The Reciprocal Interactions between Polyphenols and Gut Microbiota and Effects on Bioaccessibility

 ,
,
Abstract
:1. Introduction
2. Polyphenols, Gut Microbiota and Health
3. Metabolism of Phenolics and Microbial/Colonic Metabolic Pathways


3.1. Flavonoid-Type Phenolics
3.1.1. Flavonols
3.1.2. Flavones and Flavanones
3.1.3. Flavone C-Glycosides

3.1.4. Isoflavones
3.1.5. Flavanols

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Metabolite | Model (in Vivo/in Vitro) and References |
|---|---|---|
| (+)-C or (−)-EC | 1-(4′-hydroxyphenyl)-3-(2″,4″,6″- trihydroxyphenyl)propan-2-ol | Rat in vitro [84] |
| 1-(3′-hydroxyphenyl)-3-(2″,4″,6″- trihydroxyphenyl)propan-2-ol | Rat in vitro [84] | |
| 1-(3′,4′-dihydroxyphenyl)-3-(2″,4″,6″-trihydroxyphenyl)propan-2-ol | Rat in vitro [84] | |
| 5-(3′-hydroxyphenyl)pentanoic acid | Rat in vitro [84] | |
| 5-(3′,4′-dihydroxyphenyl)-4-oxo-valeric acid | Rat in vitro [84] | |
| 5-(3′-hydroxyphenyl)-4-oxo-valeric acid | Rat in vitro [84] | |
| 5-[(3′,4′-dihydroxyphenyl)methyl]oxolan-2-one | Rat in vitro [84]; Human in vitro [87]; Human in vivo [90] | |
| 5-[(3′-hydroxyphenyl)methyl)oxolan-2-one | Rat in vitro [84]; Human in vitro [88] | |
| 5-(3′,4′-dihydroxyphenyl)-pentanoic acid | Rat in vitro [84]; Human in vitro [87] | |
| 3,4-DHPPA | Rat in vitro [84]; Pig in vitro [89] | |
| 3-HPPA | Rat in vitro [84]; Human in vitro [87] | |
| 4-HPAA | Pig in vitro [89] | |
| 3-HBA | Pig in vitro [89] | |
| 4-HBA | Pig in vitro [89] | |
| Phloroglucinol | Pig in vitro [89] | |
| 5-[(3′,4′,5′-trihydroxyphenyl)methyl]oxolan-2-one | Human in vitro [88] | |
| (+)-GC or (–)EGC | 1-(3′,5′-dihydroxyphenyl)-3-(2″,4″,6″- trihydroxyphenyl)propan-2-ol | Human in vitro [87,88] |
| 5-[(3′,4′,5′-trihydroxyphenyl)methyl]oxolan-2-one | Human in vitro [87,88] | |
| 4-HPAA | Human in vitro [87]; Pig in vitro [89] | |
| Phloroglucinol | Pig in vitro [89] | |
| 3,4-DHPPA | Pig in vitro [89] | |
| 3-HPPA | Pig in vitro [89] | |
| 3-HBA | Pig in vitro [89] | |
| 4-HBA | Pig in vitro [89] | |
| (−)-EGC | 5-[(3′,4′,5′-trihydroxyphenyl)methyl]oxolan-2-one | Human in vivo [90] |
| 5-[(3′,4′-dihydroxyphenyl)methyl]oxolan-2-one | Human in vivo [90] | |
| 5-[(3′,5′-dihydroxyphenyl)methyl)]oxolan-2-one | Human in vivo [90] | |
| (−)-ECG | EC | Rat in vivo [92,93] |
| Gallic acid | Rat in vivo [92,93] | |
| Pyrogallol | Rat in vivo [92,93] | |
| 1-(3′,4′-dihydroxyphenyl)-3-(2″,4″,6″- trihydroxyphenyl)propan-2-ol | Rat in vivo [92,93] | |
| 5-[(3′,4′-dihydroxyphenyl)methyl]oxolan-2-one | Rat in vivo [92,93] | |
| 5-[(3′-hydroxyphenyl)methyl)]oxolan-2-one | Rat in vivo [92,93] | |
| 5-(3′,4′-dihydroxyphenyl)pentanoic acid | Rat in vivo [92,93] | |
| 3-HPPA | Rat in vivo [92,93] | |
| (E)-3-(3-hydroxyphenyl)-acrylic acid | Rat in vivo [92,93] | |
| EGC | Rat in vivo [92,93] | |
| (+)-GCG or (−)-EGCG | EGC | Rat in vitro [91]; Human in vitro [87,88]; Pig in vitro [89] |
| Gallic acid | Rat in vitro [91]; Human in vitro [87,88]; Pig in vitro [89] | |
| 5-[(3′,4′,5′-trihydroxyphenyl)methyl]oxolan-2-one | Rat in vitro [91]; Pig in vitro [89] | |
| 1-(3′,4′,5′-trihydroxyphenyl)-3-(2″,4″,6″-trihydroxyphenyl)propan-2-ol | Rat in vitro [91] | |
| 1-(3′,5′-dihydroxyphenyl)-3-(2″,4″,6″- trihydroxyphenyl)propan-2-ol | Rat in vitro [91] | |
| 5-(3′,5′,- dihydroxyphenyl) pentanoic acid | Rat in vitro [91] | |
| 5-(3′,4′,5′-trihydroxyphenyl) pentanoic acid | Rat in vitro [91] | |
| 5-(3′-hydroxyphenyl)-pentanoic acid | Rat in vitro [91] | |
| 5-[(3′,5′-dihydroxyphenyl)methyl)]oxolan-2-one | Rat in vitro [91] | |
| 3,5-DHPPA | Rat in vitro [91] | |
| 5-[(3′,4′,5′-trihydroxyphenyl)methyl]oxolan-2-one | Human in vitro [87] | |
| Pyrogallol | Human in vitro [87] | |
| Pyrocatechol | Human in vitro [87] | |
| 4-HPAA | Human in vitro [87] | |
| (−)-EGCG | EGC | Rat in vivo [92,93] |
| Gallic acid | Rat in vivo [92,93] | |
| 1-(3′,4′,5′-trihydroxyphenyl)-3-(2″,4″,6″- trihydroxyphenyl)propan-2-ol | Rat in vivo [92,93] | |
| 1-(3′,5′-dihydroxyphenyl)-3-(2″,4″,6″-trihydroxyphenyl)propan-2-ol | Rat in vivo [92,93] | |
| 5-[(3′,4′,5′-trihydroxyphenyl)methyl]oxolan-2-one | Rat in vivo [92,93] | |
| 5-[(3′,5′-dihydroxyphenyl)methyl)]oxolan-2-one | Rat in vivo [92,93] | |
| 5-[(3′,4′-dihydroxyphenyl)methyl]oxolan-2-one | Rat in vivo [92,93] |
