What Do Studies of Insect Polyphenisms Tell Us about Nutritionally-Triggered Epigenomic Changes and Their Consequences?

{kind=link}
Abstract
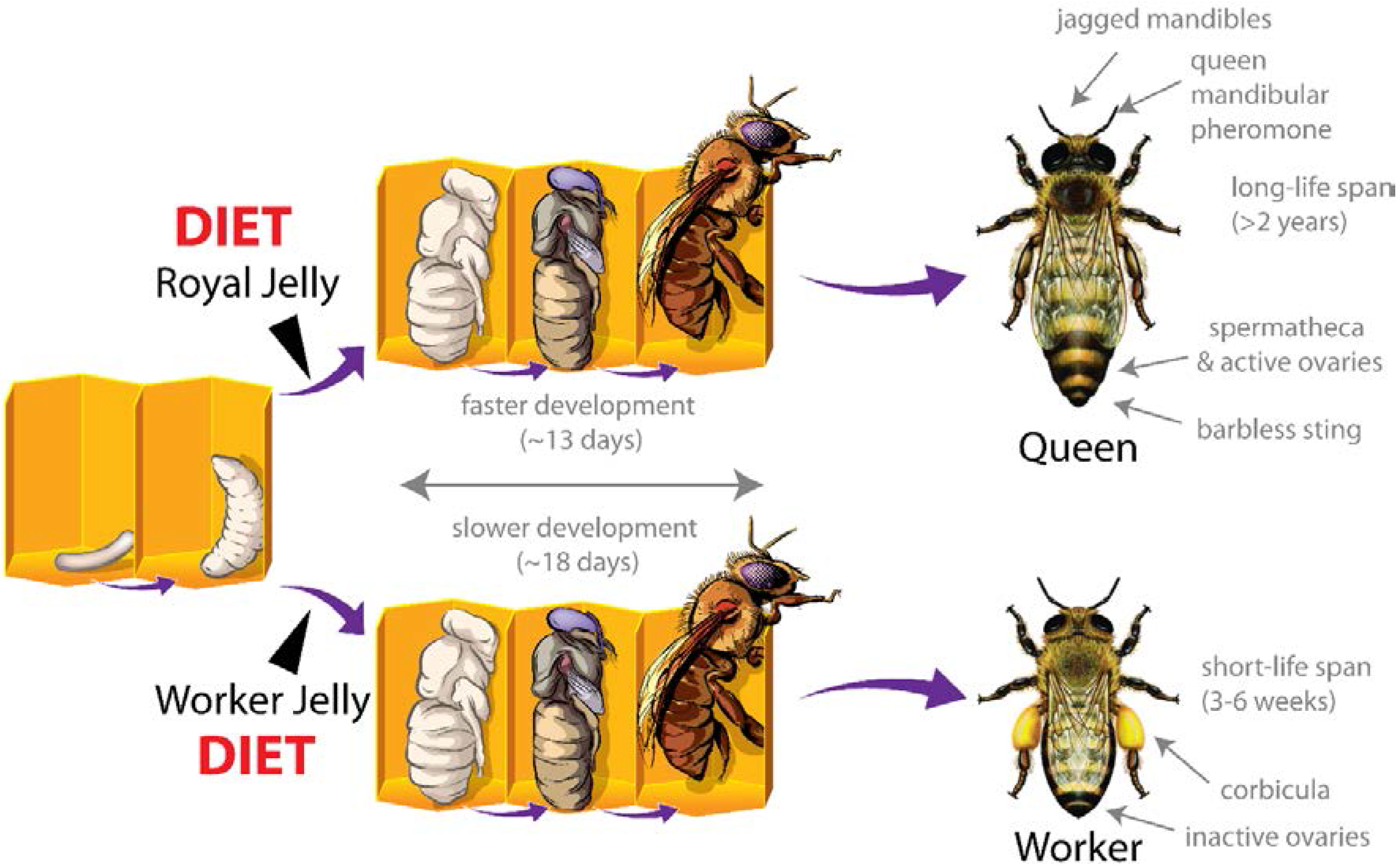
:1. Introduction

2. DNA Methylation and Nutritional Epigenetics
3. Histone Modifications and Insect Polyphenisms
4. Other Epigenetic Mechanisms and Insect Polyphenisms
4.1. Alternative Splicing
4.2. RNA-Editing
4.3. Gene Regulation by Non-Coding RNAs
4.4. Short ncRNA
4.5. miRNA
4.6. PIWI RNA
4.7. lnRNA (Long Noncodingrna)
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Glastad, K.M.; Hunt, B.G.; Yi, S.V.; Goodisman, M.A. DNA methylation in insects: On the brink of the epigenomic era. Insect Mol. Biol. 2011, 20, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Haydak, M.H. Honey bee nutrition. Annu Rev Entomol 1970, 15, 143–156. [Google Scholar] [CrossRef]
- Cameron, R.; Duncan, E.; Dearden, P. Biased gene expression in early honeybee larval development. BMC Genomics 2013, 14, 903. [Google Scholar] [CrossRef] [PubMed]
- Kucharski, R.; Maleszka, J.; Foret, S.; Maleszka, R. Nutritional control of reproductive status in honeybees via DNA methylation. Science 2008, 319, 1827–1830. [Google Scholar] [CrossRef] [PubMed]
- Duncan, E.J.; Gluckman, P.D.; Dearden, P.K. Epigenetics, plasticity, and evolution: How do we link epigenetic change to phenotype? J. Exp. Zool. Part B Mol. Dev. Evol. 2014, 322, 208–220. [Google Scholar] [CrossRef]
- Zemach, A.; McDaniel, I.E.; Silva, P.; Zilberman, D. Genome-wide evolutionary analysis of eukaryotic DNA methylation. Science 2010, 328, 916–919. [Google Scholar] [CrossRef] [PubMed]
- Lyko, F.; Foret, S.; Kucharski, R.; Wolf, S.; Falckenhayn, C.; Maleszka, R. The honey bee epigenomes: Differential methylation of brain DNA in queens and workers. PLoS Biol. 2010, 8, e1000506. [Google Scholar] [CrossRef] [PubMed]
- Foret, S.; Kucharski, R.; Pellegrini, M.; Feng, S.; Jacobsen, S.E.; Robinson, G.E.; Maleszka, R. DNA methylation dynamics, metabolic fluxes, gene splicing, and alternative phenotypes in honey bees. Proc. Natl. Acad. Sci. USA 2012, 109, 4968–4973. [Google Scholar] [CrossRef] [PubMed]
- Sati, S.; Tanwar, V.S.; Kumar, K.A.; Patowary, A.; Jain, V.; Ghosh, S.; Ahmad, S.; Singh, M.; Reddy, S.U.; Chandak, G.R.; et al. High resolution methylome map of rat indicates role of intragenic DNA methylation in identification of coding region. PLoS One 2012, 7, e31621. [Google Scholar] [CrossRef]
- Shukla, S.; Kavak, E.; Gregory, M.; Imashimizu, M.; Shutinoski, B.; Kashlev, M.; Oberdoerffer, P.; Sandberg, R.; Oberdoerffer, S. CTCF-promoted RNA polymerase II pausing links DNA methylation to splicing. Nature 2011, 479, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Maunakea, A.K.; Nagarajan, R.P.; Bilenky, M.; Ballinger, T.J.; D’Souza, C.; Fouse, S.D.; Johnson, B.E.; Hong, C.; Nielsen, C.; Zhao, Y.; et al. Conserved role of intragenic DNA methylation in regulating alternative promoters. Nature 2010, 466, 253–257. [Google Scholar] [CrossRef]
