Polyphenols and Glycemic Control



School of Pharmacy and Medical Science, University of South Australia, General Post Office Box 2471 Adelaide SA 5000, Australia
*
Author to whom correspondence should be addressed.
Nutrients 2016, 8(1), 17; https://doi.org/10.3390/nu8010017
Submission received: 27 November 2015
/
Revised: 17 December 2015
/
Accepted: 18 December 2015
/
Published: 5 January 2016
{kind=link}
{kind=link}
Abstract
:Growing evidence from animal studies supports the anti-diabetic properties of some dietary polyphenols, suggesting that dietary polyphenols could be one dietary therapy for the prevention and management of Type 2 diabetes. This review aims to address the potential mechanisms of action of dietary polyphenols in the regulation of glucose homeostasis and insulin sensitivity based on in vitro and in vivo studies, and to provide a comprehensive overview of the anti-diabetic effects of commonly consumed dietary polyphenols including polyphenol-rich mixed diets, tea and coffee, chocolate and cocoa, cinnamon, grape, pomegranate, red wine, berries and olive oil, with a focus on human clinical trials. Dietary polyphenols may inhibit α-amylase and α-glucosidase, inhibit glucose absorption in the intestine by sodium-dependent glucose transporter 1 (SGLT1), stimulate insulin secretion and reduce hepatic glucose output. Polyphenols may also enhance insulin-dependent glucose uptake, activate 5′ adenosine monophosphate-activated protein kinase (AMPK), modify the microbiome and have anti-inflammatory effects. However, human epidemiological and intervention studies have shown inconsistent results. Further intervention studies are essential to clarify the conflicting findings and confirm or refute the anti-diabetic effects of dietary polyphenols.
1. Introduction
Type 2 diabetes (T2D) is a chronic disease of metabolic dysregulation, most notably abnormal glucose metabolism, accompanied by complications including cardiovascular disease, retinopathy, nephropathy, neuropathy, leg ulcers and gangrene [1]. In 2013, 382 million people were estimated to have diabetes globally and this number is projected to be 592 million by 2035 [2]. Identification of modifiable lifestyle factors including dietary factors that reduce the incidence of T2D is a vital area of research [3,4]. One dietary factor of interest is consumption of polyphenol-rich foods. Dietary polyphenols have been suggested to lower the risk of T2D [5,6,7].
This review aims to summarize relevant epidemiological and clinical studies linking polyphenol-rich foods to the risk of T2D, and to describe the multiple mechanisms of action of polyphenol-rich foods through which these effects are mediated.
2. Polyphenols
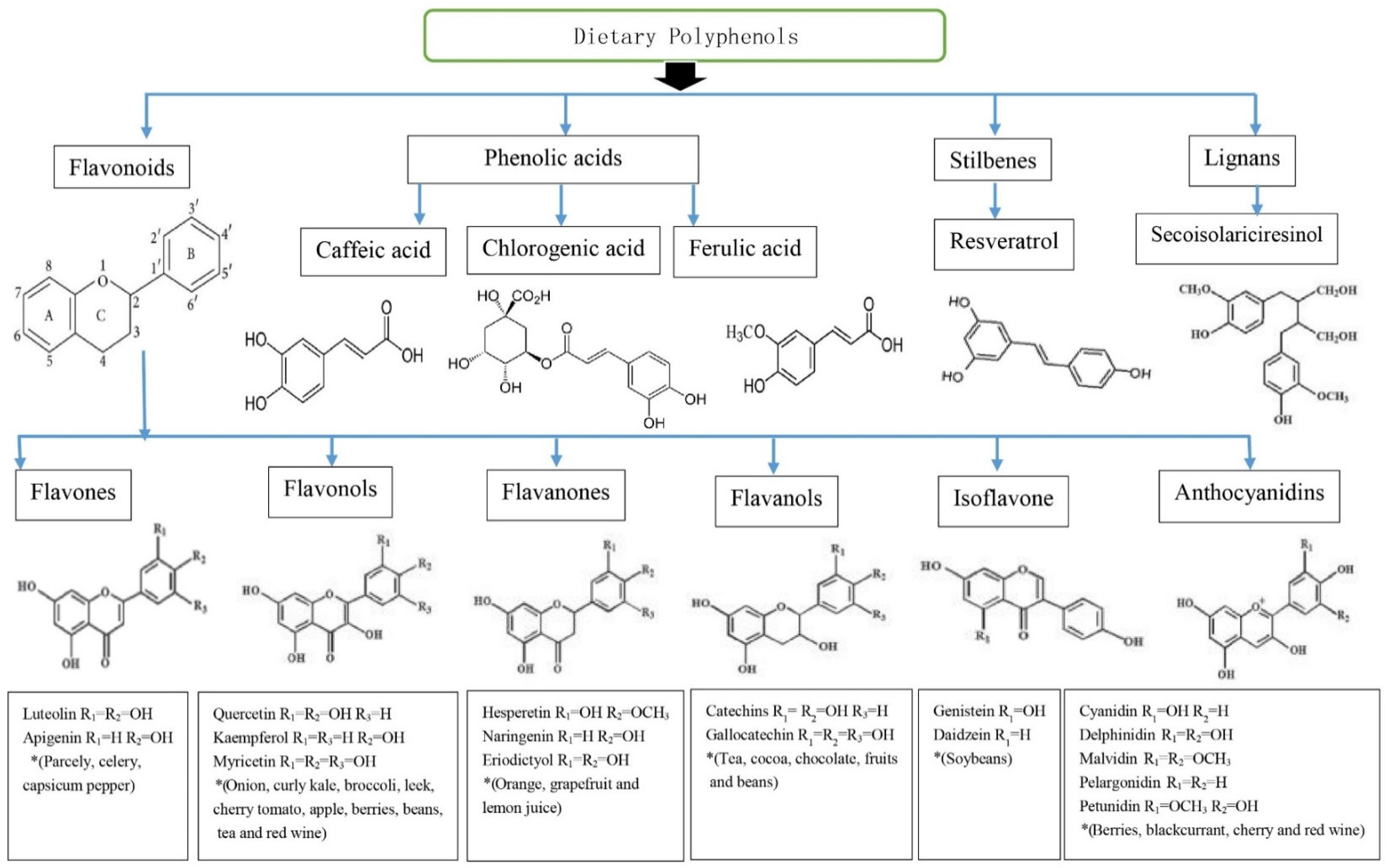
Polyphenols are a large and heterogeneous group of phytochemicals containing phenol rings [6]. Several hundred different polyphenols are found in plant-based foods including vegetables (particularly, broccoli, onion and cabbage), fruits (grapes, pears, apples, cherries and various berries contain up to 200–300 mg polyphenols per 100 g fresh weight), legumes (soybean), cereals, plant-derived beverages and chocolate [5,8,9]. Approximately 100 mg of polyphenols are identified in a cup of coffee or tea or a glass of red wine [8,9]. Polyphenols are divided into flavonoids, phenolic acids, stilbenes, and lignans [9]. Flavonoids include flavones, flavonols flavanols, flavanones, isoflavones, and anthocyanins [7,8,10]. The most common phenolic acids are caffeic acid (present in many fruits and vegetables, most often esterified with quinic acid as in chlorogenic acid, which is a major phenolic compound in coffee) and ferulic acid present in cereals which is esterified to hemicelluloses in the cell wall. The best studied stilbene is resveratrol in grapes, grape products and red wine [10]. The richest source of lignans is linseed present mainly as secoisolariciresinol (up to 3.7 g/kg dry weight) with low quantities of matairesinol, while minor sources are cereals, lentils, fruits (pears, prunes) and vegetables (garlic, asparagus, carrots) [10]. Chemical structures and dietary sources of different groups of polyphenols are shown in Figure 1.
Figure 1.
Chemical structures and dietary sources of different groups of polyphenols. Flavonoids are most abundant in four groups of dietary polyphenols and share a basic structure. Resveratrol is one of a subclass of stilbenes. Some phenolic acids are caffeic acid, chlorogenic acid and ferulic acid. Flavones, flavonols, flavanones, flavanols, isoflavone and anthocyanidins are main subclasses of flavonoids. Individual compound of those are characterised in accordance with the arrangement and the number of the hydroxyl groups and their extent of alkylation and/or glycosylation [9]. * Food sources from six main subclasses of flavonoids are briefly described. Typically, quercetin is rich in onion, tea, and apple. Hesperetin is rich in citrus fruits, genistein and daidzein are rich in soybeans. Anthocyanins including cyanidins contribute colour to many red fruits such as strawberry, raspberry and blackcurrant [8,10].
Figure 1.
Chemical structures and dietary sources of different groups of polyphenols. Flavonoids are most abundant in four groups of dietary polyphenols and share a basic structure. Resveratrol is one of a subclass of stilbenes. Some phenolic acids are caffeic acid, chlorogenic acid and ferulic acid. Flavones, flavonols, flavanones, flavanols, isoflavone and anthocyanidins are main subclasses of flavonoids. Individual compound of those are characterised in accordance with the arrangement and the number of the hydroxyl groups and their extent of alkylation and/or glycosylation [9]. * Food sources from six main subclasses of flavonoids are briefly described. Typically, quercetin is rich in onion, tea, and apple. Hesperetin is rich in citrus fruits, genistein and daidzein are rich in soybeans. Anthocyanins including cyanidins contribute colour to many red fruits such as strawberry, raspberry and blackcurrant [8,10].

3. Epidemiological Studies of Dietary Polyphenols
Epidemiological studies support favourable effects of polyphenol-rich diets in preventing and managing T2D [6].
3.1. Flavonoids
A prospective study in 1111 T2D case-control pairs in the Nurses’ Health Study I and II (NHS and NHSII) investigating urinary excretion of eight polyphenol metabolites found that flavanones (naringenin and hesperetin) and flavonols (quercetin and isorhamnetin), as well as the phenolic acid, caffeic acid, were associated with a 39%–48% lower T2D risk during early follow-up (median of ≤4.6 years since urine sample collection), but not during later follow-up which ranged from 4.6 to 11.4 years [11]. Metabolites of flavan-3-ols and ferulic acid were not associated with T2D risk in either period [11]. This is in accordance with the Women’s Health Study of 38,018 US women aged ≥45 years with an average 8.8 years of follow-up which reported that none of total flavonols and flavones, or quercetin, kaempferol, myricetin, apigenin, and luteolin were significantly associated with the risk of T2D [12]. However, apple consumption of ≥1 apple/day showed a 28% lower T2D risk compared with no apple consumption (relative risk—RR 0.72; 95% confidence interval (CI) 0.56–0.92; p = 0.006) [12]. A Finnish study of 10,054 men and women with 526 T2D cases showed that intakes of apples (hazard ratio—HR 0.73; 95% CI 0.57–0.92; p = 0.003) and berries (HR 0.74; 95% CI 0.58–0.95; p = 0.03) were significantly associated with a lower risk of T2D [13]. These findings are consistent with prospective studies of 159,560 women in the NHS and NHS II, and 41,334 men in the Health Professionals Follow-Up Study [14], which also found that higher intakes of anthocyanins were significantly associated with a lower risk of T2D (HR 0.85; 95% CI 0.80–0.91; p < 0.001) [14]. Apples/pears (HR 0.77; ≥5 servings/week vs. <1 serving/month; 95% CI 0.65–0.83; p < 0.001) and blueberries (HR 0.77; ≥2 servings/week vs. <1 serving/month; 95% CI 0.68–0.87; p < 0.001) were inversely associated with T2D [14], but total flavonoid intake or other flavonoid subclasses were not associated with a lower risk of T2D [14]. An inverse association remained after adjustment for body mass index (BMI) or weight [13,14].
The Iowa Women’s Health Study of mainly white, postmenopausal women did not observe any T2D-protective effect of total flavonoid intake or any of the flavonoid subclasses, including anthocyanins. On the other hand, regular red wine consumption of ≥1 occasion/week was associated with a 16% lower T2D risk than wine consumption <1 occasion/week (HR 0.84; 95% CI 0.71–0.99), with similar findings for white wine, beer, and liquor [15], suggesting it is the alcohol per se and not the polyphenols which is associated with protection. Inconsistent findings might be explained due to the use of the older, less complete versions of the United States Department of Agriculture (USDA) database [12,15] and misclassification of intake in the baseline questionnaires [14]. Much of the epidemiological data is based on reported food intake rather than blood or urine measures of polyphenol metabolites and is thus relatively unreliable compared with more objective measures.
3.2. Phenolic Acids
Epidemiological studies showed a fairly consistent association between coffee (caffeinated and decaffeinated) [16,17,18,19,20] or green tea [21] consumption and a lower risk of T2D in a dose-response manner in most studies [16,17,18,19,20,21] except [22].
A meta-analysis of 28 prospective studies with 1,109,272 participants and 45,335 T2D cases showed an inverse association of coffee consumption with T2D in a dose-response manner (1–6 cups/day) [16]. Coffee consumption of six cups/day was associated with a 33% lower risk of T2D, compared with no coffee consumption (RR 0.67; 95% CI 0.61–0.74; p < 0.001). Both caffeinated and decaffeinated coffee were associated with a lower risk of T2D [16]. A retrospective cohort study of 17,413 Japanese men and women aged 40–65 years, based on completed five-year follow-up questionnaire found RRs of 0.67 (95% CI 0.47–0.94) for green tea intake of ≥6 cups/day and 0.58 (95% CI 0.37–0.90) for coffee intake of ≥3 cups/day, compared with less than one cup/week [21]. The consumption of black or oolong teas was not associated with the risk of T2D [21]. Total caffeine intake was associated with a 33% lower risk of T2D [21]. These inverse associations were more prominent in women and in overweight men [21]. Similarly, another study [17] also observed a stronger inverse association between coffee, decaffeinated coffee and caffeine intake, and T2D incidence in women than in men as well as a stronger inverse association between coffee intake and T2D incidence in non-smokers and subjects with BMI <25 kg/m2 [17] compared with smokers and overweight and obese people. Three other meta-analyses showed that tea intakes of ≥3 cups/day (RR 0.84; 95% CI 0.73–0.97) [23] or ≥4 cups/day [24] had a 16% reduction in the risk of developing T2D, compared with the lowest intake (RR 0.84; 95% CI 0.73–0.94) but a third meta-analysis [25] showed no effect.