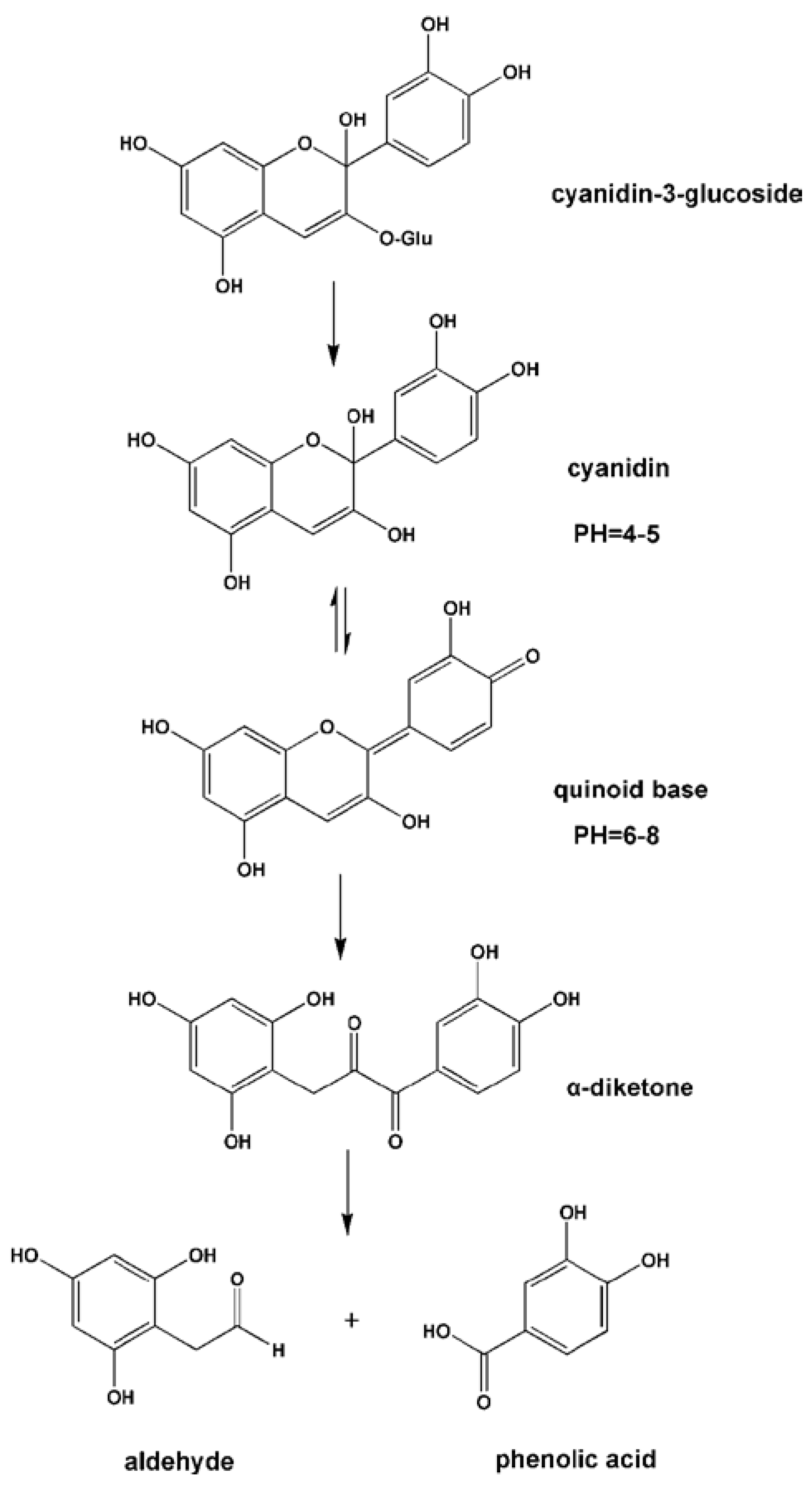
3.1.6. Anthocyanins
| Anthocyanidin | Initial B-Ring Fragmentation Product |
|---|---|
| Pelargonidin | 4-Hydroxybenzoic acid |
| Cyanidin | Protocatechuic acid |
| Delphinidin | Gallic acid |
| Peonidin | Vanillic acid |
| Petunidin | 3-Methoxy-4,5-dihydroxybenzoic acid |
| Malvidin | Syringic acid |

3.2. Nonflavonoid-Type Phenolics
3.2.1. Phenolic Acids
3.2.2. Stilbenes
3.2.3. Lignans
3.3. Limitations for the Studies on Metabolism of Phenolics and Microbial/Colonic Metabolic Pathways
4. Bioactivity and Bioavailability of Polyphenols are Affected by Gut Microbiota
5. Polyphenols Modulate the Gut Microbiota Composition
5.1. Flavonoid-Type Phenolics
5.1.1. Flavonols
5.1.2. Flavones and Flavanones
5.1.3. Isoflavones
5.1.4. Flavanols
5.1.5. Anthocyanins
5.2. Nonflavonoid-Type Phenolics
5.2.1. Phenolic Acids
5.2.2. Hydrolyzable Tannins (Ellagitannins)
5.2.3. Stilbenes
5.2.4. Lignans
| Polyphenol Type | Tested Bacteria | Growth (+)/Inhibitory (−) Effect | Type of Study | Methods Used | Duration | Doses | References |
|---|---|---|---|---|---|---|---|
| IN VITRO CELL CULTURE STUDIES | |||||||
| Flavonols | |||||||
| Quercetin | |||||||
| Bacteroides galacturonicus | (−) | In vitro | Counting on culture medium | 24 h | 4, 20 or 50 μg/mL | [166] | |
| Lactobacillus sp. | (−) | ||||||
| Enterococcus caccae | (−) | ||||||
| Bifidobacterium catenulatum | (−) | ||||||
| Ruminococcus gauvreauii | (−) | ||||||
| Escherichia coli | (−) | ||||||
| Rutin | 20, 100 or 250 μg/mL | ||||||
| Bacteroides galacturonicus, | NS | ||||||
| Lactobacillus sp. | (+) | ||||||
| Enterococcus caccae | NS | ||||||
| Bifidobacterium catenulatum | (−) | ||||||
| Ruminococcus gauvreauii | NS | ||||||
| Escherichia coli | (−) | ||||||
| Flavonols | Bifidobacterium adolescentis | In vitro | Counting on culture medium | 24 h | flavonol (galangin,kaempferol, quercetin, myricetin, or fisetin dissolved in dimethylsulphoxide (DMSO); final 25 µM; final 0.1% DMSO | [168] | |
| Galangin | (−) | ||||||
| Kaempferol | NS | ||||||
| Quercetin | NS | ||||||
| Myricetin | NS | ||||||
| Fisetin | NS | ||||||
| Isoflavones | MIC (μg/mL) | In vitro | Minimum Inhibitory Concentration Assay (MIC) | 1 h | Concentrations ranging from 62.5 to 1000 μg/mL | [170] | |
| Daidzein | Eschericia coli | 1000 | |||||
| Staphylococcus aureus | 125 | ||||||
| Salmonella typhimirum | 1000 | ||||||
| Lactobacillus rhamnosus | 1000 | ||||||
| Genistein | Eschericia coli | 1000 | |||||
| Staphylococcus aureus | 125 | ||||||
| Salmonella typhimirum | 1000 | ||||||
| Lactobacillus rhamnosus | 1000 | ||||||
| Flavanones | MIC (μg/mL) | In vitro | Minimum Inhibitory Concentration Assay (MIC) | 1 h | Concentrations ranging from 62.5 to 1000 μg/mL | [169] | |
| Naringenin | Eschericia coli | 125 | |||||
| Staphylococcus aureus | 62.5 | ||||||
| Salmonella typhimirum | 125 | ||||||
| Lactobacillus rhamnosus | 125 | ||||||
| Phenolic acids | MIC (μg/mL) | In vitro | Minimum Inhibitory Concentration Assay (MIC) | 1 h | Concentrations ranging from 62.5 to 1000 μg/mL | [169] | |
| caffeic acid | Eschericia coli | 500 | |||||
| Staphylococcus aureus | 125 | ||||||
| Salmonella typhimirum | 500 | ||||||
| Lactobacillus rhamnosus | ≤250 | ||||||
| chlorogenic acid | Eschericia coli | 1000 | |||||
| Staphylococcus aureus | 125 | ||||||
| Salmonella typhimirum | 1000 | ||||||
| Lactobacillus rhamnosus | ≤250 | ||||||
| o-coumaric acid | Eschericia coli | 250 | |||||
| Staphylococcus aureus | 125 | ||||||
| Salmonella typhimirum | 250 | ||||||
| Lactobacillus rhamnosus | 250 | ||||||
| p-coumaric acid | Eschericia coli | 500 | |||||
| Staphylococcus aureus | 125 | ||||||
| Salmonella typhimirum | 500 | ||||||
| Lactobacillus rhamnosus | 500 | ||||||
| Ellagitannins | POMx | In vitro | Liquid culturing method | POMx (100 mL) | comercial extract of pomegranate at 0.01% as well as the effect of its main constituents (0.05%) | [175] | |
| Extract of pomegranate (POMx) and its main constituents (punicalagins, punicalins, elagic acid, gallic acid) | L. acidophilus | (+) | |||||
| L. casei ssp. casei | NS | ||||||
| L. paracasei ssp. | NS | ||||||
| L. pentosus | (+) | ||||||
| L. rhamnosus | (+) | ||||||
| B. breve | (+) | ||||||
