NF-κBp65 and Expression of Its Pro-Inflammatory Target Genes Are Upregulated in the Subcutaneous Adipose Tissue of Cachectic Cancer Patients

Abstract
:1. Introduction
2. Experimental Section
2.1. Patient Recruitment
2.2. Clinical and Biochemical Parameters Assessment
2.3. Adipose Tissue Biopsies
2.4. Gene Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense (5′–3′) | Antisense (5′–3′) |
|---|---|---|
| RPL-27 (NM_000988.3) | CCGAAATGGGCAAGTTCAT | CCATCATCAATGTTCTTCACGA |
| NF-κBp65 (NM_021975.3) | CCTGGAGCAGGCTATCAGTC | ATGGGATGAGAAAGGACAGG |
| NF-κBp50 (NM_003998.3) | CATCCCATGGTGGACTACCT | TGGGTCCAGCAGTTACAGTG |
| IκB-α (NM_020529.2) | CTCCGAGACTTTCGAGGAAATAC | GCCATTGTAGTTGGTAGCCTTCA |
| IL-1β (NM_000576.2) | AGCCAATCTTCATTGCTCAAGT | AGTCATCCTCATTGCCACTGT |
| IL-6 (NM_000600.3) | CAGCCCTGAGAAAGGAGACAT | AGCCATCTTTGGAAGGTTCA |
| TNF-α (NM_000594.3) | CTCTCTCCCCTGGAAAGGAC | ATCACTCCAAAGTGCAGCAG |
| INF-γ (NM_000619.2) | TGGAAAGAGGAGAGTGACAGAA | TGGAAAGAGGAGAGTGACAGAA |
| MCP-1 (NM_002982.3) | TCAGCCAGATGCAATCAATG | ACACTTGCTGCTGGTGATTCT |
2.5. NF-κB Binding Assay
2.6. Statistical Analysis
| N | T | TC | p | |
|---|---|---|---|---|
| n | 12 | 11 | 12 | |
| Male/Female (n) | 9/3 | 7/4 | 6/6 | |
| Age (years) | 62.00 ± 2.51 | 58.64 ± 4.04 | 60.42 ± 2.93 | 0.7609 |
| Height (m) | 1.65 ± 0.03 | 1.64 ± 0.02 | 1.64 ± 0.02 | 0.9909 |
| Previous body mass (Kg) | 75.48 ± 4.86 | 75.64 ± 4.532 | 74.44 ± 2.665 | 0.9728 |
| Current body mass (Kg) | 75.48 ± 4.86 | 67.83 ± 3.87 | 64.45 ± 2.98 | 0.1432 |
| Δ Body mass (%) | 0.00 ± 0.00 | 9.36 ± 3.27 * | 13.58 ± 1.75 * | 0.0005 |
| BMI (kg/m2) | 27.76 ± 1.40 | 25.31 ± 1.58 | 23.89 ± 1.16 | 0.1573 |
| Tumor stage | ||||
| I | - | 18.2% | 0% | - |
| IIA/IIB/IIC | - | 27.3% | 25% | - |
| IIIA/IIIB/IIIC | - | 45.4% | 33.3% | - |
| IVA/IVB | - | 9.1% | 41.7% | - |
| Primary tumour site | ||||
| Colon and rectum | - | 72.7% | 58.3% | - |
| Stomach | - | 18.2% | 41.7% | - |
| Other | - | 9.1% | 0% | - |
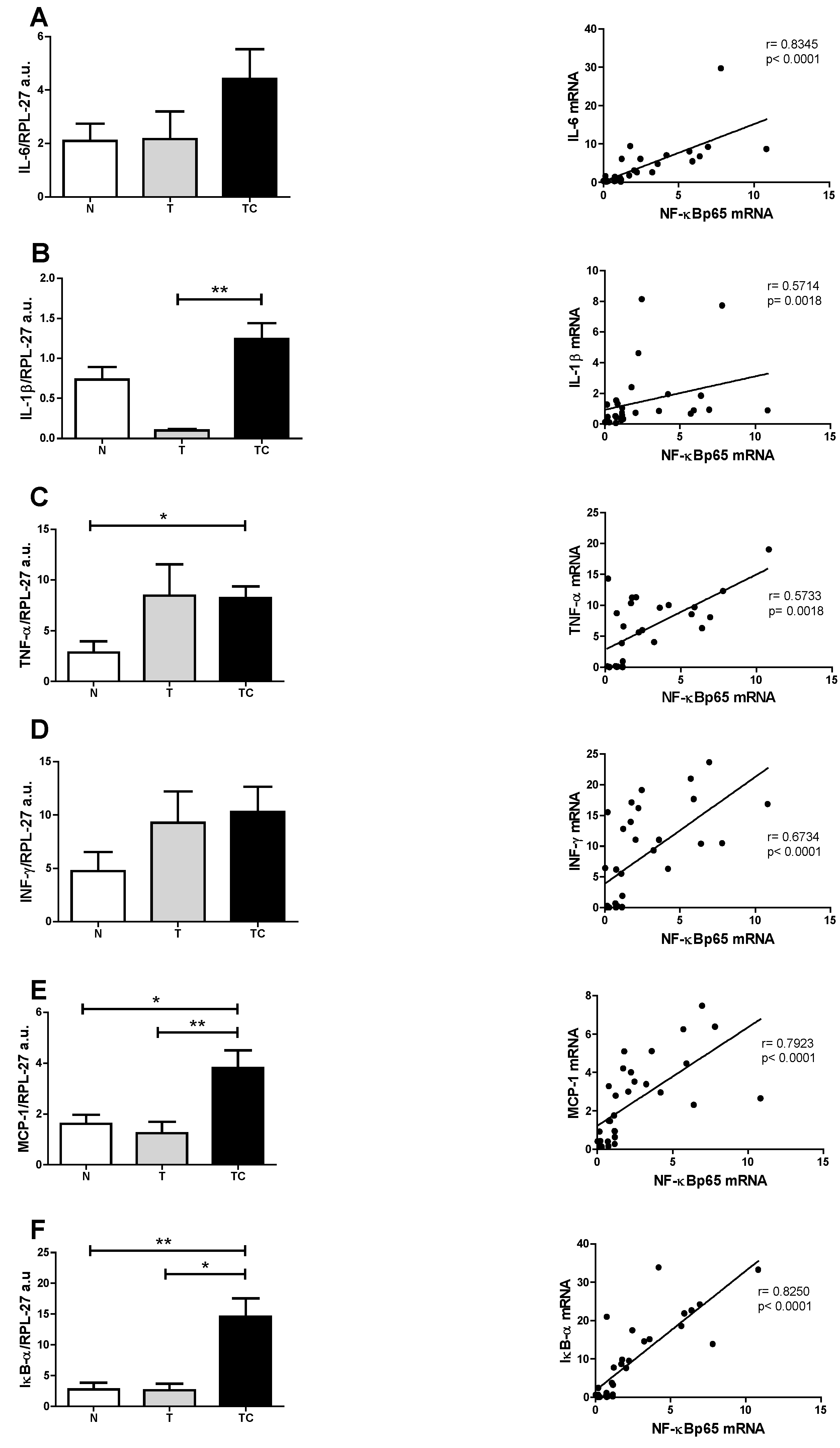
| Gene Expression | Statistical Analysis | Significance |
|---|---|---|
| (A) NF-κBp65 | p = 0.0147 | TC vs. T; TC vs. N |