A prospective cohort of 40,011 participants with a mean follow-up of 10 years and 918 cases of T2D [22] reported potential beneficial effects of both coffee and tea intakes, but there was no dose-response as coffee and/or tea intakes of at least three cups/day reduced the risk of T2D by 42%. Coffee intake of 4.1–6.0 cups/day and tea intake of >5.0 cups/day reduced the risk of T2D by 23% (HR 0.77; 95% CI 0.63–0.95; p = 0.033) and 37% (HR 0.63; 95% CI 0.47–0.86; p = 0.002), respectively [22]. The relative amount of coffee vs. tea was not important [22].
Positive effects of coffee and tea on a lower risk of T2D were independent of weight or BMI [16,18,20,21,22,24].
Observational studies have shown that higher coffee consumption was associated with decreased fasting insulin [26] and 2-h post-load glucose [27], and homeostatic model assessment levels [26,28,29], increased insulin sensitivity [30], but other studies have shown no association of coffee consumption with fasting insulin [27,31] or homeostasis model assessment of insulin resistance (HOMA–IR) [31]. Green tea consumption was positively associated with HOMA–IR (p = 0.02) with no dose-response [28]. The Whitehall II study, a British cohort study, showed no association between coffee, tea and the risk of T2D, although there was an association between combined intake of tea and coffee (≥2 cups/day of both beverages) and the risk of T2D (HR 0.68; 95% CI 0.46–0.99; p < 0.05) [32].
4. Potential Mechanisms of Action of Polyphenols in Diabetes Risk
The hypoglycemic effects of dietary polyphenolic compounds may be related to inhibition of carbohydrate digestion by inhibiting salivary and pancreatic α-amylase and α-glucosidase in the small intestinal brush border, inhibition of glucose absorption, and stimulation of insulin secretion and protection of pancreatic β-cells against glucotoxicity. Polyphenols may suppress glucose release from the liver, and improve glucose uptake in peripheral tissues by modulating intracellular signalling [6]. Polyphenols have antioxidant activity and can inhibit advanced glycation endproduct (AGE) formation [5].
4.1. Carbohydrate Digestion and Glucose Absorption in the Intestine
The key enzymes involved in digestion of dietary carbohydrate are α-amylase and α-glucosidase. Maltose, maltotriose and α-dextrins are the three major products of α-amylase digestion. Further digestion occurs in the small intestine by α-glucosidase which is a class of brush-border bound enzymes which hydrolyse the terminal α-1, 4-linked glucose residues [6,33]. Glucose is taken into cells via transporters, predominately sodium-dependent glucose transporter (SGLT1) located in the brush border membrane at the apical side of enterocytes [34].
Inhibition of α-amylase and α-glucosidase activities in vitro has been demonstrated with dietary polyphenols from berries (strawberries, raspberries, blueberries and blackcurrants) [35], vegetables (pumpkin, beans, maize and eggplant) [36,37], black rice [38], legumes [39], green and black tea [33], tea polyphenols [40] and red wine [35,41]. Inhibition of glucose transport has been shown with flavonoids and phenolic acids [42,43,44,45,46].
4.2. Tissue Uptake of Glucose
Enhanced insulin-mediated glucose uptake in vitro, a glucose transporter 4-mediated process, has been shown with dietary polyphenols including epicatechin [47], epigallocatechin-3-O-gallate (EGCG) [48], grape seed-derived procyanidins [49,50], bitter melon [51], blueberry [52], canna indica root [53] and black soy bean [54]. Black soybean seed rich in anthocyanins (cyanidin 3-glucoside) and procyanidins (PCs) lowered glucose levels and improved insulin sensitivity by activation of 5′ adenosine monophosphate-activated protein kinase (AMPK) in the skeletal muscle and liver of type 2 diabetic mice. This activation was accompanied by the up-regulation of glucose transporter 4 GLUT4 in skeletal muscle and the down-regulation of gluconeogenesis in the liver in type 2 diabetic mice [54]. EGCG maintained insulin sensitivity in rat L6 muscle cells exposed to dexamethasone and dose-dependently increased glucose uptake and GLUT4 translocation through activation of phosphoinositide 3-kinase (PI3K) signalling and AMPK and PI3K activation [48]. Cinnamon has shown potential to promote insulin signalling, and GLUT4 translocation in Swiss albino mouse embryo fibroblast line 3T3-L1 adipocytes [55], and increase glucose uptake in insulin resistant rats induced by a high fructose diet [56,57].
4.3. Gut Microbiota
Only 5%–10% of the total intake of dietary polyphenols are directly absorbed through the stomach and the small intestine [58]. The majority of the ingested polyphenols reach the colon, thereafter undergoing intensive metabolism prior to absorption [58,59]. Some polyphenols are thought to exert a prebiotic effect by stimulating the growth and activity of some bacteria in the digestive tract [59]. After absorption, polyphenols undergo phase I and II biotransformation (sulfation, glucuronidation, methylation and glycine-conjugation) by enterocytes in the liver to increase hydrophilicity favouring urinary secretion [58]. Polyphenol metabolites derived from liver metabolism interact with adipose tissue, pancreas, muscle and liver, and may exert anti-diabetic effects [59]. Absorption of polyphenols can be affected by dosage, size of phenolic compound, prior diet, food matrix, gender and differences in the gut microbial populations [59].An increased level of faecal Bifidobacteria has been associated with improved glucose tolerance and diminished inflammatory markers such as the interleukins IL-6, IL-1α and IL-1β, tumor necrosis factor α and monocyte chemoattractant protein-1 [59,60,61]. A few clinical trials [62,63,64,65,66,67] have shown the potential prebiotic effects of dietary polyphenols to increase the population of Bifidobacteria. In a randomized, controlled, double-blind, crossover study of 22 health volunteers [62], the daily consumption of 494 mg cocoa flavanols for four weeks significantly increased the growth of Lactobacillus spp. (p < 0.001) and Bifidobacterium spp. (p < 0.01) relative to that of a drink containing only 29 mg flavanols and matched for fibre and other potential prebiotics.A significant reduction in C-reactive protein (CRP) (p < 0.05) was observed and changes in CRP were associated with changes in Lactobacilli counts (p < 0.05; R2 = 0.20) [62]. Similarly, the daily consumption of red wine polyphenols for four weeks modulated the growth of gut microbiota [66] by significantly increasing Bifidobacterium and Enterococcus, Prevotella, Bacteroides, Bacteroides uniformis, Eggerthella lenta, and Blautia coccoides-Eubacterium rectale groups compared with baseline but there was no control drink for comparison. Improvements in cholesterol and CRP levels were associated with changes in the Bifidobacteria amount [66]. Daily consumption of 25 g of wild blueberry powder drink for six weeks increased Bifidobacterium spp. compared with the placebo drink (250 mL of water, 7.5 g of fructose, 7 g of glucose, 0.5 g of citric acid, 0.03 g of blueberry flavour and colors) [63]. A proanthocyanidin-rich extract from grape seeds administered to nine healthy adults for two weeks significantly increased Bifidobacterium but no control material was provided to volunteers [64].
4.4. Pancreatic β-Cell Function
Glucose-stimulated insulin secretion from pancreatic β-cells involves glucose entrance via glucose transporter 2 (GLUT2), increased ATP content through glycolysis and tricarboxylic acid cycle (TCA) activity, and then inactivation of ATP-sensitive K+ (KATP) channels on the cell membrane [6,68]. Rat pancreatic β-cells, RIN-m5F treated with EGCG and the buckwheat flavonoid, rutin showed reduced glucotoxicity with activation of insulin receptor substrate 2 (IRS2) and AMPK signalling and elevated adenosine triphosphate (ATP) levels [69]. Activation of AMPK plays a critical role in the protection of β-cell against glucolipotoxicity [70]. The anti-diabetic drugs, metformin and thiazolidinediones (TZDs) are known to activate AMPK [69]. Chronic high glucose exposure directly increased intracellular reactive oxygen species (ROS) generation and disturbed mitochondrial function, uncoupling ATP generation and impairing insulin secretion [69]. Moreover, chronic high glucose exposure suppressed AMPK activity leading to increased lipogenesis [69]. This consequently resulted in impairing normal β-cell functioning [69].
Quercetin, apigenin, and luteolin inhibited cytokine-induced pancreatic β-cell damage through suppression of nuclear factor kappaB (NF-κB) activation in RINmF5 cells [71]. Quercetin showed protective effects by decreasing oxidative stress with preservation of pancreatic β-cell integrity in streptozotocin (STZ)-induced diabetes in rats [72]. Isolated rat islets exposed to (−) epicatechin (0.8 mmol/L) or quercetin (0.01 mmol/L) showed enhanced insulin secretion by approximately 44%–70% in the presence of 20 mmol/L glucose [73].
4.5. Liver Glucose Homeostasis
The liver is a key regulator of blood glucose levels in coordination with muscle and adipose tissues [6]. In the postprandial state, the liver stores glucose as glycogen via the glycogenesis pathway involving glucokinase (GK) which is a key enzyme in the regulation of glucose utilization in the liver, and glycogen synthase (GS). Under fasting conditions, the liver produces glucose via two routes—either glycogenolysis or gluconeogenesis involving pyruvate carboxylase, phosphoenolpyruvatecarboxykinase (PEPCK), fructose-1,6-bisphosphatase, and glucose-6-phosphatase [6].
Suppressed hepatic glucose production appears to account for decreased glucose levels in EGCG and green tea feeding and in vitro studies [74,75,76]. In the rodent study [76] of EGCC supplementation of either 0.25%–1% for seven weeks, a decrease of glucose levels in EGCC-treated db/db mice was observed in a dose dependent manner, compared with placebo-treated mice [76]. The GK mRNA expression was increased in the livers of mice supplemented with EGCC for seven weeks in a dose dependent manner and a decrease of PEPCK mRNA expression was observed in adipose tissue of EGCC supplemented mice. High-dose EGCC supplementation also increased acyl CoA oxidase-1 (ACO–1) and carnitine palmitoyl transferase-1β (CPT-1β) in both livers and adipose tissues [76]. Moreover, in H4IIE hepatoma cells, EGCC downregulated genes involved in gluconeogenesis and the synthesis of fatty acid, triacylglycerol and cholesterol, whereas genes involved in glycolysis (phosphofructokinase) and glucose transport (glucose transporter 1) were increased [76]. Supplementation of the soy isoflavones genistein and daidzein (0.02% in diet) improved glucose homeostasis in NOD mice with an increase in insulin/glucagon ratio and C-peptide level with preservation of insulin staining β-cell of pancreas. In the liver, glucose-6-phosphatase (G6Pase), PEPCK activities and β-oxidation of fatty acid were suppressed and lipogenesis was increased compared with the control group [77]. In db/db mice, the citrus flavonoids (hesperidin and naringin) were also found to lower glucose levels through up-regulating the hepatic GK via peroxisome proliferator-activated receptor gamma (PPAR γ), and upregulating adipocyte GLUT4. Naringin also suppressed PEPCK and G6Pase. This mechanism of citrus flavonoid is very similar to the mechanism of thiazolidinediones [78]. Similarly, areca nut procyanidins fed 10 mg/kg to multiple low dose streptozocin (MLD–STZ) treated mice for four weeks reduced fasting blood glucose, PEPCK and G6Pase and activated AMPKα [79].
In the fed state, dietary cyanidin 3-glucoside was shown to downregulate G6Pase and PEPCK by enhancing the phosphorylation of Akt and, forkhead box protein O1 (FOXO1—an important transcription factor) and by decreasing the nuclear translocation of FOXO1 in the liver and adipose tissues of high-fat diet-fed and db/db mice [80].
4.6. Inflammation
Low-grade inflammation and obesity-induced oxidative stress in the mitochondria may be partly responsible for insulin resistance. Elevated proinflammatory cytokines can alter metabolic homeostatic control systems leading to T2D [81]. Grape-seed procyanidins inhibited the production of proinflammatory molecules such as CRP, IL-6 and TNF-α and enhanced the production of adiponectin in fat fed rats [82]. Dietary cyanidin 3-glucoside (0.2% in diet) improved insulin sensitivity in both high-fat diet fed and obese db/db mice [80], decreased mRNA abundance of inflammatory cytokines including TNF-α, IL-6 and MCP-1 and suppressed macrophage infiltration in adipose tissue [80]. Procyanidins reduced gene expression of NF-κB, cyclooxygenase-2 protein (COX2), CRP, IL-6 and TNF-α in adipocytes [83,84], muscle [84] and liver [85,86] in both rats and mice. In male C57Bl/6j mice, eight-week supplementation of a high-fat diet with an anthocyanin-rich, 4% blueberry powder attenuated gene expression in macrophages in adipose tissue, for TNF-α, IL-6, MCP-1, inducible nitric oxide synthase, and improved whole body insulin sensitivity [87]. Quercetin also showed anti-inflammatory effects in male C57Bl/6j mice [88] and in obese Zucker rats [89]. Supplementation of a high-fat diet with 0.8% quercetin for eight weeks decreased interferon-γ, interleukin-1α, and interleukin-4 in male C57Bl/6j mice [88]. Administration of both 2 and 10 mg/kg of body weight of quercetin for 10 weeks improved insulin sensitivity and blood pressure in obese Zucker rats, but only the dose of 10 mg/kg reduced NOx concentration and visceral adipose tissue (VAT) TNF-α production, increased adiponectin and downregulated fat iNOS expression [89].