| B. infantis | (+) | ||||||
| B. longum | NS | ||||||
| B. bifidum | (+) | ||||||
| B. animalis ssp. lactis | NS | ||||||
| Bacteroides fragilis | NS | ||||||
| C. perfringens | (−) | ||||||
| Clostridium clostriidoforme | NS | ||||||
| C. ramosum | (−) | ||||||
| S. aureus | (−) | ||||||
| Punicalagin | |||||||
| L. acidophilus | NS | ||||||
| L. casei ssp. casei | NS | ||||||
| L. paracasei ssp. | NS | ||||||
| L. pentosus | NS | ||||||
| L. rhamnosus | NS | ||||||
| B. breve | (+) | ||||||
| B. infantis | NS | ||||||
| B. longum | NS | ||||||
| B. bifidum | NS | ||||||
| B. animalis ssp. lactis | (+) | ||||||
| Bacteroides fragilis | NS | ||||||
| C. perfringens | (−) | ||||||
| Clostridium clostriidoforme | (−) | ||||||
| C. ramosum | (−) | ||||||
| S. aureus | (−) | ||||||
| Punicalin | |||||||
| L. acidophilus | NS | ||||||
| L. casei ssp. casei | NS | ||||||
| L. paracasei ssp. | NS | ||||||
| L. pentosus | NS | ||||||
| L. rhamnosus | NS | ||||||
| B. breve | NS | ||||||
| B. infantis | NS | ||||||
| B. longum | NS | ||||||
| B. bifidum | NS | ||||||
| B. animalis ssp. lactis | (+) | ||||||
| Bacteroides fragilis | NS | ||||||
| C. perfringens | NS | ||||||
| Clostridium clostriidoforme | NS | ||||||
| C. ramosum | NS | ||||||
| S. aureus | NS | ||||||
| IN VITRO FAECAL MICROBIOTA STUDIES | |||||||
| Flavonols | Erysipelotrichaceae | (−) | In vitro | 16S rDNA reads | 6 weeks | 30 mg/kg BW/day | [167] |
| Quercetin | Ruminococcaceae | NS | |||||
| Clostridiaceae | NS | ||||||
| Bacteroidaceae | NS | ||||||
| Lachnospiraceae | NS | ||||||
| Acidaminococcaceae | NS | ||||||
| Eubacteriaceae | NS | ||||||
| Prevotellaceae | NS | ||||||
| Acholeplasmataceae | NS | ||||||
| Lactobacillaceae | NS | ||||||
| Graciibacteraceae | NS | ||||||
| Clostridium aldenense | NS | ||||||
| Clostridium hathewayi | NS | ||||||
| Bacteroides vulgatus | (+) | ||||||
| Clostridium clariflavum | (+) | ||||||
| Clostridium methylpentosum | NS | ||||||
| Clostridium sp. C9 | NS | ||||||
| Clostridium sp. XB90 | NS | ||||||
| Clostridium sp. MLG661 | (+) | ||||||
| Blautia stercoris | NS | ||||||
| Gracilibacter thermotolerans | NS | ||||||
| Parabacteroides distansonis | NS | ||||||
| Eubacterium cylindroides | (−) | ||||||
| Akkermansia muciniphila | NS | ||||||
| Bilophila wadsworthia | NS | ||||||
| Bacteroides sp. dnLKV7 | NS | ||||||
| Barnesiella intestinihominis | NS | ||||||
| Bacteroides sp. S-18 | NS | ||||||
| Bacteroides chinchillae | NS | ||||||
| Candidatus Prevotella conceptionensis | NS | ||||||
| Flavanols | 10 h (150 mg/L) | 150 mg/L and 1000 mg/L | [171] | ||||
| (+)-catechin | Bifidobacterium spp. | (+) | 17 h (1000 mg/L) | 150 mg/L and 1000 mg/L | |||
| Bacteroides spp. | NS | ||||||
| Lactobacillus/Enterococcus spp. | NS | ||||||
| Clostridium coccoides–Eubacterium rectale group | (+) | ||||||
| C. histolyticum group | (−) | In vitro | Fluorescent in situ hybridization (FISH) | ||||
| Escherichia coli | (+) | ||||||
| (−)-epicatechin | |||||||
| Bifidobacterium spp. | NS | ||||||
| Bacteroides spp. | NS | ||||||
| Lactobacillus/Enterococcus spp. | NS | ||||||
| Clostridium coccoides–Eubacterium rectale group | (+) | ||||||
| C. histolyticum group | NS | ||||||
| Escherichia coli | NS | ||||||
| Flavan-3-ols | Lactobacillus/Enterococcus | (+) | In vitro | Fluorescent in situ hybridization (FISH) | Samples were collected at 0, 5, 10, 24, 30 and 48 h of fermentation | 600 mg/L | [172] |
| 2 fractions of grape seed | Clostridium histolyticum | (−) | Human fecal microbiota | ||||
| Anthocyanins | In vitro | Fluorescent in situ hybridization (FISH) | 0, 1, 2, 4, 5, 10, and 24 h | 20 mg/L and 200 mg/L | [174] | ||
| Malvidin-3-glucoside | Total bacteria count | (+) | |||||
| Atopobium spp. | (+) | ||||||
| Bif idobacterium spp. | (+) | ||||||
| C. cocoides−Eubacterium rectale | (+) | ||||||
| Bacteroides spp. | (−) | ||||||
| Lactobacillus spp. | (+) | ||||||
| Clostridium histolyticum | (−) | ||||||
| Stilbenes | Erysipelotrichaceae | NS | In vitro | 16S rDNA reads | 6 weeks | 15 mg/kg BW/day | [167] |
| Trans-resveratrol | Ruminococcaceae | NS | |||||
| Clostridiaceae | NS | ||||||
| Bacteroidaceae | NS | ||||||
| Lachnospiraceae | NS | ||||||
| Acidaminococcaceae | NS | ||||||