| (B) NF-κBp50 | p = 0.1719 | --- |
| (C) IL-6 | p = 0.1458 | --- |
| (D) IL-1β | p = 0.0049 | TC vs. T |
| (E) TNF-α | p = 0.0201 | TC vs. N |
| (F) INF-γ | p = 0.2255 | --- |
| (G) MCP-1 | p = 0.0033 | TC vs. T; TC vs. N |
| (H) IkB-α | p = 0.0019 | TC vs. T; TC vs. N |



3. Results
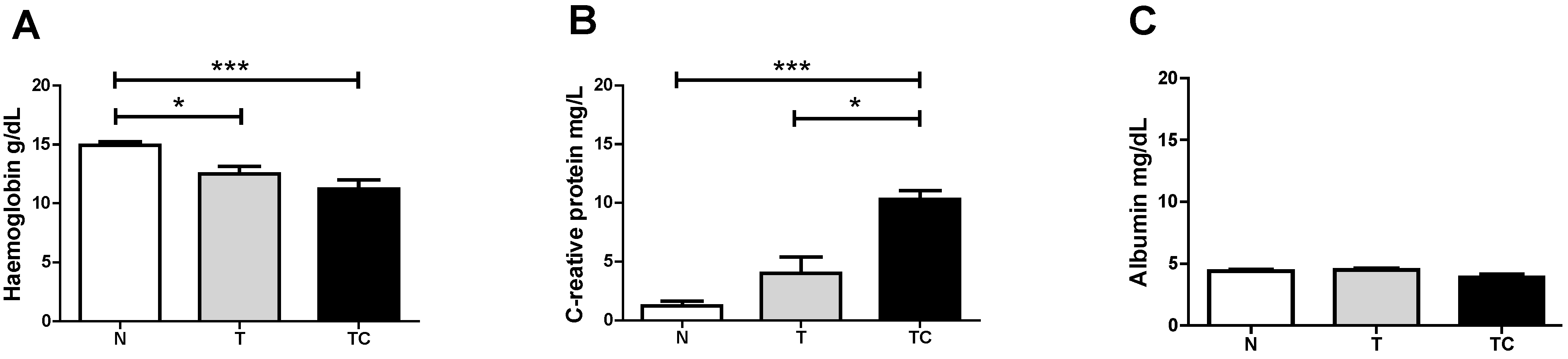
3.1. Clinical Findings
3.2. Quality of Life Analysis

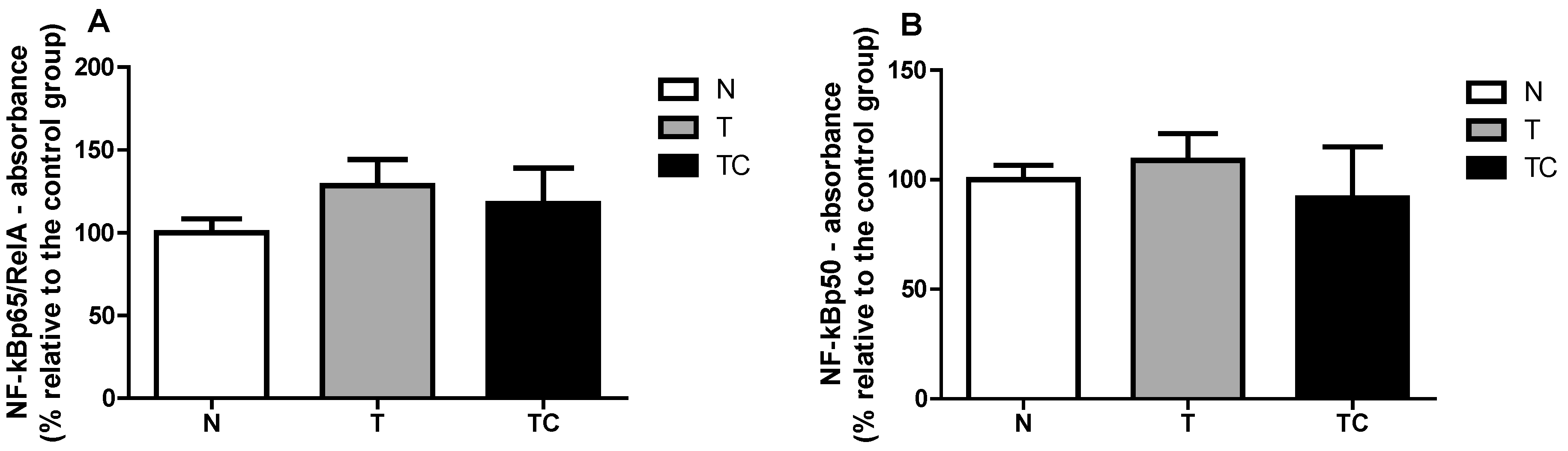
3.3. Gene Expression
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix
NF-κB Binding Assay


References
- Fearon, K.C.; Moses, A.G. Cancer cachexia. Int. J. Cardiol. 2002, 85, 73–81. [Google Scholar] [CrossRef]
- Clay, W.D. Balancing the scales: A common-sense look at global nutrition problems and what can be done about them. World Rev. Nutr. Diet. 2008, 98, 179–197. [Google Scholar] [PubMed]
- Barber, M.D.; Ross, J.A.; Fearon, K.C. Cancer cachexia. Surg. Oncol. 1999, 8, 133–141. [Google Scholar] [CrossRef]
- Lira, F.S.; Rosa, J.C.; Zanchi, N.E.; Yamashita, A.S.; Lopes, R.D.; Lopes, A.C.; Batista, M.L., Jr.; Seelaender, M. Regulation of inflammation in the adipose tissue in cancer cachexia: Effect of exercise. Cell Biochem. Funct. 2009, 27, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Pajak, B.; Orzechowska, S.; Pijet, B.; Pijet, M.; Pogorzelska, A.; Gajkowska, B.; Orzechowski, A. Crossroads of cytokine signaling—The chase to stop muscle cachexia. J. Physiol. Pharmacol. 2008, 59 (Suppl. 9), 251–264. [Google Scholar] [PubMed]
- Machado, A.P.; Costa Rosa, L.F.; Seelaender, M.C. Adipose tissue in Walker 256 tumour-induced cachexia: Possible association between decreased leptin concentration and mononuclear cell infiltration. Cell Tissue Res. 2004, 318, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Batista, M.L., Jr.; Peres, S.B.; McDonald, M.E.; Alcantara, P.S.; Olivan, M.; Otoch, J.P.; Farmer, S.R.; Seelaender, M. Adipose tissue inflammation and cancer cachexia: Possible role of nuclear transcription factors. Cytokine 2012, 57, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Booth, A.; Magnuson, A.; Fouts, J.; Foster, M. Adipose tissue, obesity and adipokines: Role in cancer promotion. Horm. Mol. Biol. Clin. Investig. 