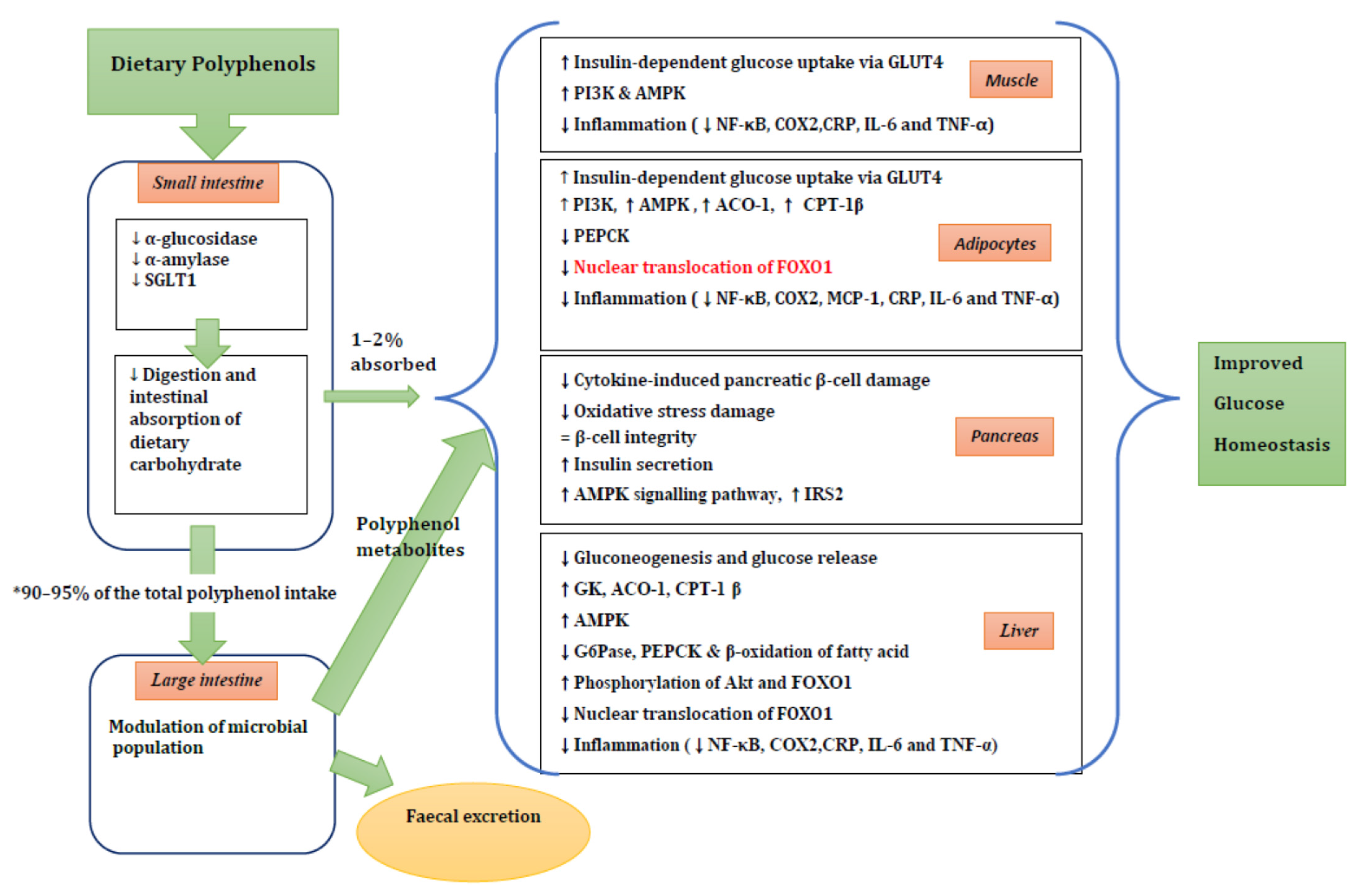
A summary of possible mechanisms of action is shown in Figure 2.
Figure 2.
The summary of potential mechanisms linking dietary polyphenol metabolites to improved glucose homeostasis. ↑, increase; ↓, decrease; ↔, maintenance of stability. * 90%–95% of the ingested polyphenols reach the colon. See the text for more details. SGLT1, sodium-dependent glucose transporter; GLUT4, glucose transporter 4; PI3K, phosphoinositide 3-kinase; AMPK, 5′ adenosine monophosphate-activated protein kinase; NF-κB, nuclear factor kappaB; COX2, cyclooxygenase-2 protein; CRP, C-reactive protein; IL-6, interleukin 6; TNFα, tumor necrosis factor α; ACO-1, acyl CoA oxidase-1; CPT-1 β, carnitine palmitoyl transferase-1β; PEPCK, phosphoenolpyruvate carboxykinase; FOXO1, forkhead box protein O1; MCP-1, monocyte chemoattractant protein-1; IRS2, insulin receptor substrate 2; GK, glucokinase; AC0-1, acyl CoA oxidase-1; G6Pase, glucose-6-phosphatase.
Figure 2.
The summary of potential mechanisms linking dietary polyphenol metabolites to improved glucose homeostasis. ↑, increase; ↓, decrease; ↔, maintenance of stability. * 90%–95% of the ingested polyphenols reach the colon. See the text for more details. SGLT1, sodium-dependent glucose transporter; GLUT4, glucose transporter 4; PI3K, phosphoinositide 3-kinase; AMPK, 5′ adenosine monophosphate-activated protein kinase; NF-κB, nuclear factor kappaB; COX2, cyclooxygenase-2 protein; CRP, C-reactive protein; IL-6, interleukin 6; TNFα, tumor necrosis factor α; ACO-1, acyl CoA oxidase-1; CPT-1 β, carnitine palmitoyl transferase-1β; PEPCK, phosphoenolpyruvate carboxykinase; FOXO1, forkhead box protein O1; MCP-1, monocyte chemoattractant protein-1; IRS2, insulin receptor substrate 2; GK, glucokinase; AC0-1, acyl CoA oxidase-1; G6Pase, glucose-6-phosphatase.

5. Clinical Studies of Dietary Polyphenols
There are a few clinical studies investigating the effects of specific polyphenols or food products rich in polyphenols in reducing the risk of insulin resistance [6].
5.1. Tea and Coffee
In a meta-analysis of green tea or green tea extract of seven randomised controlled trials, the consumption of green tea did not decrease the levels of fasting plasma glucose, fasting serum insulin, 2-h oral glucose tolerance test (OGTT), glucose, glycated hemoglobin (HbA1c) or HOMA–IR index [90]. However the authors stated that limitations of the studies included the small sample sizes, poor quality and low evidence levels of the included studies. In contrast, another meta-analysis of 17 randomised controlled trials including seven high quality studies and 10 low quality studies, demonstrated that green tea had favourable effects significantly reducing the fasting glucose and HbA1c by 0.09 mmol/L (95% CI 0.15–0.03 mmol/L; p < 0.01) and 0.30% (95% CI 0.37%–0.22%; p < 0.01), respectively. Subgroup analyses from studies with high Jadad scores showed a significant reduction in fasting insulin (−1.16 μIU/mL, 95% CI −1.91 to −0.40 μIU/mL; p = 0.03) [91]. This study attributed these findings to measures of glucose control or insulin sensitivity not coming from primary outcomes, limited high quality studies, and concealment of null findings in most selected studies [91].
A third meta-analysis of 22 randomized controlled trials with 1584 subjects showed that green tea catechins (EGCG capsules or powder, whole green tea or green tea extract powder) with or without caffeine reduced fasting glucose (−1.48 mg/dL; 95% CI −2.57 to −0.40 mg/dL), whereas fasting insulin, HbA1c, and HOMA–IR were not affected. In subgroup analyses, the glucose-lowering effect was observed when the duration of follow-up was over a median of 12 weeks. This finding suggested that longer trials were required to support favourable effects of green tea consumption on glycemic control [92].
In summary, given the mixed results from interventions in contrast to epidemiological studies, it is still unclear if polyphenol-rich green tea has anti-diabetic effects.
A randomized cross over study investigating the effect of a single dose of espresso caffeinated coffee, decaffeinated coffee, or water on glucose tolerance and insulin sensitivity (Matsuda index) during an OGTT in 18 habitual coffee drinkers with T2D showed no difference in area under the curve AUC insulin, glucose or insulin sensitivity [31]. However, a three-way, randomized, crossover study of nine healthy subjects [93] showed that chlorogenic acid in coffee acutely modulated gastrointestinal hormone secretion and glucose uptake [93]. The consumption of 25 g glucose in either 400 mL water (control) or 400 mL caffeinated or decaffeinated coffee (equivalent to 2.5 mmol chlorogenic acid/L), did not affect glycemia or insulinemia but attenuated the postprandial increase of glucose-dependent insulinotropic-polypeptide (GIP) (p < 0.005) while glucagon-like peptide 1 secretion increased 0–120 min (p < 0.01) with a delayed intestinal glucose absorption compared with the control [93]. This finding suggested that chlorogenic acid in coffee decreased the rate of intestinal absorption of glucose or slowed gastric empting [93].
Overall, there is little evidence available to assess the effect of coffee on glucose homeostasis.
5.2. Chocolate and Cocoa
Meta-analyses of acute or short-term, chronic (≤18 weeks), randomized controlled trials showed that chocolate or cocoa reduced fasting insulin (−2.65 µU/mL) and insulin after glucose challenge (−17 µU/mL at 30 min; 95% CI −20.7 to −13.4 µU/mL) and improved insulin resistance (HOMA–IR −0.67; 95% CI −0.98 to −0.36). There was no effect on fasting glucose and HbA1c [94]. Dark chocolate (100 g bar containing approximately 500 mg of polyphenols) for 15 days decreased blood pressure and improved insulin sensitivity in healthy individuals as demonstrated by decreasing HOMA–IR (0.94 ± 0.42 vs. 1.72 ± 0.62; p < 0.001) and increasing quantitative insulin sensitivity check index (QUICKI) (0.398 ± 0.039 vs. 0.356 ± 0.023; p = 0.001), compared with 90 g white chocolate bar containing no polyphenols [95], and similar results were shown in hypertensive [96] and glucose-intolerant hypertensive individuals [97]. A four-week consumption of dark chocolate containing 500 mg polyphenols reduced blood pressure (BP), fasting glucose and HOMA–IR in lean and overweight females [98] compared with 20 g of placebo dark chocolate containing negligible polyphenol but similar nutritional composition. Consumption of cocoa flavanols (902 mg) for 12 weeks also improved insulin sensitivity in overweight and obese individuals compared with a low-flavanol cocoa drink [99]. In contrast, consumption of flavanol-rich cocoa drink (150 mL twice a day, containing approximately 900 mg flavanols) for two weeks did not improve insulin resistance or blood pressure in people with essential hypertension [100]. Daily consumption of 25 g dark chocolate for eight weeks did not improve fasting glucose, insulin and HbA1c levels in hypertensive diabetic subjects compared with 25 g white chocolate consumption [101]. Similarly, in acute studies [102,103], dark chocolate containing 450 mg of polyphenols did not improve insulin and glucose in hypertensive diabetic subjects given a 75 g oral glucose load 60 min after ingestion of either low (control) or a high polyphenol dark chocolate [102], and cocoa beverage containing 960 mg total polyphenols and 480 mg flavanols showed no effect on glucose and increased insulin levels in obese subjects with T2D given a high-fat breakfast (766 kcal; 50 g fat) compared with a placebo (110 mg total polyphenols; <0.1 mg flavanols) [103].
In summary, given conflicting results in interventions, current data is insufficient to recommend chocolate and cocoa for glycemic control. Therefore, large placebo controlled trials with good quality designs are required to clarify the effects of cocoa polyphenols.
5.3. Cinnamon
Cinnamon is known to contain polyphenols such as catechin, procyanidin, cinnamtannin trans-cinnamic acid and flavones (cinnamaldehyde and trans-cinnamaldehyde) [5,104,105]. Eleven studies have shown positive effects of cinnamon on glycemic control. Three studies [106,107,108] included subjects with T2D with baseline HbA1c levels >8%. Seven studies [109,110,111,112,113,114,115] included healthy normal subjects [111,112,113,114], subjects with impaired fasting glucose [109] and subjects with metabolic syndrome [115]. One study showed beneficial effects on improved insulin sensitivity in patients with polycystic ovary syndrome [105]. Two studies [116,117] showed positive effects on fasting blood glucose levels, but did not report baseline HbA1c levels [116] and no significant changes of HbA1c, total cholesterol, low-density lipoprotein (LDL), high-density lipoprotein (HDL) or TG [117]. Five studies [118,119,120,121,122] showed no significant changes in fasting glucose, lipids, HbA1c, or insulin levels in 25 postmenopausal women with T2D taking 1.5 g of cinnamon daily for six weeks [120], in 43 subjects with T2D receiving 1 g of cinnamon daily for three months [118], in 60 subjects with T2D taking 1 g of cinnamon daily for 12 weeks [121], in 72 adolescents with Type 1 diabetes (T1D) taking 1 g of cinnamon daily [122] and in 11 healthy subjects taking 3 g of cinnamon daily for four weeks [119]. A randomized, controlled trial [108] (no placebo used with the concern that the strong taste of cinnamon capsules would preclude any blinding), investigating whether daily cinnamon plus usual care versus usual care alone lowers HbA1c in 109 patients with T2D (HbA1c > 7.0), showed that supplementation with 1 g of daily cassia cinnamon did not significantly lower HbA1c over 90 days [108]. A randomized, placebo-controlled, double-blind clinical trial of 58 subjects with T2D found that intake of 2 g daily of cinnamon for 12 weeks significantly reduced HbA1c (−0.36%; p < 0.005), systolic blood pressure—SBP (−4 mm Hg; p < 0.001) and diastolic blood pressure—DBP (−4 mmHg; p < 0.001) among 58 poorly controlled patients with T2D, compared with the placebo [107]. A randomized, double-blinded clinical study of 66 Chinese with T2D treated with gliclazide found significant reductions in HbA1c in those receiving 360 mg of cassia cinnamon extract (−0.92%; p = 0.0004) and 120 mg of cassia cinnamon extract (−0.67%; p = 0.003) after 90 days. No significant reductions were seen in the placebo group but the groups were not compared statistically. Fasting glucose levels were also significantly reduced over time in both treatment groups [106]. A study [116] in 60 subjects with T2D treated with sulfonylureas found significant reductions in fasting glucose (18%–29%), triglycerides (23%–30%), LDL (7%–27%), and total cholesterol (12%–26%), following supplementation with either 1, 3, or 6 g of cassia cinnamon daily for 40 days, while no change was observed in the placebo group but the groups again were not compared statistically. Significant effects were observed even after 20 days of washout [116]. A double-blind study [117] found that intake of 3 g daily cinnamon powder for 120 days significantly decreased fasting glucose but not HbA1c in 65 patients with T2D treated with oral antidiabetics or diet alone. Fasting glucose was reduced by 10.3% ± 13.2% in the treatment group and 3.37% ± 14.2% in the placebo group (p = 0.046). No significant changes in HbA1c, total cholesterol, low-density lipoprotein (LDL), high-density lipoprotein (HDL) or triglycerides (TG) were observed. Baseline HbA1c values were below 7% [117]. Two recently updated meta-analyses showed that the intake of cinnamon/cinnamon extracts lowered fasting glucose levels [123,124]. A meta-analysis of 10 randomized controlled trials (543 patients) reported that the consumption of cinnamon ranging from 120 mg/day to 6 g/day for 4–18 weeks decreased levels of fasting glucose, total cholesterol, LDL, and triglyceride and increased levels of HDL. No significant effect on HbA1c was observed [124]
In summary, the consumption of cinnamon may have glycaemic-lowering effects, but more intervention studies covering different types and amounts of cinnamon and types of subjects, are required as not all studies have shown positive effects of cinnamon on insulin resistance and many have been poorly analysed. The usual dose of cinnamon used was 1–2 g/day but some studies used larger doses of 6 g/day which would be difficult to maintain long term.