| Eubacteriaceae | NS | ||||||
| Prevotellaceae | NS | ||||||
| Acholeplasmataceae | NS | ||||||
| Lactobacillaceae | NS | ||||||
| Graciibacteraceae | (−) | ||||||
| Clostridium aldenense | (−) | ||||||
| Clostridium hathewayi | (−) | ||||||
| Bacteroides vulgatus | NS | ||||||
| Clostridium clariflavum | NS | ||||||
| Clostridium methylpentosum | NS | ||||||
| Clostridium sp. C9 | (−) | ||||||
| Clostridium sp. XB90 | (+) | ||||||
| Clostridium sp. MLG661 | (−) | ||||||
| Blautia stercoris | NS | ||||||
| Gracilibacter thermotolerans | (−) | ||||||
| Parabacteroides distansonis | (−) | ||||||
| Eubacterium cylindroides | NS | ||||||
| Akkermansia muciniphila | NS | ||||||
| Bilophila wadsworthia | NS | ||||||
| Bacteroides sp. dnLKV7 | NS | ||||||
| Barnesiella intestinihominis | NS | ||||||
| Bacteroides sp. S-18 | NS | ||||||
| Bacteroides chinchillae | NS | ||||||
| Candidatus Prevotella conceptionensis | NS | ||||||
| Phenolic acids | Total bacteria count | (+) | In vitro | Fluorescent in situ hybridization (FISH) | 0, 1, 2, 4, 5, 10, and 24 h | 150 mg/L and 1000 mg/L | [174] |
| Gallic acid | Atopobium spp. | (+) | |||||
| Bif idobacterium spp. | (+) | ||||||
| C. cocoides−Eubacterium rectale | (+) | ||||||
| Bacteroides spp. | (−) | ||||||
| Lactobacillus spp. | (+) | ||||||
| Clostridium histolyticum | (−) | ||||||
| Ellagitannins | Total bacteria | (+) | In vitro | batch-culture fermentation system inoculated with fecal samples from healthy individuals, FISH | Samples collected at 0, 5, 10, 24 and 48 h | POMx (1.5 mL) and punicalagins (250 mg) were inoculated in stirring batch-culture vessels (one per treatment) containing faecal slurry (1:10, w/v). | [175] |
| pomegranate by-product (POMx) | Bifidobacterium spp. | (+) | |||||
| Lactobacillus spp. | (+) | ||||||
| Clostridium coccoides–Eubacterium rectale group C. histolyticum group | (+) | ||||||
| NS | |||||||
| Lignans | Lactobacillus rhamnosus VTT E-97800 | (+) | In vitro colon model | Counting on culture medium | 0, 2, 4, 6, 8, and 24 h | 8 mL of fecal suspension, and a 16.7% (w/v) final concentration of fresh fecal matter | [179] |
| Lignins | L. rhamnosus VTT E-97948 | (+) | |||||
| Lactobacillus paracasei VTT E-97949 | (+) | ||||||
| Lactobacillus salivarius VTT E-981006 | (+) | ||||||
| Bifidobacterium adolescentis VTT E-981074, Bifidobacterium breve VTT E-981075, Bifidobacterium longum VTT E-96664 | (+) | ||||||
| Lactobacillus rhamnosus VTT E-97800, | (+) | ||||||
| L. rhamnosus VTT E-97948 | (+) | ||||||
| IN VIVO STUDIES | |||||||
| Isoflavones | Clostridium coccoides-Eubacterium rectale cluster | (+) | In vivo | FISH and flow cytometry analyses | 2 months (Fecal samples were collected on day 0, 30, and 60) | 100 mg/day of isoflavones aglycon equivalents | [170] |
| Lactobacillus-Enterococcus group, | (+) | ||||||
| Faecalibacterium prausnitzii subgroup, Bifidobacterium genus | (+) | ||||||
| Clostridium coccoides-Eubacterium rectale cluster | (+) | ||||||
| Condensed Tannins | Lachnospiraceae | (+) | In vivo | culture-independent barcoded next generation sequencing | 3 days normal diet | 1% w/w Grape Seed Extract | [173] |
| Proanthocyanidins | Clostridiales | (+) | 6 days treatment diet | ||||
| Lactobacillus | (+) | 3 days post treatment control-feeding | |||||
| Ruminococcaceae | (+) | fecessamples taken daily | |||||
| Stilbenes | In vivo | Agar dilution method | 25 days | 1 mg/kg/day | [179] | ||
| Resveratrol | Lactobacillus | (+) | |||||
| Bifidobacterium | (+) | ||||||
| Enterobacteria | Diminished the increase | ||||||
| Stilbenes | Bacteroidetes-to-Firmicutes ratio, | (+) | In vivo | FISH and flow cytometry | 12 weeks | 200 mg/kg/day | [180] |
| Resveratrol | Enterococcus faecalis | (−) | |||||
| Lactobacillus | (+) | ||||||
| Bifidobacterium | (+) | ||||||
| Ellagitannins | Lactobacilli | (+) | In vivo | Agar dilution method with fecal microbiota of rats | Samples collected at days 0, 10, 20 | 250 mg/kg/day | [177] |
| Pomegranate ellagitannins and their microbiota-derived metabolite urolithin A | Bifidobacterium | (+) | |||||
| Ellagitannins | Actinobacteria | (+) | In vivo | FISH and flow cytometry | 4 weeks | 1000 mg POM extract | [178] |
| Pomegranate (POM) ellagitannins | Firmicutes | (−) | |||||