2015, 21, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P.; Wood, I.S. Adipokines: Inflammation and the pleiotropic role of white adipose tissue. Br. J. Nutr. 2004, 92, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Arner, P. Medicine. Lipases in cachexia. Science 2011, 333, 163–164. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J.; Karin, M. Nuclear factor-kappaB: A pivotal transcription factor in chronic inflammatory diseases. N. Engl. J. Med. 1997, 336, 1066–1071. [Google Scholar] [PubMed]
- Fraser, C.C. Exploring the positive and negative consequences of NF-kappaB inhibition for the treatment of human disease. Cell Cycle 2006, 5, 1160–1163. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Aggarwal, B.B. Nuclear transcription factor-kappaB as a target for cancer drug development. Leukemia 2002, 16, 1053–1068. [Google Scholar] [CrossRef] [PubMed]
- Laurencikiene, J.; van Harmelen, V.; Arvidsson Nordstrom, E.; Dicker, A.; Blomqvist, L.; Naslund, E.; Langin, D.; Arner, P.; Ryden, M. NF-kappaB is important for TNF-alpha-induced lipolysis in human adipocytes. J. Lipid Res. 2007, 48, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Saccani, A.; Schioppa, T.; Porta, C.; Biswas, S.K.; Nebuloni, M.; Vago, L.; Bottazzi, B.; Sica, A.; Colombo, M.P.; Mantovani, A. p50 nuclear factor-kappaB overexpression in tumor-associated macrophages inhibits M1 inflammatory responses and antitumor resistance. Cancer Res. 2006, 66, 11432–11440. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.J.; Morley, J.E.; Argiles, J.; Bales, C.; Baracos, V.; Guttridge, D.; Jatoi, A.; Lochs, H.; Kalantar-Zadeh, K.; Mantovani, G.; et al. Cachexia: A new definition. Clin. Nutr. 2008, 27, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Aaronson, N.K.; Ahmedzai, S.; Bergman, B.; Bullinger, M.; Cull, A.; Duez, N.J.; Filiberti, A.; Flechtner, H.; Fleishman, S.B.; Haes, J.C.J.M.; et al. The European Organization for Research and Treatment of Cancer QLQ-C30: A Quality-of-Life Instrument for Use in International Clinical Trials in Oncology. J. Natl. Inst. 1986, 85, 365–376. [Google Scholar] [CrossRef]