5.4. Grape Polyphenols
A double-blinded, randomized, crossover trial of 32 subjects with T2D showed positive effects of grape seed extract intake of 600 mg/day for four weeks on fructosamine, whole blood glutathione (GSH; p < 0.01), high sensitivity C reactive protein hsCRP, (p = 0.0006) and total cholesterol concentration (p = 0.05), but did not show significant changes in HOMA–IR [125]. Another study showed protective effects of grape polyphenols on fructose-induced oxidative stress and insulin resistance in first-degree relatives of patients with T2D. Subjects were randomised to eight weeks of supplementation with 2 g/day grape polyphenols or placebo and then ingested 3 g fructose/kg fat free mass/day as a 20% fructose solution with the three meals during the six days preceding muscle biopsies and a two-step hyperinsulinemia-euglycemic clamp. After six-day fructose loading, the placebo group showed a 20% decrease in hepatic insulin sensitivity (11.9 ± 1.5 vs. 9.6 ± 0.9; p < 0.05) associated with an 11% decrease in glucose infusion rate (p < 0.05). However, the grape polyphenol group did not show these deleterious effects of fructose. Moreover, grape polyphenol supplementation showed protective effects against fructose-induced oxidative stress markers of urinary F2-isoprostanes, muscle thiobarbituric acid reactive substances (TBARS), while fructose load induced a downregulation of mitochondrial genes and decreased mitochondrial respiration in permeabilized muscle fibers [126]. In a randomized, controlled study of 38 males with at least one component of metabolic syndrome [127], the daily consumption of 20 g of wine grape pomace flour containing 10 g of dietary fiber, 822 mg of polyphenols for 16 weeks showed a significant reduction (p < 0.05) in postprandial insulin and fasting glucose levels compared with the baseline, but there were no significant differences in fasting insulin, postprandial glucose, insulin, glycosylated haemoglobin and HOMA–IR between a wine grape pomace flour group and a control group [127].
In summary, there is little persuasive evidence of an effect of mixed grape polyphenols on glucose homeostasis.
Resveratrol (3, 5, 4′-trihydroxy-trans-stilbene) is a phytoallexin naturally synthesized by plants in response to infection and injury pathogens such as bacteria or fungi. Berries, grape skins, red wine, Japanese knotweed, peanuts, and the roots of rhubarb are sources of resveratrol. Resveratrol exists in two isomeric forms. The main form is trans-resveratrol mainly found in red grape juice (3.38 mg/L) [128]. The levels of resveratrol in food vary. Its levels in red wine are higher (5.8 mg/L vs. 0.2 mg/L) than white wine as red wine is fermented with skins [129].
A randomized, double-blind, crossover study of 11 healthy, obese men showed that resveratrol supplementation of 150 mg/day for 30 days reduced glucose, insulin, HOMA–IR and leptin, decreased inflammatory markers (TNF-α, leukocyte count, alanine aminotransferase) and SBP by 5 mmHg, improved muscle mitochondrial respiration on a fatty acid-derived substrate from the vastus lateralis muscle biopsy (~30 mg), and decreased adipose tissue lipolysis and plasma fatty acid and glycerol in the postprandial state [130]. However, there was no control group or placebo period. Nineteen subjects with T2D took 5 mg trans-resveratrol daily for four weeks and showed improved insulin sensitivity as assessed by HOMA–IR, decreased glucose levels and a delayed glucose peak following a standard meal. There was no control group or placebo used. This study considered these favourable effects were attributed to decreased oxidative stress leading to more efficient insulin signalling via the Akt pathway, with an increase in the ratio of phosphorylated vs. total Akt (pAkt: Akt) ratio in platelets [131]. However, a combination of other polyphenolic compounds found in red wine may contribute to the many beneficial effects, as resveratrol is known to have low bioavailability [104].
In summary, it is hard to conclude that resveratrol has anti-diabetic effects. More clinical trials of resveratrol are required to confirm the findings described above in this article.
5.5. Red Wine
In a randomized, controlled study, red wine significantly improved insulin sensitivity, lipid profiles and endothelial function compared with other alcoholic beverages [132,133].
A study [134] comparing the acute effects of red wine, beer and vodka on oxidative stress reported that only red wine demonstrated protection against oxygen-induced oxidative stress, while all three alcoholic beverages provided similar protection against oxygen-induced (via hyperoxia) increase in arterial stiffness. Ten males randomly consumed one of four tested beverage of red wine, vodka, beer (0.32 g ethanol/kg body weight) and water (control). Every beverage was consumed once, a week apart in a cross-over design. The volunteers breathed 100% normobaric O2 between 60th and 90th min of 3-h study protocol. Plasma lipid peroxides (LOOH—biochemical marker of oxidative stress) and uric acid (UA) concentration, blood alcohol concentration (BAC) and arterial stiffness—indicated by augmentation index (AIx)—were measured before and 30, 60, 90, 120 and 180 min after beverage consumption [134]. Moreover, 2 g/day of grape seed extract (1 g of polyphenols) also improved endothelial function in subjects with high vascular risk [135].
However, overall, there is little evidence of an effect of wine polyphenols on glucose and insulin.
5.6. Pomegranate
Fresh pomegranate juice is comprised of phenolic acids, including gallic acid, chlorogenic acid, caffeic acid, ferulic acid, and coumaric acids, as well as flavonoids, such as catechin, phloridzin, and quercetin. Other flavan-3-ols such as epicatechin and epigallocatechin are present in pomegranate [136]. Various in vivo and in vitro studies have demonstrated favourable effects of pomegranate on insulin sensitivity, inhibition of a-glucosidase and reduced oxidative stress and lipid peroxidation [136,137]. Very limited human studies have focused on the effects of pomegranate juice on T2D [137]. One hundred and twenty milliliters of pomegranate juice/day in obese subjects did not modify insulin secretion and sensitivity after one month of administration in a fasted state [138]. Similarly, 50 mL of pomegranate juice/day for four weeks in subjects with T2D induced no changes in fasting glucose, insulin and HbA1c [139]. Overall, there appears to be little benefit of pomegranate juice.
5.7. Berries
In a randomized, cross-over, double-blind, placebo-controlled acute meal study of 20 healthy subjects, no significant effects were observed on plasma glucose, plasma triacylglycerol, plasma non esterified fatty acids, as well as markers of vascular inflammation, intercellular adhesion molecule-1 (sICAM-1) and vascular cell adhesion molecule-1 (sVCAM-1) after consumption of a black currant (20% juice), compared with consumption of placebo juice devoid of polyphenols and Vitamin C. Both drinks contained 50% citric acid solution, 1% aspartame and 8% acesulfame K. However, an increase in plasma insulin levels was detected [140]. Freeze-dried blueberry beverage containing 50 g of blueberry powder (equivalent to approximately 350 g fresh blueberries) was given daily to 48 individuals with metabolic syndrome for eight weeks in a randomized controlled trial. The serum glucose concentration, lipid profiles, fasting plasma hs-CRP, IL-6, adiponectin, sICAM-1 and sVCAM-1 were not changed significantly, whereas blood pressure decreased in the blueberry-supplemented group compared with the 960 mL water control group [141]. Insulin sensitivity improved more in a 22.5 g blueberry group (22.2% ± 5.8%) than in the placebo group (4.9% ± 4.5%) (p = 0.02) as measured by a high-dose hyperinsulinemic-euglycemic clamp [142]. The thirty-two obese, nondiabetic, and insulin-resistant participants of this study showed no significant changes in adiposity, energy intake, and inflammatory biomarkers.
Polyphenol-rich berries may reduce sucrose digestion and absorption leading to delayed glycaemic response [143,144]. In a randomised, controlled, cross-over study of two 3-h meal tests in healthy subjects, a berry puree (150 g) made of equal amounts (37.5 g) of bilberries, blackcurrants, cranberries and strawberries, and sweetened with 35 g sucrose lowered glucose levels at 15 (p < 0.05) and 30 min (p < 0.01) compared with the control meal including 250 mL water, 35 g sucrose, 4.5 g glucose and 5.1 g fructose which are similar in glycemic profile and amounts of available carbohydrates. The berry meal and the control meal showed the peak glucose concentration at 45 and 30 min, respectively. No significant difference in the 3-h area under the glucose response curve was observed. These findings indicated that berries rich in polyphenols delayed digestion of sucrose and/or absorption of glucose from the berry meal or slowed gastric empting [144]. In three randomised, controlled, cross-over, 2-h meal studies [143], healthy female volunteers underwent four 2-h meal tests on separate visits, at least three days apart with a mix of berries and bread strawberries, and a berry mix reduced insulin but not glucose to both wheat and rye breads [143].
A randomized, single-blind, placebo-controlled, cross-over trial in an acute-meal setting showed beneficial effects of polyphenolic- and antioxidant-rich strawberry (Fragaria) on postprandial inflammation and insulin sensitivity. Twenty-four overweight subjects consumed two isocaloric meals of high-carbohydrate, moderate-fat (HCF) accompanied by either a strawberry or a strawberry-flavoured beverage that served as a placebo. The strawberry beverage significantly attenuated the postprandial inflammatory response as measured by hsCRP and IL-6 (p < 0.05) and reduced postprandial insulin response (p < 0.05). In addition to polyphenolic compounds, Vitamin C and glutathione were also suggested to contribute to the antioxidant activity of strawberry. The test meals including strawberry and placebo beverage did not contain any soluble fibre [145]. The same team conducted an extended study [146] to investigate the effects of six weeks of strawberry consumption (~95 mg total strawberry phenols/day) on HCF meal-induced increases in glucose, insulin and indicators of inflammation and homeostasis over 6 h. Twenty-four overweight subjects consumed either a strawberry beverage or a placebo beverage for six weeks after a seven-day run-in of low/no berry consumption. Strawberry consumption attenuated postprandial plasminogen activator inhibitor-1,plasminogen activator inhibitor-1 (PAI-1) (p = 0.002), IL-1 β (p = 0.05) with moderate suppression of IL-6, but no significant changes in platelet aggregation, hsCRP, TNF-α, insulin, or glucose were observed compared with the placebo [146]. Consumption of a beverage of 50 g freeze-dried strawberries (2006 mg polyphenols) for eight weeks decreased total cholesterol and LDL-C, small LDL particles and VCAM-1, whereas waist circumference, SBP/DBP, fasting glucose, triglycerides, and HDL-C were not affected, compared with control (equivalent amounts of water) in 27 obese subjects with metabolic syndrome after eight weeks [147]. Daily consumption of two cups of a strawberry drink for four weeks in women with metabolic syndrome (each cup had 25 g of freeze-dried strawberry powder equivalent to approximately 3.5 cups or 500 g fresh strawberries) decreased total cholesterol and LDL-C levels by 5% and 6%, respectively (p < 0.05) and decreased lipid peroxidation in the form of malondialdehyde and hydroxynonenal by 14% (p < 0.01) at four weeks versus baseline. There was no control group or control period. This study suggested that phytosterol, dietary fiber (8 g/day), or polyphenolic flavonoids content of strawberries might contribute to the hypocholesterolemic effects of freeze dried strawberry powder [148].
A small study suggested that dried cranberries might lower the glycemic response for people with T2D [149], but other studies did not find glucose-lowering effects, despite antioxidant, anti-inflammatory, and HDL-raising and LDL-lowering effects of cranberry juice supplementation [150,151,152,153].
In summary, berries appear to reduce the insulin response to a glucose load and have anti-inflammatory effects. However, the results are very mixed and there is limited data.
5.8. Extra Virgin Olive Oil
In the PREDIMED study of 3541 patients with high cardiovascular risk with a 4.1-year follow-up period, a Mediterranean diet rich in extra virgin oil showed a 40% reduction (95% CI 15–57) in the risk of T2D compared with the control group [154]. An olive-rich Mediterranean diet also improved glucose metabolism and reduced body weight in subjects with T2D. Insulin resistance was measured by HOMA–IR, adiponectin/leptin and adiponectin/HOMA–R ratios after one year of follow-up, but HbA1c was not measured [155]. Moreover, the Mediterranean diet supplemented with virgin olive oil or nuts showed an anti-inflammatory effect by decreasing CRP, IL-6 and adhesion molecules, chemokines and T-lymphocytes (CD49d) and monocytes (CD 11b, CD49d, CD40) [156]. In contrast, in a study of a six-week olive oil supplementation in healthy adults, fasting glucose and HDL-C increased compared with baseline in both the low- and high-phenolic olive oil group. There was no change in fructosamine, or plasma total phenols within either group. No significance in a proteomic biomarker score for diabetes was observed with either low- or high-phenolic olive oil [157]. Phenolic components such as oleuropein and hydroxytyrosol abundant in olive leaves as well as oil may protect against diabetes and metabolic disorders [158]. Only two intervention studies of olive leaf polyphenols [159,160] have been conducted so far. Supplementation with olive leaf polyphenols (51.1 mg oleuropein and 9.7 mg hydroxytyrosol per day) for 12 weeks significantly improved insulin sensitivity and pancreatic β-cells secretory capacity after oral glucose challenge in overweight, middle-aged men at the risk of developing the metabolic syndrome [159]. Similarly, in a randomized, placebo-controlled trial in subjects with T2D, supplementation of a 500 mg olive leaf extract tablet once daily for 14 weeks was shown to significantly lower HbA1c and fasting insulin with no significant changes in postprandial insulin levels [159,160].
More clinical studies evaluating the potential effects of olive oil or leaf polyphenols on insulin sensitivity and glycaemic control should be conducted to confirm these beneficial effects.
5.9. Whole Grains, Nuts and Seeds
Whole grains such as wheat, rye, soy and flaxseed are a good source of polyphenols as are nuts such as chestnuts, hazelnuts, pecans and almonds [161]. Whole grain intake is associated with a reduced risk of T2D but the mechanism of the protection is not clear [162]. Whole grain intake is also associated with a lower body weight, lower waist circumference and lower fasting insulin in the Framingham offspring study [163]. However, interventions comparing whole grain with refined grain cereals for 12 weeks have shown no effect on insulin sensitivity or insulin release in 146 individuals with the metabolic syndrome [164] although 2- and 3-h postprandial insulin and triglyceride was lower after the whole grain intervention [165] compared with a standard test meal during the run-in period. A large study with 316 individuals compared usual diet (<30 g/day of whole grains) to 60 and 120 g/day for four months. No effects were seen on glucose, insulin, inflammatory, coagulation or endothelial markers [166]. It would appear despite the epidemiology that increasing whole grain intake with more fibre and polyphenols has little effect on metabolic risk. However, in the Predimed study, nuts reduced the incidence of T2D assessed annually by OGTT by 52% in the Reus Substudy over four years [167].