| Verrucomicrobia (Akkermansia muciniphila) | (+) | ||||||
| Butyrivibrio | (+) | ||||||
| Enterobacter | (+) | ||||||
| Eschericia | (+) | ||||||
| Lactobacillus | (+) | ||||||
| Prevotella | (+) | ||||||
| Serratia | (+) | ||||||
| Veillonella | (+) | ||||||
| Collinsella | (−) | ||||||
5.3. Limitations for the Studies on Gut Microbiota Composition Modulation by Polyphenols
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 3,4-DHPPA | 3,4-dihydroxyphenylpropionic acid |
| 3-HPPA | 3-hydroxyphenylpropionic acid |
| 4-HPPA | 4-hydroxyphenylpropionic acid |
| 3-HBA | 3-hydroxybenzoic acid |
| 4-HBA | 4-hydroxybenzoic acid |
| 3,4-DHPPA | 3,4-dihydroxyphenylacetic acid |
| 3-HPAA | 3-hydroxyphenylacetic acid |
| 4-HPAA | 4-hydroxyphenylacetic acid |
| FISH | fluorescent in situ hybridisation |
References
- Vinson, J.A.; Su, X.; Zubik, L.; Bose, P. Phenol antioxidant quantity and quality in foods: Fruits. J. Agric. Food Chem. 2001, 49, 5315–5321. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Manach, C.; Morand, C.; Remesy, C.; Jimenez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Venkataraman, K.; Hollingsworth, A.; Piche, M.; Tai, T.C. Polyphenols: Benefits to the cardiovascular system in health and in ageing. Nutrients 2013, 5, 3779–3827. [Google Scholar] [CrossRef] [PubMed]
- Meydani, M.; Hasan, S.T. Dietary polyphenols and obesity. Nutrients 2010, 2, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Li, A.-N.; Li, S.; Zhang, Y.-J.; Xu, X.-R.; Chen, Y.-M.; Li, H.-B. Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Remesy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230–242. [Google Scholar]
- Parada, J.; Aguilera, J.M. Food microstructure affects the bioavailability of several nutrients. J. Food Sci. 2007, 72, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073–2085. [Google Scholar]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Van Duynhoven, J.; Vaughan, E.E.; Jacobs, D.M.; Kemperman, R.A.; van Velzen, E.J.J.; Gross, G.; Roger, L.C.; Possemiers, S.; Smilde, A.K.; Doré, J.; et al. Metabolic fate of polyphenols in the human superorganism. Proc. Natl. Acad. Sci. USA 2011, 108, 4531–4538. [Google Scholar] [CrossRef] [PubMed]
- Mosele, J.; Macià, A.; Motilva, M.-J. Metabolic and microbial modulation of the large intestine ecosystem by non-absorbed diet phenolic compounds: A review. Molecules 2015, 20, 17429–17468. [Google Scholar] [CrossRef] [PubMed]
- Moco, S.; Martin, F.P.J.; Rezzi, S. Metabolomics view on gut microbiome modulation by polyphenol-rich foods. J. Proteome Res. 2012, 11, 4781–4790. [Google Scholar] [CrossRef] [PubMed]
- Tuohy, K.M.; Conterno, L.; Gasperotti, M.; Viola, R. Up-regulating the human intestinal microbiome using whole plant foods, polyphenols, and/or fiber. J. Agric. Food Chem. 2012, 60, 8776–8782. [Google Scholar] [CrossRef] [PubMed]
- Valdés, L.; Cuervo, A.; Salazar, N.; Ruas-Madiedo, P.; Gueimonde, M.; González, S. The relationship between phenolic compounds from diet and microbiota: Impact on human health. Food Funct. 2015, 6, 2424–2439. [Google Scholar] [CrossRef] [PubMed]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, M.; Muñoz-González, I.; Cueva, C.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolome, B. A survey of modulation of gut microbiota by dietary polyphenols. Biomed. Res. Int. 2015, 2015, 1–15. [Google Scholar]
- He, X.; Marco, M.L.; Slupsky, C.M. Emerging aspects of food and nutrition on gut microbiota. J. Agric. Food Chem. 2013, 61, 9559–9574. [Google Scholar] [CrossRef] [PubMed]
- Possemiers, S.; Bolca, S.; Verstraete, W.; Heyerick, A. The intestinal microbiome: A separate organ inside the body with the metabolic potential to influence the bioactivity of botanicals. Fitoterapia 2011, 82, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Alminger, M.; Aura, A.M.; Bohn, T.; Dufour, C.; El, S.N.; Gomes, A.; Karakaya, S.; Martines-Cuesta, M.C.; McDougall, G.J.; Requena, T.; et al. In vitro models for studying secondary plant metabolite digestion and bioaccessibility. Compr. Rev. Food Sci. Food Saf. 2014, 13, 413–436. [Google Scholar] [CrossRef]
- Puupponen-Pimiä, R.; Aura, A.M.; Oksman-Caldentey, K.M.; Myllärinen, P.; Saarela, M.; Mattila-Sandholm, T.; Poutenen, K. Development of functional ingredients for gut health. Trends Food Sci. Technol. 2002, 13, 3–11. [Google Scholar] [CrossRef]