- Pais-Ribeiro, J.; Pinto, C.; Santos, C. Validation studyof the portuguese version of the QLC-C30-V.3. Psicol. Saúde Doenças 2008, 9, 89–102. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Richards, C.H.; Roxburgh, C.S.D.; McMillan, M.T.; Isswiasi, S.; Robertson, E.G.; Guthrie, G.K.; Horgan, P.G.; McMillan, D.C. The Relationships between Body Composition and the Systemic Inflammatory Response in Patients with Primary Operable Colorectal Cancer. PLoS ONE 2012, 7, e41883. [Google Scholar] [CrossRef] [PubMed]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Argiles, J.M.; Moore-Carrasco, R.; Fuster, G.; Busquets, S.; Lopez-Soriano, F.J. Cancer cachexia: The molecular mechanisms. Int. J. Biochem. Cell Biol. 2003, 35, 405–409. [Google Scholar] [CrossRef]
- McMillan, D.C. Systemic inflammation, nutritional status and survival in patients with cancer. Curr. Opin. Clin. Nutr. Metab.Care 2009, 12, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Kim, J.H.; Kannappan, R.; Reuter, S.; Dougherty, P.M.; Aggarwal, B.B. Role of nuclear factor kappaB-mediated inflammatory pathways in cancer-related symptoms and their regulation by nutritional agents. Exp. Biol. Med. 2011, 236, 658–671. [Google Scholar] [CrossRef] [PubMed]
- Deans, C.; Wigmore, S.J. Systemic inflammation, cachexia and prognosis in patients with cancer. Curr. Opin. Clin. Nutr. Metab.Care 2005, 8, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Sen, R.; Baltimore, D. Inducibility of kappa immunoglobulin enhancer-binding protein Nf-kappa B by a posttranslational mechanism. Cell 1986, 47, 921–928. [Google Scholar] [CrossRef]
- Schwartz, S.A.; Hernandez, A.; Mark Evers, B. The role of NF-kappaB/IkappaB proteins in cancer: Implications for novel treatment strategies. Surg. Oncol. 1999, 8, 143–153. [Google Scholar] [CrossRef]
