; Fielding, K.A.; Chen, J.; Comelli, E.M.; Ward, W.E. Long-Term Vitamin D3 Supplementation Does Not Prevent Colonic Inflammation or Modulate Bone Health in IL-10 Knockout Mice at Young Adulthood. Nutrients 2014, 6, 3847-3862. https://doi.org/10.3390/nu6093847
Glenn AJ, Fielding KA, Chen J, Comelli EM, Ward WE. Long-Term Vitamin D3 Supplementation Does Not Prevent Colonic Inflammation or Modulate Bone Health in IL-10 Knockout Mice at Young Adulthood. Nutrients. 2014; 6(9):3847-3862. https://doi.org/10.3390/nu6093847
Chicago/Turabian StyleGlenn, Andrea J., Kristina A. Fielding, Jianmin Chen, Elena M. Comelli, and Wendy E. Ward. 2014. "Long-Term Vitamin D3 Supplementation Does Not Prevent Colonic Inflammation or Modulate Bone Health in IL-10 Knockout Mice at Young Adulthood" Nutrients 6, no. 9: 3847-3862. https://doi.org/10.3390/nu6093847