- Lorincz, M.C.; Dickerson, D.R.; Schmitt, M.; Groudine, M. Intragenic DNA methylation alters chromatin structure and elongation efficiency in mammalian cells. Nat. Struct. Mol. Biol. 2004, 11, 1068–1075. [Google Scholar] [CrossRef] [PubMed]
- Cedar, H.; Bergman, Y. Linking DNA methylation and histone modification: Patternspatterns and paradigms. Nature reviews. Genetics 2009, 10, 295–304. [Google Scholar] [PubMed]
- Hunt, B.G.; Glastad, K.M.; Yi, S.V.; Goodisman, M.A. Patterning and regulatory associations of DNA methylation are mirrored by histone modifications in insects. Genome Biol. Evol. 2013, 5, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Lambrot, R.; Xu, C.; Saint-Phar, S.; Chountalos, G.; Cohen, T.; Paquet, M.; Suderman, M.; Hallett, M.; Kimmins, S. Low paternal dietary folate alters the mouse sperm epigenome and is associated with negative pregnancy outcomes. Nat. Commun. 2013, 4, 2889. [Google Scholar] [CrossRef] [PubMed]
- Parle-McDermott, A.; Ozaki, M. The impact of nutrition on differential methylated regions of the genome. Adv. Nutr. 2011, 2, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Wolff, G.L.; Kodell, R.L.; Moore, S.R.; Cooney, C.A. Maternal epigenetics and methyl supplements affect agouti gene expression in avy/a mice. FASEB J. 1998, 12, 949–957. [Google Scholar] [PubMed]
- Kamakura, M. Royalactin induces queen differentiation in honeybees. Nature 2011, 473, 478–483. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, A.R.; Rouleau, J.; Szyf, M. Regulation of DNA methylation by the ras signaling pathway. J. Boil. Chem. 1995, 270, 11327–11337. [Google Scholar] [CrossRef]
- Samudio-Ruiz, S.L.; Hudson, L.G. Increased DNA methyltransferase activity and DNA methylation following epidermal growth factor Epidermal Growth Factor stimulation in ovarian cancer cells. Epigenetics 2012, 7, 216–224. [Google Scholar]
- Snell-Rood, E.C.; Troth, A.; Moczek, A.P. DNA methylation as a mechanism of nutritional plasticity: Limited support from horned beetles. J. Exp. Zool. Part B Mol. Dev. Evol. 2013, 320, 22–34. [Google Scholar] [CrossRef]
- Bonasio, R.; Li, Q.; Lian, J.; Mutti, N.S.; Jin, L.; Zhao, H.; Zhang, P.; Wen, P.; Xiang, H.; Ding, Y.; et al. Genome-wide and caste-specific DNA methylomes of the ants camponotus floridanus and harpegnathos saltator. Curr. Biol. 2012, 22, 1755–1764. [Google Scholar] [CrossRef]