- Rhoads, M.G.; Kandarian, S.C.; Pacelli, F.; Doglietto, G.B.; Bossola, M. Expression of NF-kappaB and IkappaB proteins in skeletal muscle of gastric cancer patients. Eur. J. Cancer 2010, 46, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Op den Kamp, C.M.; Langen, R.C.; Snepvangers, F.J.; de Theije, C.C.; Schellekens, J.M.; Laugs, F.; Dingemans, A.M.; Schols, A.M. Nuclear transcription factor kappa B activation and protein turnover adaptations in skeletal muscle of patients with progressive stages of lung cancer cachexia. Am. J. Clin. Nutr. 2013, 98, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Frantz, J.D.; Tawa, N.E., Jr.; Melendez, P.A.; Oh, B.C.; Lidov, H.G.; Hasselgren, P.O.; Frontera, W.R.; Lee, J.; Glass, D.J.; et al. IKKbeta/NF-kappaB activation causes severe muscle wasting in mice. Cell 2004, 119, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Jiang, Z.W.; Tian, J.; Jiang, J.; Li, N.; Li, J.S. Role of NF-κB and cytokine in experimental cancer cachexia. World J. Gastroenterol. 2003, 9, 1567–1570. [Google Scholar] [PubMed]
- Arner, P.; Langin, D. Lipolysis in lipid turnover, cancer cachexia, and obesity-induced insulin resistance. Trends Endocrinol. Metab. 2014, 25, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Batista, M.L., Jr.; Olivan, M.; Alcantara, P.S.; Sandoval, R.; Peres, S.B.; Neves, R.X.; Silverio, R.; Maximiano, L.F.; Otoch, J.P.; Seelaender, M. Adipose tissue-derived factors as potential biomarkers in Cachectic cancer patients. Cytokine 2013, 61, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Meng, T.; Murray, N.R.; Fields, A.P.; Brasier, A.R. Interleukin-1-induced nuclear factor-kappaB-IkappaBalpha autoregulatory feedback loop in hepatocytes. A role for protein kinase calpha in post-transcriptional regulation of ikappabalpha resynthesis. J. Biol. Chem. 1999, 274, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Argiles, J.M.; Busquets, S.; Lopez-Soriano, F.J. Anti-inflammatory therapies in cancer cachexia. Eur. J. Pharmacol. 2011, 668 (Suppl. 1), S81–S86. [Google Scholar] [CrossRef] [PubMed]
- Le Bail, O.; Schmidt-Ullrich, R.; Israel, A. Promoter analysis of the gene encoding the I kappa B-alpha/MAD3 inhibitor of NF-kappa B: Positive regulation by members of the rel/NF-kappa B family. EMBO J. 1993, 12, 5043–5049. [Google Scholar] [PubMed]