- Chen, D.; Wan, S.B.; Yang, H.; Yuan, J.; Chan, T.H.; Dou, Q.P. EGCG, green tea polyphenols and their synthetic analogs and prodrugs for human cancer prevention and treatment. Adv. Clin. Chem. 2011, 53, 155–177. [Google Scholar] [PubMed]
- Weng, C.J.; Yen, G.C. Chemopreventive effects of dietary phytochemicals against cancer invasion and metastasis: Phenolic acids, monophenol, polyphenol and their derivatives. Cancer Treat. Rev. 2012, 38, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Kuriyama, S.; Shimazu, T.; Ohmori, K.; Kikuchi, N.; Nakaya, N.; Nishino, Y.; Tsubono, Y.; Tsuji, I. Green tea consumption and mortality due to cardiovascular disease, cancer and all causes in Japan. J. Am. Med. Assoc. 2006, 296, 1255–1265. [Google Scholar] [CrossRef] [PubMed]
- Mursu, J.; Voutilainen, S.; Nurmi, T.; Tuomainen, T.P.; Kurt, S.; Salonen, J.T. Flavonoid intake and the risk of ischaemic stroke and CVD mortality in middle-ages Finnish men. J. Nutr. 2008, 100, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.Y.; Jin, J.S.; Cho, Y.A.; Lee, J.H.; Park, S.; Jeong, S.W.; Kim, Y.-H.; Lim, C.-S.; El-Aty, A.M.; Kim, G.-S.; et al. Determination of polyphenols in three Capsicum annuum L. (bell pepper) varieties using high-performance liquid chromatography–tandem mass spectrometry: Their contribution to overall antioxidant and anticancer activity. J. Sep. Sci. 2011, 34, 2967–2974. [Google Scholar] [CrossRef] [PubMed]
- Ogunleye, A.A.; Xue, F.; Michels, K.B. Green tea consumption and breast cancer risk or recurrence: A meta-analysis. Breast Cancer Res. Treat. 2009, 119, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zubik, L.; Collins, F.W.; Marko, M.; Meydani, M. The antiatherogenic potential of oat phenolic compounds. Atherosclerosis 2004, 175, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Huff, M.W. Antiatherogenic properties of flavonoids: Implications for cardiovascular health. Can. J. Cardiol. 2010, 26, 17–21. [Google Scholar] [CrossRef]
- Zakaria, Z.A.; Hisam, E.E.A.; Rofiee, M.S.; Norhafizah, M.; Somchit, M.N.; Teh, L.K.; Salleh, M.Z. In vivo antiulcer activity of the aqueous extract of Bauhinia purpurea leaf. J. Ethnopharmacol. 2011, 137, 1047–1054. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Gu, Y.; Ye, C.; Cao, Y.; Liu, Z.; Yin, J. Antithrombotic activity of fractions and components obtained from raspberry leaves (Rubus chingii). Food Chem. 2012, 132, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.W.; Duan, J.A.; Yang, N.Y.; Tang, Y.P.; Liu, M.Z.; Qian, Y.F. Antithrombotic phenolic compounds from Glycyrrhiza uralensis. Fitoterapia 2012, 83, 422–425. [Google Scholar] [CrossRef] [PubMed]
- Beara, I.N.; Lesjak, M.M.; Orcic, D.Z.; Simin, N.D.; Cetoyevic-Simin, D.D.; Bozin, B.; Mimica-Dukic, N.M. Comparative analysis of phenolic profile, antioxidant, anti-inflammatory and cytotoxic activity of two closely-related plantain species: Plantago altissima L. and Plantago lanceolata L. LWT Food Sci. Technol. 2012, 47, 64–70. [Google Scholar] [CrossRef]
- Zimmer, A.R.; Leonardi, B.; Miron, D.; Schapoval, E.; Oliveira, J.R.; Gosmann, G. Antioxidant and anti-inflammatory properties of Capsicum baccatum: From traditional use to scientific approach. J. Ethnopharmacol. 2012, 139, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.Y.; Champagne, E.T. Reducing the allergenic capacity of peanut extracts and liquid peanut butter by phenolic compounds. Food Chem. 2009, 115, 1345–1349. [Google Scholar] [CrossRef]
- Schmitz-Eiberger, M.A.; Blanke, M.M. Bioactive components in forced sweet cherry fruit (Prunus avium L.) antioxidative capacity and allergenic potential as dependent on cultivation under cover. LWT Food Sci. Technol. 2012, 46, 388–392. [Google Scholar] [CrossRef]
- Bijak, M.; Bobrowski, M.; Borowiecka, M.; Podsędek, A.; Golański, J.; Nowak, P. Anticoagulant effect of polyphenols-rich extracts from black chokeberry and grape seeds. Fitoterapia 2011, 82, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Schütz, K.; Saß, M.; With, A.; Graubaum, H.J.; Grünwald, J. Immunemodulating efficacy of a polyphenol-rich beverage on symptoms associated with the common cold: A double-blind, randomised, placebo-controlled, multi-centric clinical study. Br. J. Nutr. 2010, 104, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Rodrigues, S.; Feas, X.; Estevinho, L.M. Antimicrobial activity, phenolic profile and role in the inflammation of propolis. Food Chem. Toxicol. 2012, 50, 1790–1795. [Google Scholar] [CrossRef] [PubMed]