5.10. Fruits and Vegetables
Most prospective studies have shown no effect of vegetables and fruits on insulin resistance or the risk of T2D [168,169,170,171]. Only a high daily consumption of green leafy vegetables was inversely associated with T2D [169,170], while a high intake of berries was associated with a 35% lower risk of T2D (HR 0.65; 95% CI 0.49–0.88; p = 0.15) [171].
5.11. Dietary Patterns
5.11.1. Polyphenol-Rich Diets
Polyphenol-rich diets have been shown to reduce blood glucose in a 3-h OGTT by increasing early insulin secretion and improving insulin sensitivity. Eighty-six participants aged 35–70 years with a BMI of 27–35 kg/m2 at high cardiometabolic risk were randomized to one of four eight-week isoenergetic diets with a 2 × 2 factorial design [90]: (1) high in polyphenols and high in long chain n-3 polyunsaturated fatty acids (2861 mg of polyphenols/day); (2) high in polyphenols and low in long chain n-3 polyunsaturated fatty acids (2903 mg/day); (3) low in polyphenols and high in long chain n-3 polyunsaturated fatty acids from fish (363 mg/day); (4) low in polyphenols and low in long chain n-3 polyunsaturated fatty acids (365 mg/day). Polyphenols came from decaffeinated green tea and coffee, dark chocolate, blueberry jam, extra-virgin olive oil and polyphenol rich vegetables such as steamed artichokes, onions and spinach, and raw rocket. The polyphenols were 57% flavonoids (41% flavanols) and 43% phenolic acids. The high polyphenol diet decreased plasma glucose area under the curve (AUC; p = 0.038) and increased the early phase of insulin secretion (AUC 0–30 min; p = 0.048) [172].
5.11.2. Nordic Diet
Scandinavian Nordic diets are considered healthy diets as they are rich in high-fibre plant foods (such as vegetables, fruits and berries). They also include whole grains, fish, rapeseed oils, nuts and low-fat dairy but are low in salt, added sugars and saturated fats [173,174,175]. This diet appears to be related to a lower risk of T2D.
Cross-sectional studies [176] and randomized clinical trials [175,177,178] of this Nordic diet showed weight loss compared with controls. In a randomized controlled trial of 88 mildly hypercholesterolaemic subjects [175], subjects who consumed a Nordic diet for six weeks had decreased insulin (−9%; p = 0.01) and SBP by −6.6 ± 13.2 mmHg (−5%; p < 0.05) compared with the control group following their usual Western diet. However, the six-week Nordic diet caused a 4% weight loss compared with the control group (p < 0.001). After adjustment for weight change, no significant differences in DBP or triglyceride or glucose was observed although blood lipids were still improved [175]. Similarly, a weight-stable Nordic diet for 18–24 weeks [177] did not alter insulin sensitivity and blood pressure in people with metabolic syndrome. Significant changes between the groups were found in non-HDL-C, LDL-C to HDL-C and apolipoprotein B to apolipoprotein A1 ratios favouring the Nordic diet.
In contrast, a prospective study of 10-year follow-up showed no association between a healthy Nordic diet and the risk of T2D [179]. In a systematic review [173] of prospective observational and randomized intervention studies investigating the association between five food groups (red and processed meat, whole grains, berries, milk products and potatoes) commonly consumed in Nordic countries and the risk of T2D, only wholegrain was inversely associated with the risk of T2D [173]. The association between a Nordic diet and a lower risk of increased hs-CRP levels was observed in a meta-analysis of three cross-sectional Finnish studies [180]. Nevertheless, high adherence to this diet was associated with a lower risk of low HDL-C levels, especially in women [180].
5.11.3. Mediterranean Diet
There have been no interventions comparing a Mediterranean diet to a Western control diet for the prevention of T2D but there have been many interventions in people with T2D. A recent meta-analysis of nine interventions showed that a Mediterranean diet reduced HbA1c by 0.3% and also improved many other metabolic parameters [181].
6. Conclusions
In vitro and in vivo studies have shown that dietary polyphenolic compounds improved glucose homeostasis through potential multiple mechanisms of action in the intestine, liver, muscle adipocytes and pancreatic β-cells, as well as through prebiotic effects in the digestive tract. Overall, most epidemiological studies showed that dietary polyphenols were associated with a lower risk of T2D, but this association was not entirely consistent. Berries and cassia cinnamon may be promising candidates for diabetes prevention and management. There have only been limited clinical studies of dietary polyphenols in relation to glucose and insulin homeostasis and these have not been as successful as in vitro and in vivo studies. The mixed results may be related to poor statistical analyses, small sample size (many are acute studies) and no dynamic measures of glucose metabolism. Larger and better designed studies are required before any recommendations can be made.
Acknowledgments
Peter Clifton is supported by a NHMRC Principal Research Fellowship. University of South Australia. Associate Professor Jennifer Keogh is a Fellow of the South Australian Cardiovascular Research Development Program funded by the Heart Foundation and the Government of South Australia.
Author Contributions
All authors conceived of the manuscript structure and contributed to the writing and editing.
Conflict of Interest
The authors have no conflicts of interest related to this manuscript.
References
- Nathan, D.M. Long-term complications of diabetes mellitus. N. Engl. J. Med. 1993, 328, 1676–1685. [Google Scholar] [CrossRef] [PubMed]
- Guariguata, L.; Whiting, D.R.; Hambleton, I.; Beagley, J.; Linnenkamp, U.; Shaw, J.E. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res. Clin. Pract. 2014, 103, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, X.; Brown, J.; Vistisen, D.; Sicree, R.; Shaw, J.; Nichols, G. Global healthcare expenditure on diabetes for 2010 and 2030. Diabetes Res. Clin. Pract. 2010, 87, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Pfeiffer, A.F. Foods for the prevention of diabetes: How do they work? Diabetes Metab. Res. Rev. 2012, 28, 25–49. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.B.; Hogger, P. Dietary polyphenols and type 2 diabetes: Current insights and future perspectives. Curr. Med. Chem. 2015, 22, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Hanhineva, K.; Torronen, R.; Bondia-Pons, I.; Pekkinen, J.; Kolehmainen, M.; Mykkanen, H.; Poutanen, K. Impact of dietary polyphenols on carbohydrate metabolism. Int. J. Mol. Sci. 2010, 11, 1365–1402. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G. Possible effects of dietary polyphenols on sugar absorption and digestion. Mol. Nutr. Food Res. 2013, 57, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [PubMed]
- Sun, Q.; Wedick, N.M.; Tworoger, S.S.; Pan, A.; Townsend, M.K.; Cassidy, A.; Franke, A.A.; Rimm, E.B.; Hu, F.B.; van Dam, R.M. Urinary excretion of select dietary polyphenol metabolites is associated with a lower risk of type 2 diabetes in proximate but not remote follow-up in a prospective investigation in 2 cohorts of US women. J. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Manson, J.E.; Buring, J.E.; Sesso, H.D.; Liu, S. Associations of dietary flavonoids with risk of type 2 diabetes, and markers of insulin resistance and systemic inflammation in women: A prospective study and cross-sectional analysis. J. Am. Coll. Nutr. 2005, 24, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Knekt, P.; Kumpulainen, J.; Jarvinen, R.; Rissanen, H.; Heliovaara, M.; Reunanen, A.; Hakulinen, T.; Aromaa, A. Flavonoid intake and risk of chronic diseases. Am. J. Clin. Nutr. 2002, 76, 560–568. [Google Scholar] [PubMed]
- Wedick, N.M.; Pan, A.; Cassidy, A.; Rimm, E.B.; Sampson, L.; Rosner, B.; Willett, W.; Hu, F.B.; Sun, Q.; van Dam, R.M. Dietary flavonoid intakes and risk of type 2 diabetes in US men and women. Am. J. Clin. Nutr. 2012, 95, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Nettleton, J.A.; Harnack, L.J.; Scrafford, C.G.; Mink, P.J.; Barraj, L.M.; Jacobs, D.R., Jr. Dietary flavonoids and flavonoid-rich foods are not associated with risk of type 2 diabetes in postmenopausal women. J. Nutr. 2006, 136, 3039–3045. [Google Scholar] [PubMed]
- Ding, M.; Bhupathiraju, S.N.; Chen, M.; van Dam, R.M.; Hu, F.B. Caffeinated and decaffeinated coffee consumption and risk of type 2 diabetes: A systematic review and a dose-response meta-analysis. Diabetes Care 2014, 37, 569–586. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Zhang, D.; Jiang, W. Coffee and caffeine intake and incidence of type 2 diabetes mellitus: A meta-analysis of prospective studies. Eur. J. Nutr. 2014, 53, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, R.M.; Hu, F.B. Coffee consumption and risk of type 2 diabetes: A systematic review. JAMA 2005, 294, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.A.; Parker, E.D.; Folsom, A.R. Coffee consumption and risk of type 2 diabetes mellitus: An 11-year prospective study of 28,812 postmenopausal women. Arch. Intern. Med. 2006, 166, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, R.M.; Feskens, E.J. Coffee consumption and risk of type 2 diabetes mellitus. Lancet 2002, 360, 1477–1478. [Google Scholar] [CrossRef]
- Iso, H.; Date, C.; Wakai, K.; Fukui, M.; Tamakoshi, A. The relationship between green tea and total caffeine intake and risk for self-reported type 2 diabetes among japanese adults. Ann. Int. Med. 2006, 144, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Van Dieren, S.; Uiterwaal, C.S.; van der Schouw, Y.T.; van der, A.D.; Boer, J.M.; Spijkerman, A.; Grobbee, D.E.; Beulens, J.W. Coffee and tea consumption and risk of type 2 diabetes. Diabetologia 2009, 52, 2561–2569. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Mao, Q.X.; Xu, H.X.; Ma, X.; Zeng, C.Y. Tea consumption and risk of type 2 diabetes mellitus: A systematic review and meta-analysis update. BMJ Open 2014, 4, e005632. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Wang, W.Y.; Fan, W.Y.; Deng, Q.; Wang, X. Tea consumption and risk of type 2 diabetes: A dose-response meta-analysis of cohort studies. Br. J. Nutr. 2014, 111, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Huxley, R.; Lee, C.M.; Barzi, F.; Timmermeister, L.; Czernichow, S.; Perkovic, V.; Grobbee, D.E.; Batty, D.; Woodward, M. Coffee, decaffeinated coffee, and tea consumption in relation to incident type 2 diabetes mellitus: A systematic review with meta-analysis. Arch. Int. Med. 2009, 169, 2053–2063. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, R.M.; Dekker, J.M.; Nijpels, G.; Stehouwer, C.D.; Bouter, L.M.; Heine, R.J. Coffee consumption and incidence of impaired fasting glucose, impaired glucose tolerance, and type 2 diabetes: The hoorn study. Diabetologia 2004, 47, 2152–2159. [Google Scholar] [CrossRef] [PubMed]
- Yarmolinsky, J.; Mueller, N.T.; Duncan, B.B.; Molina, M.B.C.; Goulart, A.C.; Schmidt, M.I. Coffee consumption, newly diagnosed diabetes, and other alterations in glucose homeostasis: A cross-sectional analysis of the longitudinal study of adult health (ELSA-Brasil). PLoS ONE 2015, 10, e0126469. [Google Scholar] [CrossRef] [PubMed]
- Pham, N.M.; Nanri, A.; Kochi, T.; Kuwahara, K.; Tsuruoka, H.; Kurotani, K.; Akter, S.; Kabe, I.; Sato, M.; Hayabuchi, H.; et al. Coffee and green tea consumption is associated with insulin resistance in Japanese adults. Metabolism 2014, 63, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Agardh, E.E.; Carlsson, S.; Ahlbom, A.; Efendic, S.; Grill, V.; Hammar, N.; Hilding, A.; Ostenson, C.G. Coffee consumption, type 2 diabetes and impaired glucose tolerance in Swedish men and women. J. Int. Med. 2004, 255, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Arnlov, J.; Vessby, B.; Riserus, U. Coffee consumption and insulin sensitivity. JAMA 2004, 291, 1199–1201. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, K.; Yatsuya, H.; Muramatsu, T.; Toyoshima, H.; Murohara, T.; Tamakoshi, K. Association of coffee consumption with serum adiponectin, leptin, inflammation and metabolic markers in Japanese workers: A cross-sectional study. Nutr. Diabetes 2012, 2, e33. [Google Scholar] [CrossRef] [PubMed]
- Hamer, M.; Witte, D.R.; Mosdol, A.; Marmot, M.G.; Brunner, E.J. Prospective study of coffee and tea consumption in relation to risk of type 2 diabetes mellitus among men and women: The whitehall II study. Br. J. Nutr. 2008, 100, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Koh, L.W.; Wong, L.L.; Loo, Y.Y.; Kasapis, S.; Huang, D. Evaluation of different teas against starch digestibility by mammalian glycosidases. J. Agric. Food Chem. 2010, 58, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Roder, P.V.; Geillinger, K.E.; Zietek, T.S.; Thorens, B.; Koepsell, H.; Daniel, H. The role of sglt1 and GLUT2 in intestinal glucose transport and sensing. PLoS ONE 2014, 9, e89977. [Google Scholar] [CrossRef] [PubMed]