- Wang, X.; Kang, L. Molecular mechanisms of phase change in locusts. Annual review of entomology. Annu. Rev. Entomol. 2014, 59, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Lim, U.; Song, M.A. Dietary and lifestyle factors of DNA methylation. Methods Mol. Biol. 2012, 863, 359–376. [Google Scholar] [PubMed]
- Lan, X.; Cretney, E.C.; Kropp, J.; Khateeb, K.; Berg, M.A.; Penagaricano, F.; Magness, R.; Radunz, A.E.; Khatib, H. Maternal diet during pregnancy induces gene expression Pregnancy Induces Gene Expression and DNA methylation changes Methylation Changes in fetal tissues Fetal Tissues in sheep. Front. Genet. 2013, 4, 49. [Google Scholar] [CrossRef] [PubMed]
- Carone, B.R.; Fauquier, L.; Habib, N.; Shea, J.M.; Hart, C.E.; Li, R.; Bock, C.; Li, C.; Gu, H.; Zamore, P.D.; et al. Paternally induced transgenerational environmental reprogramming of metabolic gene expression in mammals. Cell 2010, 143, 1084–1096. [Google Scholar] [CrossRef]
- Martinez, D.; Pentinat, T.; Ribo, S.; Daviaud, C.; Bloks, V.W.; Cebria, J.; Villalmanzo, N.; Kalko, S.G.; Ramon-Krauel, M.; Diaz, R.; et al. In utero undernutrition in male mice programs liver lipid metabolism in the second-generation offspring involving altered Lxra DNA methylation. Cell Metab. 2014, 19, 941–951. [Google Scholar] [CrossRef]
- Ng, S.F.; Lin, R.C.; Laybutt, D.R.; Barres, R.; Owens, J.A.; Morris, M.J. Chronic high-fat diet in fathers programs beta-cell dysfunction in female rat offspring. Nature 2010, 467, 963–966. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Yang, C.R.; Wei, Y.P.; Zhao, Z.A.; Hou, Y.; Schatten, H.; Sun, Q.Y. Paternally induced transgenerational inheritance of susceptibility to diabetes in mammals. Proc. Natl. Acad. Sci. USA 2014, 111, 1873–1878. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, M.; Smith, M.M. Functional consequences of histone modifications. Curr. Opin. Genet. Dev. 2003, 13, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Margueron, R.; Reinberg, D. The polycomb complex prc2PRC2 and its mark in life. Nature 2011, 469, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, C.; Prado, J.R.M.; Eun, S.H.; Fuller, M.T. Sequential changes at differentiation gene promoters as they become active in a stem cell lineage. Development 2011, 138, 2441–2450. [Google Scholar] [CrossRef] [PubMed]
- Iovino, N.; Ciabrelli, F.; Cavalli, G. Prc2 controls drosophila oocyte cell fate by repressing cell cycle genes. Devel. Cell 2013, 26, 431–439. [Google Scholar] [CrossRef]