- Narsale, A.A.; Enos, R.T.; Puppa, M.J.; Chatterjee, S.; Murphy, E.A.; Fayad, R.; Pena, M.O.; Durstine, J.L.; Carson, J.A. Liver Inflammation and Metabolic Signaling in ApcMin/+ Mice: The Role of Cachexia Progression. PLoS ONE 2015, 10, e0119888. [Google Scholar] [CrossRef] [PubMed]
- McClung, J.M.; Judge, A.R.; Powers, S.K.; Yan, Z. p38 MAPK links oxidative stress to autophagy-related gene expression in Cachectic muscle wasting. Am. J. Physiol. Cell Physiol. 2010, 298, C542–C549. [Google Scholar] [CrossRef] [PubMed]
- Moore-Carrasco, R.; Garcia-Martinez, C.; Busquets, S.; Ametller, E.; Barreiro, E.; Lopez-Soriano, F.J.; Argiles, J.M. The AP-1/CJUN signaling cascade is involved in muscle differentiation: Implications in muscle wasting during cancer cachexia. FEBS Lett. 2006, 580, 691–696. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camargo, R.G.; Riccardi, D.M.d.R.; Ribeiro, H.Q.T.; Carnevali, L.C., Jr.; De Matos-Neto, E.M.; Enjiu, L.; Neves, R.X.; Lima, J.D.C.C.; Figuerêdo, R.G.; De Alcântara, P.S.M.; et al. NF-κBp65 and Expression of Its Pro-Inflammatory Target Genes Are Upregulated in the Subcutaneous Adipose Tissue of Cachectic Cancer Patients. Nutrients 2015, 7, 4465-4479. https://doi.org/10.3390/nu7064465
Camargo RG, Riccardi DMdR, Ribeiro HQT, Carnevali LC Jr., De Matos-Neto EM, Enjiu L, Neves RX, Lima JDCC, Figuerêdo RG, De Alcântara PSM, et al. NF-κBp65 and Expression of Its Pro-Inflammatory Target Genes Are Upregulated in the Subcutaneous Adipose Tissue of Cachectic Cancer Patients. Nutrients. 2015; 7(6):4465-4479. https://doi.org/10.3390/nu7064465
Chicago/Turabian StyleCamargo, Rodolfo Gonzalez, Daniela Mendes dos Reis Riccardi, Henrique Quintas Teixeira Ribeiro, Luiz Carlos Carnevali, Jr., Emidio Marques De Matos-Neto, Lucas Enjiu, Rodrigo Xavier Neves, Joanna Darck Carola Correia Lima, Raquel Galvão Figuerêdo, Paulo Sérgio Martins De Alcântara, and et al. 2015. "NF-κBp65 and Expression of Its Pro-Inflammatory Target Genes Are Upregulated in the Subcutaneous Adipose Tissue of Cachectic Cancer Patients" Nutrients 7, no. 6: 4465-4479. https://doi.org/10.3390/nu7064465