- McDougall, G.J.; Shpiro, F.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Different polyphenolic components of soft fruits inhibit alpha-amylase and alpha-glucosidase. J. Agric. Food Chem. 2005, 53, 2760–2766. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.I.; Apostolidis, E.; Kim, Y.C.; Shetty, K. Health benefits of traditional corn, beans, and pumpkin: In vitro studies for hyperglycemia and hypertension management. J. Med. Food 2007, 10, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.I.; Apostolidis, E.; Shetty, K. In vitro studies of eggplant (solanum melongena) phenolics as inhibitors of key enzymes relevant for type 2 diabetes and hypertension. Bioresour. Technol. 2008, 99, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Sang, W.; Zhou, M.; Ren, G. Antioxidant and α-glucosidase inhibitory activity of colored grains in China. J. Agric. Food Chem. 2009, 58, 770–774. [Google Scholar] [CrossRef] [PubMed]
- Ademiluyi, A.O.; Oboh, G. Phenolic-rich extracts from selected tropical underutilized legumes inhibit alpha-amylase, alpha-glucosidase, and angiotensin I converting enzyme in vitro. J. Basic Clin. Physiol. Pharmacol. 2012, 23, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y.; Honda, M. The inhibition of α-amylase by tea polyphenols. Agric. Biol. Chem. 1990, 54, 1939–1945. [Google Scholar] [CrossRef]
- Kwon, Y.I.; Apostolidis, E.; Shetty, K. Inhibitory potential of wine and tea against α-amylase and α-glucosidase for management of hyperglycemia linked to type 2 diabetes. J. Food Biochem. 2008, 32, 15–31. [Google Scholar] [CrossRef]
- Hanamura, T.; Mayama, C.; Aoki, H.; Hirayama, Y.; Shimizu, M. Antihyperglycemic effect of polyphenols from Acerola (malpighia emarginata DC.) fruit. Biosci. Biotechnol. Biochem. 2006, 70, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Welsch, C.A.; Lachance, P.A.; Wasserman, B.P. Dietary phenolic compounds: Inhibition of Na+-dependent D-glucose uptake in rat intestinal brush border membrane vesicles. J. Nutr. 1989, 119, 1698–1704. [Google Scholar] [PubMed]
- Cermak, R.; Landgraf, S.; Wolffram, S. Quercetin glucosides inhibit glucose uptake into brush-border-membrane vesicles of porcine jejunum. Br. J. Nutr. 2004, 91, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Johnston, K.; Sharp, P.; Clifford, M.; Morgan, L. Dietary polyphenols decrease glucose uptake by human intestinal caco-2 cells. Febs Lett. 2005, 579, 1653–1657. [Google Scholar] [CrossRef] [PubMed]
- Manzano, S.; Williamson, G. Polyphenols and phenolic acids from strawberry and apple decrease glucose uptake and transport by human intestinal Caco-2 cells. Mol. Nutr. Food Res. 2010, 54, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Ueda-Wakagi, M.; Mukai, R.; Fuse, N.; Mizushina, Y.; Ashida, H. 3-O-acyl-epicatechins increase glucose uptake activity and GLUT4 translocation through activation of PI3K signaling in skeletal muscle cells. Int. J. Mol. Sci. 2015, 16, 16288–16299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; Li, Q.; Liang, J.; Dai, X.Q.; Ding, Y.; Wang, J.B.; Li, Y. Epigallocatechin-3-O-gallate (EGCG) protects the insulin sensitivity in rat l6 muscle cells exposed to dexamethasone condition. Phytomedicine 2010, 17, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Pinent, M.; Blay, M.; Blade, M.; Salvado, M.; Arola, L.; Ardevol, A. Grape seed-derived procyanidins have an antihyperglycemic effect in streptozotocin-induced diabetic rats and insulinomimetic activity in insulin-sensitive cell lines. Endocrinology 2004, 145, 4985–4990. [Google Scholar] [CrossRef] [PubMed]
- Montagut, G.; Onnockx, S.; Vaqué, M.; Bladé, C.; Blay, M.; Fernández-Larrea, J.; Pujadas, G.; Salvadó, M.J.; Arola, L.; Pirson, I. Oligomers of grape-seed procyanidin extract activate the insulin receptor and key targets of the insulin signaling pathway differently from insulin. J. Nutr. Biochem. 2010, 21, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Cummings, E.; Hundal, H.S.; Wackerhage, H.; Hope, M.; Belle, M.; Adeghate, E.; Singh, J. Momordica charantia fruit juice stimulates glucose and amino acid uptakes in L6 myotubes. Mol. Cell. Biochem. 2004, 261, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Vuong, T.; Martineau, L.C.; Ramassamy, C.; Matar, C.; Haddad, P.S. Fermented Canadian lowbush blueberry juice stimulates glucose uptake and AMP-activated protein kinase in insulin-sensitive cultured muscle cells and adipocytes. Can. J. Physiol. Pharmacol. 2007, 85, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Purintrapiban, J.; Suttajit, M.; Forsberg, N.E. Differential activation of glucose transport in cultured muscle cells by polyphenolic compounds from Canna indica L. Root. Biol. Pharm. Bull. 2006, 29, 1995–1998. [Google Scholar] [CrossRef] [PubMed]
- Kurimoto, Y.; Shibayama, Y.; Inoue, S.; Soga, M.; Takikawa, M.; Ito, C.; Nanba, F.; Yoshida, T.; Yamashita, Y.; Ashida, H.; et al. Black soybean seed coat extract ameliorates hyperglycemia and insulin sensitivity via the activation of AMP-activated protein kinase in diabetic mice. J. Agric. Food Chem. 2013, 61, 5558–5564. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Polansky, M.M.; Anderson, R.A. Cinnamon extract and polyphenols affect the expression of tristetraprolin, insulin receptor, and glucose transporter 4 in mouse 3T3-L1 adipocytes. Arch. Biochem. Biophys. 2007, 459, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Nagasaki, M.; Ren, M.; Bajotto, G.; Oshida, Y.; Sato, Y. Cinnamon extract (traditional herb) potentiates in vivo insulin-regulated glucose utilization via enhancing insulin signaling in rats. Diabetes Res. Clin. Pract. 2003, 62, 139–148. [Google Scholar] [CrossRef]
- Qin, B.; Nagasaki, M.; Ren, M.; Bajotto, G.; Oshida, Y.; Sato, Y. Cinnamon extract prevents the insulin resistance induced by a high-fructose diet. Horm. Metab. Res. 2004, 36, 119–125. [Google Scholar] [PubMed]
- Cardona, F.; Andres-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuno, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Anhê, F.F.; Desjardins, Y.; Pilon, G.; Dudonné, S.; Genovese, M.I.; Lajolo, F.M.; Marette, A. Polyphenols and type 2 diabetes: A prospective review. PharmaNutrition 2013, 1, 105–114. [Google Scholar] [CrossRef]
- Cani, P.D.; Possemiers, S.; van de Wiele, T.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut 2009, 58, 1091–1103. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Neyrinck, A.M.; Fava, F.; Knauf, C.; Burcelin, R.G.; Tuohy, K.M.; Gibson, G.; Delzenne, N.M. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia 2007, 50, 2374–2383. [Google Scholar] [PubMed]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Prebiotic evaluation of cocoa-derived flavanols in healthy humans by using a randomized, controlled, double-blind, crossover intervention study. Am. J. Clin. Nutr. 2011, 93, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Vendrame, S.; Guglielmetti, S.; Riso, P.; Arioli, S.; Klimis-Zacas, D.; Porrini, M. Six-week consumption of a wild blueberry powder drink increases bifidobacteria in the human gut. J. Agric. Food Chem. 2011, 59, 12815–12820. [Google Scholar] [CrossRef] [PubMed]
- Yamakoshi, J.; Tokutake, S.; Kikuchi, M.; Kubota, Y.; Konishi, H.; Mitsuoka, T. Effect of proanthocyanidin-rich extract from grape seeds on human fecal flora and fecal odor. Microb. Ecol. Health Dis. 2001, 13, 25–31. [Google Scholar] [CrossRef]
- Jin, J.S.; Touyama, M.; Hisada, T.; Benno, Y. Effects of green tea consumption on human fecal microbiota with special reference to bifidobacterium species. Microbiol. Immunol. 2012, 56, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Queipo-Ortuno, M.I.; Boto-Ordonez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Cardona Diaz, F.; Andres-Lacueva, C.; Tinahones, F.J. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am. J. Clin. Nutr. 2012, 95, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Cuervo, A.; Hevia, A.; Lopez, P.; Suarez, A.; Sanchez, B.; Margolles, A.; Gonzalez, S. Association of polyphenols from oranges and apples with specific intestinal microorganisms in systemic lupus erythematosus patients. Nutrients 2015, 7, 1301–1317. [Google Scholar] [CrossRef] [PubMed]
- Henquin, J.C. Triggering and amplifying pathways of regulation of insulin secretion by glucose. Diabetes 2000, 49, 1751–1760. [Google Scholar] [CrossRef] [PubMed]
- Cai, E.P.; Lin, J.K. Epigallocatechin gallate (EGCG) and rutin suppress the glucotoxicity through activating IRS2 and AMPK signaling in rat pancreatic beta cells. J. Agric. Food Chem. 2009, 57, 9817–9827. [Google Scholar] [CrossRef] [PubMed]
- Rutter, G.A.; da Silva, X.G.; Leclerc, I. Roles of 5′-AMP-activated protein kinase (AMPK) in mammalian glucose homoeostasis. Biochem. J. 2003, 375, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Kwon, K.B.; Song, M.Y.; Han, M.J.; Lee, J.H.; Lee, Y.R.; Lee, J.H.; Ryu, D.G.; Park, B.H.; Park, J.W. Flavonoids protect against cytokine-induced pancreatic beta-cell damage through suppression of nuclear factor κB activation. Pancreas 2007, 35, e1–e9. [Google Scholar] [CrossRef] [PubMed]
- Coskun, O.; Kanter, M.; Korkmaz, A.; Oter, S. Quercetin, a flavonoid antioxidant, prevents and protects streptozotocin-induced oxidative stress and beta-cell damage in rat pancreas. Pharmacol. Res. 2005, 51, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Hii, C.S.; Howell, S.L. Effects of flavonoids on insulin secretion and 45Ca2+ handling in rat islets of Langerhans. J. Endocrinol. 1985, 107, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Waltner-Law, M.E.; Wang, X.L.; Law, B.K.; Hall, R.K.; Nawano, M.; Granner, D.K. Epigallocatechin gallate, a constituent of green tea, represses hepatic glucose production. J. Biol. Chem. 2002, 277, 34933–34940. [Google Scholar] [CrossRef] [PubMed]
- Collins, Q.F.; Liu, H.Y.; Pi, J.; Liu, Z.; Quon, M.J.; Cao, W. Epigallocatechin-3-gallate (EGCG), a green tea polyphenol, suppresses hepatic gluconeogenesis through 5′-AMP-activated protein kinase. J. Biol. Chem. 2007, 282, 30143–30149. [Google Scholar] [CrossRef] [PubMed]
- Wolfram, S.; Raederstorff, D.; Preller, M.; Wang, Y.; Teixeira, S.R.; Riegger, C.; Weber, P. Epigallocatechin gallate supplementation alleviates diabetes in rodents. J. Nutr. 2006, 136, 2512–2518. [Google Scholar] [PubMed]
- Choi, M.S.; Jung, U.J.; Yeo, J.; Kim, M.J.; Lee, M.K. Genistein and daidzein prevent diabetes onset by elevating insulin level and altering hepatic gluconeogenic and lipogenic enzyme activities in non-obese diabetic (NOD) mice. Diabetes Metab. Res Rev 2008, 24, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Lee, M.K.; Park, Y.B.; Kang, M.A.; Choi, M.S. Effect of citrus flavonoids on lipid metabolism and glucose-regulating enzyme mrna levels in type-2 diabetic mice. Int. J. Biochem. Cell Biol. 2006, 38, 1134–1145. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.L.; Chi, C.W.; Liu, T.Y. Areca nut procyanidins ameliorate streptozocin-induced hyperglycemia by regulating gluconeogenesis. Food Chem. Toxicol. 2013, 55, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Xia, M.; Zou, T.; Ling, W.; Zhong, R.; Zhang, W. Cyanidin 3-glucoside attenuates obesity-associated insulin resistance and hepatic steatosis in high-fat diet-fed and db/db mice via the transcription factor FoxO1. J. Nutr. Biochem. 2012, 23, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Keane, K.N.; Cruzat, V.F.; Carlessi, R.; de Bittencourt, P.I., Jr.; Newsholme, P. Molecular events linking oxidative stress and inflammation to insulin resistance and beta-cell dysfunction. Oxid. Med. Cell. Longev. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Terra, X.; Montagut, G.; Bustos, M.; Llopiz, N.; Ardevol, A.; Blade, C.; Fernandez-Larrea, J.; Pujadas, G.; Salvado, J.; Arola, L.; et al. Grape-seed procyanidins prevent low-grade inflammation by modulating cytokine expression in rats fed a high-fat diet. J. Nutr. Biochem. 2009, 20, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Kanamoto, Y.; Yamashita, Y.; Nanba, F.; Yoshida, T.; Tsuda, T.; Fukuda, I.; Nakamura-Tsuruta, S.; Ashida, H. A black soybean seed coat extract prevents obesity and glucose intolerance by up-regulating uncoupling proteins and down-regulating inflammatory cytokines in high-fat diet-fed mice. J. Agric. Food Chem. 2011, 59, 8985–8993. [Google Scholar] [CrossRef] [PubMed]
- Terra, X.; Pallares, V.; Ardevol, A.; Blade, C.; Fernandez-Larrea, J.; Pujadas, G.; Salvado, J.; Arola, L.; Blay, M. Modulatory effect of grape-seed procyanidins on local and systemic inflammation in diet-induced obesity rats. J. Nutr. Biochem. 2011, 22, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Yokozawa, T.; Kim, H.J.; Cho, E.J. Gravinol ameliorates high-fructose-induced metabolic syndrome through regulation of lipid metabolism and proinflammatory state in rats. J. Agric. Food Chem. 2008, 56, 5026–5032. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.A.; Cho, E.J.; Yokozawa, T. Effects of proanthocyanidin preparations on hyperlipidemia and other biomarkers in mouse model of type 2 diabetes. J. Agric. Food Chem. 2008, 56, 7781–7789. [Google Scholar] [CrossRef] [PubMed]