- Feldmeyer, B.; Elsner, D.; Foitzik, S. Gene expression patterns associated with caste and reproductive status in ants: Worker-specific genes are more derived than queen-specific ones. Mol. Ecol. 2014, 23, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Bonasio, R.; Zhang, G.; Ye, C.; Mutti, N.S.; Fang, X.; Qin, N.; Donahue, G.; Yang, P.; Li, Q.; Li, C. Genomic comparison of the ants Camponotus floridanus and Harpegnathos saltator. Science 2010, 329, 1068–1071. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.; Zeng, Z.J.; Wang, Z.L.; Yan, W.Y.; Pan, Q.Z. Expression of SIR2, HDAC1 SIR2, HDAC1 and Ash2Ash2 in honey bee (Apis mellifera L.) queens Honey Bee (Apis Mellifera L.) Queens and workers. J. Apic. Sci. 2013, 57, 67–73. [Google Scholar]
- Simola, D.F.; Wissler, L.; Donahue, G.; Waterhouse, R.M.; Helmkampf, M.; Roux, J.; Nygaard, S.; Glastad, K.M.; Hagen, D.E.; Viljakainen, L.; et al. Social insect genomes exhibit dramatic evolution in gene composition and regulation while preserving regulatory features linked to sociality. Genome Res. 2013, 23, 1235–1247. [Google Scholar] [CrossRef]
- Spannhoff, A.; Kim, Y.K.; Raynal, N.J.M.; Gharibyan, V.; Su, M.B.; Zhou, Y.Y.; Li, J.; Castellano, S.; Sbardella, G.; Issa, J.P.J. Histone deacetylase inhibitor activity in royal jelly might facilitate caste switching in bees. EMBO Rep. 2011, 12, 238–243. [Google Scholar] [PubMed]
- Paoli, P.P.; Wakeling, L.A.; Wright, G.A.; Ford, D. The dietary proportion of essential amino acids and Sir2 influence lifespan in the honeybee. Age 2014, 36, 9649. [Google Scholar] [CrossRef] [PubMed]
- Flores, K.; Wolschin, F.; Corneveaux, J.J.; Allen, A.N.; Huentelman, M.J.; Amdam, G.V. Genome-wide association between DNA methylation and alternative splicing in an invertebrate. BMC Genomics 2012, 13, 480. [Google Scholar] [CrossRef] [PubMed]
- Li-Byarlay, H.; Li, Y.; Stroud, H.; Feng, S.; Newman, T.C.; Kaneda, M.; Hou, K.K.; Worley, K.C.; Elsik, C.G.; Wickline, S.A.; et al. RNA interference knockdown of DNA methyl-transferase 3 affects gene alternative splicing in the honey bee. Proc. Natl. Acad. Sci. USA 2013, 110, 12750–12755. [Google Scholar] [CrossRef] [PubMed]
- Gott, J.M.; Emeson, R.B. Functions and mechanisms of RNA editing. Annu. Rev. Genet. 2000, 34, 499–531. [Google Scholar] [CrossRef] [PubMed]
- Bass, B.L. RNA editing by adenosine deaminases that act on RNA. Annu. Rev. Biochem. 2002, 71, 817–846. [Google Scholar] [CrossRef] [PubMed]
- Laurencikiene, J.; Kallman, A.M.; Fong, N.; Bentley, D.L.; Ohman, M. RNA editing and alternative splicing: The importance of co-transcriptional coordination. EMBO Rep. 2006, 7, 303–307. [Google Scholar] [PubMed]
- Kawahara, Y.; Zinshteyn, B.; Sethupathy, P.; Iizasa, H.; Hatzigeorgiou, A.G.; Nishikura, K. Redirection of silencing targets by adenosine-to-inosine editing of mirnasmiRNAs. Science 2007, 315, 1137–1140. [Google Scholar] [CrossRef] [PubMed]