- Xia, D.; Wu, X.; Shi, J.; Yang, Q.; Zhang, Y. Phenolic compounds from the edible seeds extract of Chinese Mei (Prunus mume Sieb. Et Zucc) and their antimicrobial activity. LWT Food Sci. Technol. 2011, 44, 347–349. [Google Scholar] [CrossRef]
- Mudnic, I.; Modun, D.; Rastija, V.; Vukovic, J.; Brizic, I.; Katalinic, V.; Kozina, B.; Medic-Saric, M.; Boban, M. Antioxidant and vasodilatory effects of phenolic acids in wine. Food Chem. 2010, 119, 1205–1210. [Google Scholar] [CrossRef]
- Santoz, M.D.; Almeida, M.C.; Lopez, N.P.; Souza, G.E.P. Evaluation of the anti-inflammatory, analgesic and antipyretic activities of the natural polyphenols CGA. Biol. Pharm. Bull. 2010, 29, 2236–2240. [Google Scholar] [CrossRef]
- Selma, M.V.; Espín, J.C.; Tomás-Barberán, F.A. Interaction between phenolics and gut microbiota: Role in human health. J. Agric. Food. Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef] [PubMed]
- Faria, A.; Fernandes, I.; Norberto, S.; Mateus, N.; Calhau, C. Interplay between anthocyanins and gut microbiota. J. Agric. Food Chem. 2014, 62, 6898–6902. [Google Scholar] [CrossRef] [PubMed]
- Guinane, C.M.; Cotter, P.D. Role of the gut microbiota in health and chronic gastrointestinal disease: Under-standing a hidden metabolic organ. Ther. Adv. Gastroenterol. 2013, 6, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Delzenne, N.M. The role of the gut microbiota in energy metabolism and metabolic disease. Curr. Pharm. Des. 2009, 15, 1546–1558. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arboleya, S.; Binetti, A.; Salazar, N.; Fernandez, N.; Solis, G.; Hernandez-Barranco, A.; Margolles, A.; de los Reyes-Gavilan, C.G.; Gueimonde, M. Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiol. Ecol. 2012, 79, 763–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Prebiotic evaluation of cocoa-derived flavanols in healthy humans by using a randomized, controlled, double-blind, crossover intervention study. Am. J. Clin. Nutr. 2011, 93, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Queipo-Ortuño, M.I.; Boto-Ordóñez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Diaz, F.C.; Andres-Lacueva, C.; Tinahones, F.J. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am. J. Clin. Nutr. 2012, 95, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Rastmanesh, R. High polyphenol, low probiotic diet for weight loss because of intestinal microbiota interaction. Chem. Biol. Interact. 2011, 189, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hervert-Hernandez, D.; Goñi, I. Dietary polyphenols and human gut microbiota: A review. Food. Rev. Int. 2011, 27, 154–169. [Google Scholar] [CrossRef]
- Laparra, J.M.; Sanz, Y. Interactions of gut microbiota with functional food components and nutraceuticals. Pharmacol. Res. 2010, 61, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Gotteland, M.; Andrews, M.; Toledo, M.; Muñoz, L.; Caceres, P.; Anziani, A.; Wittig, E.; Speisky, H.; Salazar, G. Modulation of Helicobacter pylori colonization with cranberry juice and Lactobacillus johnsonii La1 in children. Nutrition 2008, 24, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Vitali, B.; Ndagijimana, M.; Cruciani, F.; Carnevali, P.; Candela, M.; Guerzoni, M.E.; Brigidi, P. Impact of a synbiotic food on the gut microbial ecology and metabolic profiles. BMC Microbiol. 2010, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Hollman, P.C.; Katan, M.B. Dietary flavonoids: Intake, health effects and bioavailability. Food Chem. Toxicol. 1999, 37, 937–942. [Google Scholar] [CrossRef]
- Winter, J.; Moore, L.H.; Dowell, V.R.; Bokkenheuser, V.D. C-Ring cleavage of flavonoids by human Intestinal bacteria. Appl. Environ. Microbiol. 1989, 55, 1203–1208. [Google Scholar] [PubMed]
- Winter, J.; Popoff, M.R.; Grimont, P.; Bokkenheuser, V.D. Clostridium orbiscindens sp. Nov., a human intestinal bacterium capable of cleaving the flavonoid C-ring. Int. J. Syst. Bacteriol. 1991, 41, 355–357. [Google Scholar] [CrossRef] [PubMed]