- DeFuria, J.; Bennett, G.; Strissel, K.J.; Perfield, J.W., II; Milbury, P.E.; Greenberg, A.S.; Obin, M.S. Dietary blueberry attenuates whole-body insulin resistance in high fat-fed mice by reducing adipocyte death and its inflammatory sequelae. J. Nutr. 2009, 139, 1510–1516. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.K.; Soileau, J.L.; Ribnicky, D.; Wang, Z.Q.; Raskin, I.; Poulev, A.; Majewski, M.; Cefalu, W.T.; Gettys, T.W. Quercetin transiently increases energy expenditure but persistently decreases circulating markers of inflammation in C57BL/6J mice fed a high-fat diet. Metabolism 2008, 57, S39–S46. [Google Scholar] [CrossRef] [PubMed]
- Rivera, L.; Moron, R.; Sanchez, M.; Zarzuelo, A.; Galisteo, M. Quercetin ameliorates metabolic syndrome and improves the inflammatory status in obese Zucker rats. Obesity 2008, 16, 2081–2087. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tian, J.; Jiang, J.; Li, L.; Ying, X.; Tian, H.; Nie, M. Effects of green tea or green tea extract on insulin sensitivity and glycaemic control in populations at risk of type 2 diabetes mellitus: A systematic review and meta-analysis of randomised controlled trials. J. Hum. Nutr. Diet. 2014, 27, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhou, R.; Wang, B.; Chen, K.; Shi, L.Y.; Zhu, J.D.; Mi, M.T. Effect of green tea on glucose control and insulin sensitivity: A meta-analysis of 17 randomized controlled trials. Am. J. Clin. Nutr. 2013, 98, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.X.; Xu, Y.L.; Li, S.H.; Hui, R.; Wu, Y.J.; Huang, X.H. Effects of green tea catechins with or without caffeine on glycemic control in adults: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2013, 97, 750–762. [Google Scholar] [CrossRef] [PubMed]
- Johnston, K.L.; Clifford, M.N.; Morgan, L.M. Coffee acutely modifies gastrointestinal hormone secretion and glucose tolerance in humans: Glycemic effects of chlorogenic acid and caffeine. Am. J. Clin. Nutr. 2003, 78, 728–733. [Google Scholar] [PubMed]
- Hooper, L.; Kay, C.; Abdelhamid, A.; Kroon, P.A.; Cohn, J.S.; Rimm, E.B.; Cassidy, A. Effects of chocolate, cocoa, and flavan-3-ols on cardiovascular health: A systematic review and meta-analysis of randomized trials. Am. J. Clin. Nutr. 2012, 95, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Grassi, D.; Lippi, C.; Necozione, S.; Desideri, G.; Ferri, C. Short-term administration of dark chocolate is followed by a significant increase in insulin sensitivity and a decrease in blood pressure in healthy persons. Am. J. Clin. Nutr. 2005, 81, 611–614. [Google Scholar] [PubMed]
- Grassi, D.; Necozione, S.; Lippi, C.; Croce, G.; Valeri, L.; Pasqualetti, P.; Desideri, G.; Blumberg, J.B.; Ferri, C. Cocoa reduces blood pressure and insulin resistance and improves endothelium-dependent vasodilation in hypertensives. Hypertension 2005, 46, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Grassi, D.; Desideri, G.; Necozione, S.; Lippi, C.; Casale, R.; Properzi, G.; Blumberg, J.B.; Ferri, C. Blood pressure is reduced and insulin sensitivity increased in glucose-intolerant, hypertensive subjects after 15 days of consuming high-polyphenol dark chocolate. J. Nutr. 2008, 138, 1671–1676. [Google Scholar] [PubMed]
- Almoosawi, S.; Tsang, C.; Ostertag, L.M.; Fyfe, L.; al-Dujaili, E.A. Differential effect of polyphenol-rich dark chocolate on biomarkers of glucose metabolism and cardiovascular risk factors in healthy, overweight and obese subjects: A randomized clinical trial. Food Funct. 2012, 3, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Davison, K.; Coates, A.M.; Buckley, J.D.; Howe, P.R. Effect of cocoa flavanols and exercise on cardiometabolic risk factors in overweight and obese subjects. Int. J. Obes. 2008, 32, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Muniyappa, R.; Hall, G.; Kolodziej, T.L.; Karne, R.J.; Crandon, S.K.; Quon, M.J. Cocoa consumption for 2 weeks enhances insulin-mediated vasodilatation without improving blood pressure or insulin resistance in essential hypertension. Am. J. Clin. Nutr. 2008, 88, 1685–1696. [Google Scholar] [CrossRef] [PubMed]
- Rostami, A.; Khalili, M.; Haghighat, N.; Eghtesadi, S.; Shidfar, F.; Heidari, I.; Ebrahimpour-Koujan, S.; Eghtesadi, M. High-cocoa polyphenol-rich chocolate improves blood pressure in patients with diabetes and hypertension. ARYA Atheroscler. 2015, 11, 21–29. [Google Scholar] [PubMed]
- Mellor, D.D.; Madden, L.A.; Smith, K.A.; Kilpatrick, E.S.; Atkin, S.L. High-polyphenol chocolate reduces endothelial dysfunction and oxidative stress during acute transient hyperglycaemia in type 2 diabetes: A pilot randomized controlled trial. Diabetes Med. 2013, 30, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Betts, N.M.; Leyva, M.J.; Fu, D.; Aston, C.E.; Lyons, T.J. Acute cocoa supplementation increases postprandial hdl cholesterol and insulin in obese adults with type 2 diabetes after consumption of a high-fat breakfast. J. Nutr. 2015, 145, 2325–2332. [Google Scholar] [CrossRef] [PubMed]
- Sikand, G.; Kris-Etherton, P.; Boulos, N.M. Impact of functional foods on prevention of cardiovascular disease and diabetes. Curr. Cardiol. Rep. 2015, 17, 39. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Panickar, K.S.; Anderson, R.A. Cinnamon: Potential role in the prevention of insulin resistance, metabolic syndrome, and type 2 diabetes. J. Diabetes Sci. Technol. 2010, 4, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Sheng, H.; Wu, J.; Cheng, Y.; Zhu, J.; Chen, Y. Cinnamon extract improves fasting blood glucose and glycosylated hemoglobin level in Chinese patients with type 2 diabetes. Nutr. Res. 2012, 32, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Akilen, R.; Tsiami, A.; Devendra, D.; Robinson, N. Glycated haemoglobin and blood pressure-lowering effect of cinnamon in multi-ethnic type 2 diabetic patients in the UK: A randomized, placebo-controlled, double-blind clinical trial. Diabetes Med. 2010, 27, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Crawford, P. Effectiveness of cinnamon for lowering hemoglobin A1C in patients with type 2 diabetes: A randomized, controlled trial. J. Am. Board Fam. Med. 2009, 22, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Roussel, A.M.; Hininger, I.; Benaraba, R.; Ziegenfuss, T.N.; Anderson, R.A. Antioxidant effects of a cinnamon extract in people with impaired fasting glucose that are overweight or obese. J. Am. Coll. Nutr. 2009, 28, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.G.; Anderson, R.A.; Graham, G.M., III; Chu, M.C.; Sauer, M.V.; Guarnaccia, M.M.; Lobo, R.A. The effect of cinnamon extract on insulin resistance parameters in polycystic ovary syndrome: A pilot study. Fertil. Steril. 2007, 88, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.P.; Blannin, A.K. Effects of short-term cinnamon ingestion on in vivo glucose tolerance. Diabetes Obes. Metab. 2007, 9, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.; Blannin, A. Changes in glucose tolerance and insulin sensitivity following 2 weeks of daily cinnamon ingestion in healthy humans. Eur. J. Appl. Physiol. 2009, 105, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Hlebowicz, J.; Darwiche, G.; Bjorgell, O.; Almer, L.O. Effect of cinnamon on postprandial blood glucose, gastric emptying, and satiety in healthy subjects. Am. J. Clin. Nutr. 2007, 85, 1552–1556. [Google Scholar] [PubMed]
- Hlebowicz, J.; Hlebowicz, A.; Lindstedt, S.; Bjorgell, O.; Hoglund, P.; Holst, J.J.; Darwiche, G.; Almer, L.O. Effects of 1 and 3 g cinnamon on gastric emptying, satiety, and postprandial blood glucose, insulin, glucose-dependent insulinotropic polypeptide, glucagon-like peptide 1, and ghrelin concentrations in healthy subjects. Am. J. Clin. Nutr. 2009, 89, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Ziegenfuss, T.N.; Hofheins, J.E.; Mendel, R.W.; Landis, J.; Anderson, R.A. Effects of a water-soluble cinnamon extract on body composition and features of the metabolic syndrome in pre-diabetic men and women. J. Int. Soc. Sports Nutr. 2006, 3, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Safdar, M.; Ali Khan, M.M.; Khattak, K.N.; Anderson, R.A. Cinnamon improves glucose and lipids of people with type 2 diabetes. Diabetes Care 2003, 26, 3215–3218. [Google Scholar] [CrossRef] [PubMed]
- Mang, B.; Wolters, M.; Schmitt, B.; Kelb, K.; Lichtinghagen, R.; Stichtenoth, D.O.; Hahn, A. Effects of a cinnamon extract on plasma glucose, hba, and serum lipids in diabetes mellitus type 2. Eur. J. Clin. Investig. 2006, 36, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Blevins, S.M.; Leyva, M.J.; Brown, J.; Wright, J.; Scofield, R.H.; Aston, C.E. Effect of cinnamon on glucose and lipid levels in non insulin-dependent type 2 diabetes. Diabetes Care 2007, 30, 2236–2237. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Larson-Meyer, D.E.; Liebman, M. Effect of cinnamon and turmeric on urinary oxalate excretion, plasma lipids, and plasma glucose in healthy subjects. Am. J. Clin. Nutr. 2008, 87, 1262–1267. [Google Scholar] [PubMed]
- Vanschoonbeek, K.; Thomassen, B.J.; Senden, J.M.; Wodzig, W.K.; van Loon, L.J. Cinnamon supplementation does not improve glycemic control in postmenopausal type 2 diabetes patients. J. Nutr. 2006, 136, 977–980. [Google Scholar] [PubMed]
- Suppapitiporn, S.; Kanpaksi, N.; Suppapitiporn, S. The effect of cinnamon cassia powder in type 2 diabetes mellitus. J. Med. Assoc. Thail. 2006, 89, S200–S205. [Google Scholar]
- Altschuler, J.A.; Casella, S.J.; MacKenzie, T.A.; Curtis, K.M. The effect of cinnamon on A1C among adolescents with type 1 diabetes. Diabetes Care 2007, 30, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.A.; Yokoyama, W. Cinnamon intake lowers fasting blood glucose: Meta-analysis. J. Med. Food 2011, 14, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.W.; Schwartzman, E.; Baker, W.L.; Coleman, C.I.; Phung, O.J. Cinnamon use in type 2 diabetes: An updated systematic review and meta-analysis. Ann. Fam. Med. 2013, 11, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Kar, P.; Laight, D.; Rooprai, H.K.; Shaw, K.M.; Cummings, M. Effects of grape seed extract in type 2 diabetic subjects at high cardiovascular risk: A double blind randomized placebo controlled trial examining metabolic markers, vascular tone, inflammation, oxidative stress and insulin sensitivity. Diabet. Med. 2009, 26, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Hokayem, M.; Blond, E.; Vidal, H.; Lambert, K.; Meugnier, E.; Feillet-Coudray, C.; Coudray, C.; Pesenti, S.; Luyron, C.; Lambert-Porcheron, S.; et al. Grape polyphenols prevent fructose-induced oxidative stress and insulin resistance in first-degree relatives of type 2 diabetic patients. Diabetes Care 2013, 36, 1454–1461. [Google Scholar] [CrossRef] [PubMed]
- Urquiaga, I.; D’Acuna, S.; Perez, D.; Dicenta, S.; Echeverria, G.; Rigotti, A.; Leighton, F. Wine grape pomace flour improves blood pressure, fasting glucose and protein damage in humans: A randomized controlled trial. Biol. Res. 2015, 48, 49. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Moustaid-Moussa, N.; Chen, L.; Mo, H.; Shastri, A.; Su, R.; Bapat, P.; Kwun, I.; Shen, C.L. Novel insights of dietary polyphenols and obesity. J. Nutr. Biochem. 2014, 25, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Creasy, L.; Kester, A.; Zeece, M. Capillary electrophoretic determination of resveratrol in wines. J. Agric. Food Chem. 1999, 47, 3223–3227. [Google Scholar] [CrossRef] [PubMed]
- Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R.H.; van de Weijer, T.; Goossens, G.H.; Hoeks, J.; van der Krieken, S.; Ryu, D.; Kersten, S.; et al. Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab. 2011, 14, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Brasnyo, P.; Molnar, G.A.; Mohas, M.; Marko, L.; Laczy, B.; Cseh, J.; Mikolas, E.; Szijarto, I.A.; Merei, A.; Halmai, R.; et al. Resveratrol improves insulin sensitivity, reduces oxidative stress and activates the Akt pathway in type 2 diabetic patients. Br. J. Nutr. 2011, 106, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Chiva-Blanch, G.; Urpi-Sarda, M.; Ros, E.; Valderas-Martinez, P.; Casas, R.; Arranz, S.; Guillen, M.; Lamuela-Raventos, R.M.; Llorach, R.; Andres-Lacueva, C.; et al. Effects of red wine polyphenols and alcohol on glucose metabolism and the lipid profile: A randomized clinical trial. Clin. Nutr. 2013, 32, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Chiva-Blanch, G.; Urpi-Sarda, M.; Ros, E.; Arranz, S.; Valderas-Martinez, P.; Casas, R.; Sacanella, E.; Llorach, R.; Lamuela-Raventos, R.M.; Andres-Lacueva, C.; et al. Dealcoholized red wine decreases systolic and diastolic blood pressure and increases plasma nitric oxide: Short communication. Circ. Res. 2012, 111, 1065–1068. [Google Scholar] [CrossRef] [PubMed]
- Krnic, M.; Modun, D.; Budimir, D.; Gunjaca, G.; Jajic, I.; Vukovic, J.; Salamunic, I.; Sutlovic, D.; Kozina, B.; Boban, M. Comparison of acute effects of red wine, beer and vodka against hyperoxia-induced oxidative stress and increase in arterial stiffness in healthy humans. Atherosclerosis 2011, 218, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Clifton, P.M. Effect of grape seed extract and quercetin on cardiovascular and endothelial parameters in high-risk subjects. J. Biomed. Biotechnol. 2004, 2004, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Medjakovic, S.; Jungbauer, A. Pomegranate: A fruit that ameliorates metabolic syndrome. Food Funct. 2013, 4, 19–39. [Google Scholar] [CrossRef] [PubMed]