- Borchert, G.M.; Gilmore, B.L.; Spengler, R.M.; Xing, Y.; Lanier, W.; Bhattacharya, D.; Davidson, B.L. Adenosine deamination in human transcripts generates novel micrornamicroRNA binding sites. Hum. Mol. Genet. 2009, 18, 4801–4807. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, Z.; Lian, J.; Schiott, M.; Jin, L.; Zhang, P.; Zhang, Y.; Nygaard, S.; Peng, Z.; Zhou, Y.; et al. Caste-specific RNA editomes in the leaf-cutting ant Acromyrmex echinatior. Nat. Commun. 2014, 5, 4943. [Google Scholar] [CrossRef] [PubMed]
- Bonasio, R. Emerging topics in epigenetics: Antsants, brains, and noncoding RNAs. Ann. NY Acad. Sci. 2012, 126014–126023. [Google Scholar] [CrossRef]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, J.K.; Xia, J.; Zhou, X.; Thatcher, S.R.; Gu, X.; Ament, S.A.; Newman, T.C.; Green, P.J.; Zhang, W.; Robinson, G.E.; et al. Behavioral plasticity in honey bees is associated with differences in brain micrornamicro RNA transcriptome. Genes Brain Behav. 2012, 11, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Su, S.; Skogerboe, G.; Dai, S.; Li, W.; Li, Z.; Liu, F.; Ni, R.; Guo, Y.; Chen, S.; et al. Recipe for a busy bee: Micrornas Busy Bee: MicroRNAs in honey bee caste determination. PLoS One 2013, 8, e81661. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.Y.; Wu, X.B.; Huang, Z.Y.; Wang, Z.L.; Yan, W.Y.; Zeng, Z.J. Epigenetic modification of gene expression in honey bees by heterospecific gland secretions. PLoS One 2012, 7, e43727. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Jia, Q.; Li, F.; Han, Z. Identification of two PIWI genes and their expression profile in honeybee, Apis mellifera. Arch. Insect Biochem. Physiol. 2010, 74, 91–102. [Google Scholar] [PubMed]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [PubMed]
- Humann, F.C.; Tiberio, G.J.; Hartfelder, K. Sequence and expression characteristics of long noncoding RNAs in honey bee caste development—Potential novel regulators for transgressive ovary size. PLoS One 2013, 8, e78915. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cridge, A.G.; Leask, M.P.; Duncan, E.J.; Dearden, P.K. What Do Studies of Insect Polyphenisms Tell Us about Nutritionally-Triggered Epigenomic Changes and Their Consequences? Nutrients 2015, 7, 1787-1797. https://doi.org/10.3390/nu7031787
Cridge AG, Leask MP, Duncan EJ, Dearden PK. What Do Studies of Insect Polyphenisms Tell Us about Nutritionally-Triggered Epigenomic Changes and Their Consequences? Nutrients. 2015; 7(3):1787-1797. https://doi.org/10.3390/nu7031787
Chicago/Turabian StyleCridge, Andrew G., Megan P. Leask, Elizabeth J. Duncan, and Peter K. Dearden. 2015. "What Do Studies of Insect Polyphenisms Tell Us about Nutritionally-Triggered Epigenomic Changes and Their Consequences?" Nutrients 7, no. 3: 1787-1797. https://doi.org/10.3390/nu7031787