- Banihani, S.; Swedan, S.; Alguraan, Z. Pomegranate and type 2 diabetes. Nutr. Res. 2013, 33, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ortiz, M.; Martinez-Abundis, E.; Espinel-Bermudez, M.C.; Perez-Rubio, K.G. Effect of pomegranate juice on insulin secretion and sensitivity in patients with obesity. Ann. Nutr. Metab. 2011, 58, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Rock, W.; Rosenblat, M.; Miller-Lotan, R.; Levy, A.P.; Elias, M.; Aviram, M. Consumption of wonderful variety pomegranate juice and extract by diabetic patients increases paraoxonase 1 association with high-density lipoprotein and stimulates its catalytic activities. J. Agric. Food Chem. 2008, 56, 8704–8713. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Alimbetov, D.; George, T.; Gordon, M.H.; Lovegrove, J.A. A randomised trial to investigate the effects of acute consumption of a blackcurrant juice drink on markers of vascular reactivity and bioavailability of anthocyanins in human subjects. Eur. J. Clin. Nutr. 2011, 65, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Du, M.; Leyva, M.J.; Sanchez, K.; Betts, N.M.; Wu, M.; Aston, C.E.; Lyons, T.J. Blueberries decrease cardiovascular risk factors in obese men and women with metabolic syndrome. J. Nutr. 2010, 140, 1582–1587. [Google Scholar] [CrossRef] [PubMed]
- Stull, A.J.; Cash, K.C.; Johnson, W.D.; Champagne, C.M.; Cefalu, W.T. Bioactives in blueberries improve insulin sensitivity in obese, insulin-resistant men and women. J. Nutr. 2010, 140, 1764–1768. [Google Scholar] [CrossRef] [PubMed]
- Torronen, R.; Kolehmainen, M.; Sarkkinen, E.; Poutanen, K.; Mykkanen, H.; Niskanen, L. Berries reduce postprandial insulin responses to wheat and rye breads in healthy women. J. Nutr. 2013, 143, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Torronen, R.; Sarkkinen, E.; Tapola, N.; Hautaniemi, E.; Kilpi, K.; Niskanen, L. Berries modify the postprandial plasma glucose response to sucrose in healthy subjects. Br. J. Nutr. 2010, 103, 1094–1097. [Google Scholar] [PubMed]
- Edirisinghe, I.; Banaszewski, K.; Cappozzo, J.; Sandhya, K.; Ellis, C.L.; Tadapaneni, R.; Kappagoda, C.T.; Burton-Freeman, B.M. Strawberry anthocyanin and its association with postprandial inflammation and insulin. Br. J. Nutr. 2011, 106, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.L.; Edirisinghe, I.; Kappagoda, T.; Burton-Freeman, B. Attenuation of meal-induced inflammatory and thrombotic responses in overweight men and women after 6-week daily strawberry (fragaria) intake. A randomized placebo-controlled trial. J. Atheroscler. Thromb. 2011, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Fu, D.X.; Wilkinson, M.; Simmons, B.; Wu, M.; Betts, N.M.; Du, M.; Lyons, T.J. Strawberries decrease atherosclerotic markers in subjects with metabolic syndrome. Nutr. Res. 2010, 30, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Wilkinson, M.; Penugonda, K.; Simmons, B.; Betts, N.M.; Lyons, T.J. Freeze-dried strawberry powder improves lipid profile and lipid peroxidation in women with metabolic syndrome: Baseline and post intervention effects. Nutr. J. 2009, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.; Luebke, J.L.; Morcomb, E.F.; Carrell, E.J.; Leveranz, M.C.; Kobs, L.; Schmidt, T.P.; Limburg, P.J.; Vorsa, N.; Singh, A.P. Glycemic responses to sweetened dried and raw cranberries in humans with type 2 diabetes. J. Food Sci. 2010, 75, H218–H223. [Google Scholar] [CrossRef] [PubMed]
- Ruel, G.; Pomerleau, S.; Couture, P.; Lemieux, S.; Lamarche, B.; Couillard, C. Low-calorie cranberry juice supplementation reduces plasma oxidized ldl and cell adhesion molecule concentrations in men. Br. J. Nutr. 2008, 99, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Betts, N.M.; Ortiz, J.; Simmons, B.; Wu, M.; Lyons, T.J. Low-energy cranberry juice decreases lipid oxidation and increases plasma antioxidant capacity in women with metabolic syndrome. Nutr. Res. 2011, 31, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.T.; Chan, Y.C.; Lin, C.W.; Lee, W.J.; Sheu, W.H. Effect of cranberry extracts on lipid profiles in subjects with type 2 diabetes. Diabet. Med. 2008, 25, 1473–1477. [Google Scholar] [CrossRef] [PubMed]
- Ruel, G.; Pomerleau, S.; Couture, P.; Lemieux, S.; Lamarche, B.; Couillard, C. Favourable impact of low-calorie cranberry juice consumption on plasma hdl-cholesterol concentrations in men. Br. J. Nutr. 2006, 96, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Salas-Salvado, J.; Bullo, M.; Estruch, R.; Ros, E.; Covas, M.I.; Ibarrola-Jurado, N.; Corella, D.; Aros, F.; Gomez-Gracia, E.; Ruiz-Gutierrez, V.; et al. Prevention of diabetes with mediterranean diets: A subgroup analysis of a randomized trial. Ann. Int. Med. 2014, 160, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lasa, A.; Miranda, J.; Bullo, M.; Casas, R.; Salas-Salvado, J.; Larretxi, I.; Estruch, R.; Ruiz-Gutiérrez, V.; Portillo, M. Comparative effect of two mediterranean diets versus a low-fat diet on glycaemic control in individuals with type 2 diabetes. Eur. J. Clin. Nutr. 2014, 68, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Estruch, R. Anti-inflammatory effects of the mediterranean diet: The experience of the predimed study. Proc. Nutr. Soc. 2010, 69, 333–340. [Google Scholar] [PubMed]
- Silva, S.; Bronze, M.R.; Figueira, M.E.; Siwy, J.; Mischak, H.; Combet, E.; Mullen, W. Impact of a 6-week olive oil supplementation in healthy adults on urinary proteomic biomarkers of coronary artery disease, chronic kidney disease, and diabetes (types 1 and 2): A randomized, parallel, controlled, double-blind study. Am. J. Clin. Nutr. 2015, 101, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Bulotta, S.; Celano, M.; Lepore, S.M.; Montalcini, T.; Pujia, A.; Russo, D. Beneficial effects of the olive oil phenolic components oleuropein and hydroxytyrosol: Focus on protection against cardiovascular and metabolic diseases. J. Transl. Med. 2014, 12, 219. [Google Scholar] [CrossRef] [PubMed]
- De Bock, M.; Derraik, J.G.; Brennan, C.M.; Biggs, J.B.; Morgan, P.E.; Hodgkinson, S.C.; Hofman, P.L.; Cutfield, W.S. Olive (olea europaea l.) leaf polyphenols improve insulin sensitivity in middle-aged overweight men: A randomized, placebo-controlled, crossover trial. PLoS ONE 2013, 8, e57622. [Google Scholar] [CrossRef] [PubMed]
- Wainstein, J.; Ganz, T.; Boaz, M.; Bar Dayan, Y.; Dolev, E.; Kerem, Z.; Madar, Z. Olive leaf extract as a hypoglycemic agent in both human diabetic subjects and in rats. J. Med. Food 2012, 15, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Perez-Jimenez, J.; Neveu, V.; Vos, F.; Scalbert, A. Identification of the 100 richest dietary sources of polyphenols: An application of the phenol-explorer database. Eur. J. Clin. Nutr. 2010, 64 (Suppl. S3), S112–S120. [Google Scholar] [CrossRef] [PubMed]
- Ye, E.Q.; Chacko, S.A.; Chou, E.L.; Kugizaki, M.; Liu, S. Greater whole-grain intake is associated with lower risk of type 2 diabetes, cardiovascular disease, and weight gain. J. Nutr. 2012, 142, 1304–1313. [Google Scholar] [CrossRef] [PubMed]
- McKeown, N.M.; Meigs, J.B.; Liu, S.; Wilson, P.W.; Jacques, P.F. Whole-grain intake is favorably associated with metabolic risk factors for type 2 diabetes and cardiovascular disease in the framingham offspring study. Am. J. Clin. Nutr. 2002, 76, 390–398. [Google Scholar] [PubMed]
- Giacco, R.; Lappi, J.; Costabile, G.; Kolehmainen, M.; Schwab, U.; Landberg, R.; Uusitupa, M.; Poutanen, K.; Pacini, G.; Rivellese, A.A.; et al. Effects of rye and whole wheat versus refined cereal foods on metabolic risk factors: A randomised controlled two-centre intervention study. Clin. Nutr. 2013, 32, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Giacco, R.; Costabile, G.; Della Pepa, G.; Anniballi, G.; Griffo, E.; Mangione, A.; Cipriano, P.; Viscovo, D.; Clemente, G.; Landberg, R.; et al. A whole-grain cereal-based diet lowers postprandial plasma insulin and triglyceride levels in individuals with metabolic syndrome. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, I.A.; Moore, C.; Chatfield, M.; Richardson, D.P.; Ashby, P.; Kuznesof, S.A.; Jebb, S.A.; Seal, C.J. Markers of cardiovascular risk are not changed by increased whole-grain intake: The wholeheart study, a randomised, controlled dietary intervention. Br. J. Nutr. 2010, 104, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Salas-Salvado, J.; Bullo, M.; Babio, N.; Martinez-Gonzalez, M.A.; Ibarrola-Jurado, N.; Basora, J.; Estruch, R.; Covas, M.I.; Corella, D.; Aros, F.; et al. Reduction in the incidence of type 2 diabetes with the mediterranean diet: Results of the predimed-reus nutrition intervention randomized trial. Diabetes Care 2011, 34, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Hamer, M.; Chida, Y. Intake of fruit, vegetables, and antioxidants and risk of type 2 diabetes: Systematic review and meta-analysis. J. Hypertens. 2007, 25, 2361–2369. [Google Scholar] [CrossRef] [PubMed]
- Carter, P.; Gray, L.J.; Troughton, J.; Khunti, K.; Davies, M.J. Fruit and vegetable intake and incidence of type 2 diabetes mellitus: Systematic review and meta-analysis. BMJ 2010, 341, c4229. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.J.; Forouhi, N.G.; Ye, Z.; Buijsse, B.; Arriola, L.; Balkau, B.; Barricarte, A.; Beulens, J.W.; Boeing, H.; Buchner, F.L.; et al. Fruit and vegetable intake and type 2 diabetes: Epic-interact prospective study and meta-analysis. Eur. J. Clin. Nutr. 2012, 66, 1082–1092. [Google Scholar] [CrossRef] [PubMed]
- Mursu, J.; Virtanen, J.; Tuomainen, T.P.; Nurmi, T.; Voutilainen, S. Intake of fruit, berries, and vegetables and risk of type 2 diabetes in Finnish men: The kuopio ischaemic heart disease risk factor study. Am. J. Clin. Nutr. 2014, 99, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Bozzetto, L.; Annuzzi, G.; Pacini, G.; Costabile, G.; Vetrani, C.; Vitale, M.; Griffo, E.; Giacco, A.; de Natale, C.; Cocozza, S.; et al. Polyphenol-rich diets improve glucose metabolism in people at high cardiometabolic risk: A controlled randomised intervention trial. Diabetologia 2015, 58, 1551–1560. [Google Scholar] [CrossRef] [PubMed]
- Akesson, A.; Andersen, L.F.; Kristjansdottir, A.G.; Roos, E.; Trolle, E.; Voutilainen, E.; Wirfalt, E. Health effects associated with foods characteristic of the Nordic diet: A systematic literature review. Food Nutr. Res. 2013, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brader, L.; Uusitupa, M.; Dragsted, L.O.; Hermansen, K. Effects of an isocaloric healthy Nordic diet on ambulatory blood pressure in metabolic syndrome: A randomized sysdiet sub-study. Eur. J. Clin. Nutr. 2014, 68, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Adamsson, V.; Reumark, A.; Fredriksson, I.B.; Hammarstrom, E.; Vessby, B.; Johansson, G.; Riserus, U. Effects of a healthy Nordic diet on cardiovascular risk factors in hypercholesterolaemic subjects: A randomized controlled trial (NORDIET). J. Int. Med. 2011, 269, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Kanerva, N.; Kaartinen, N.E.; Schwab, U.; Lahti-Koski, M.; Mannisto, S. Adherence to the baltic sea diet consumed in the Nordic countries is associated with lower abdominal obesity. Br. J. Nutr. 2013, 109, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Uusitupa, M.; Hermansen, K.; Savolainen, M.J.; Schwab, U.; Kolehmainen, M.; Brader, L.; Mortensen, L.S.; Cloetens, L.; Johansson-Persson, A.; Onning, G.; et al. Effects of an isocaloric healthy Nordic diet on insulin sensitivity, lipid profile and inflammation markers in metabolic syndrome—A randomized study (SYSDIET). J. Int. Med. 2013, 274, 52–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulsen, S.K.; Due, A.; Jordy, A.B.; Kiens, B.; Stark, K.D.; Stender, S.; Holst, C.; Astrup, A.; Larsen, T.M. Health effect of the new Nordic diet in adults with increased waist circumference: A 6-mo randomized controlled trial. Am. J. Clin. Nutr. 2014, 99, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Kanerva, N.; Rissanen, H.; Knekt, P.; Havulinna, A.S.; Eriksson, J.G.; Mannisto, S. The healthy Nordic diet and incidence of type 2 diabetes—10-year follow-up. Diabetes Res. Clin. Pract. 2014, 106, e34–e37. [Google Scholar] [CrossRef] [PubMed]
- Kanerva, N.; Kaartinen, N.E.; Rissanen, H.; Knekt, P.; Eriksson, J.G.; Saaksjarvi, K.; Sundvall, J.; Mannisto, S. Associations of the baltic sea diet with cardiometabolic risk factors—A meta-analysis of three Finnish studies. Br. J. Nutr. 2014, 112, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Huo, R.; Du, T.; Xu, Y.; Xu, W.; Chen, X.; Sun, K.; Yu, X. Effects of mediterranean-style diet on glycemic control, weight loss and cardiovascular risk factors among type 2 diabetes individuals: A meta-analysis. Eur. J. Clin. Nutr. 2015, 69, 1200–1208. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, Y.; Keogh, J.B.; Clifton, P.M. Polyphenols and Glycemic Control. Nutrients 2016, 8, 17. https://doi.org/10.3390/nu8010017
AMA Style
Kim Y, Keogh JB, Clifton PM. Polyphenols and Glycemic Control. Nutrients. 2016; 8(1):17. https://doi.org/10.3390/nu8010017
Chicago/Turabian StyleKim, Yoona, Jennifer B. Keogh, and Peter M. Clifton. 2016. "Polyphenols and Glycemic Control" Nutrients 8, no. 1: 17. https://doi.org/10.3390/nu8010017
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.