- Rechner, A.R.; Smith, M.A.; Kuhnle, G.; Gibson, G.R.; Debnam, E.S.; Srai, S.K.S.; Moore, K.P.; Rice-Evans, C.A. Colonic metabolism of dietary polyphenol: Influence of structure on microbial fermentation products. Free Radic. Biol. Med. 2004, 36, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Toma’s-Barbera’n, F.A.; Clifford, M.N. Flavanones, chalcones and dihydrochalcones nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1073–1080. [Google Scholar] [CrossRef]
- Robards, K.; Li, X.; Antolovich, M.; Boyd, S. Characterisation of citrus by chromatographic analysis of flavonoids. J. Sci. Food Agric. 1997, 75, 87–101. [Google Scholar] [CrossRef]
- Xiao, J.B. Dietary flavonoid aglycones and their glycosides: What show better biological benefits? Crit. Rev. Food Sci. Nutr. 2016. [Google Scholar] [CrossRef]
- Xiao, J.B.; Högger, P. Dietary polyphenols and type 2 diabetes: Current insights and future perspectives. Curr. Med. Chem. 2015, 22, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.B.; Capanoglu, E.; Jassbi, A.R.; Miron, A. Advance on the flavonoid C-glycosides and health benefits. Crit. Rev. Food Sci. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tie, X.W.; Bao, B.L.; Wu, X.Q.; Zhang, Y. Metabolism of flavone C-glucosides and p-coumaric acid from antioxidant of bamboo leaves (AOB) in rats. Br. J. Nutr. 2007, 97, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Wang, J.; Liu, X. Metabolism of dietary soy isoflavones to equol by human intestinal microflora; implications for health. Mol. Nutr. Food Res. 2007, 51, 765–781. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, F.H.; Li, Y.W. Mechanisms of cancer chemoprevention by soy isoflavone genistein. Cancer Metastasis Rev. 2002, 21, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Messina, M. Soyfoods and soybean phyto-oestrogens (isoflavones) as possible alternatives to hormone replacement therapy (HRT). Eur. J. Cancer 2000, 36, 71–77. [Google Scholar] [CrossRef]
- Cornwell, T.; Cohick, W.; Raskin, I. Dietary phytoestrogens and health. Phytochemistry 2004, 65, 995–1016. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.R.; Brown, N.M.; Zimmer-Nechemias, L.; Brashear, W.T.; Wolfe, B.; Kirscher, A.S.; Heubi, J.E. Evidence for lack of absorption of soy isoflavone glycosides in humans, supporting the crucial role of intestinal metabolism for bioavailability. Am. J. Clin. Nutr. 2002, 76, 447–453. [Google Scholar] [PubMed]
- Linford, N.J.; Dorsa, D.M. 17β-Estradiol and the phytoestrogen genistein attenuate neuronal apoptosis induced by the endoplasmic reticulum calcium-ATPase inhibitor thapsigargin. Steroids 2002, 67, 1029–1040. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Brown, N.M.; Lydeking-Olsen, E. The clinical importance of the metabolite equol-A clue to the effectiveness of soy and its isoflavones. J. Nutr. 2002, 32, 3577–3584. [Google Scholar]
- Zubik, L.; Meydani, M. Bioavailability of soybean isoflavones from aglycone and glucoside forms in American women. Am. J. Clin. Nutr. 2003, 77, 1459–1465. [Google Scholar] [PubMed]
- Hur, H.; Rafii, F. Biotransformation of the isoflavonoids biochanin A, formononetin and glycitein by Eubacterium limosum. FEMS Microbiol. Lett. 2000, 192, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Simons, A.L.; Renouf, M.; Hendrich, S.; Murphy, P.A. Metabolism of glycitein (7;4′-dihydroxy-6-methoxy-isoflavone) by human gut microflora. J. Agric. Food Chem. 2005, 53, 8519–8525. [Google Scholar] [CrossRef] [PubMed]
- Heinonen, S.; Hoikkala, A.; Wahala, K.; Adlercreutz, H. Metabolism of the soy isoflavones daidzein, genistein and glycitein in human subjects. Identification of new metabolites having an intact isoflavonoid skeleton. J. Steroid Biochem. Mol. Biol. 2003, 87, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Kuhnle, G.; Spencer, J.P.; Schroeter, H.; Shenoy, B.; Debnam, E.S.; Srai, S.K.; Rice-Evans, C.; Hahn, U. Epicatechin and catechin are O-methylated and glucuronidated in the small intestine. Biochem. Biophys. Res. Commun. 2000, 277, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, J.B.; Walle, T. Glucuronidation and sulfation of the tea flavonoid (−)-epicatechin by the human and rat enzymes. Drug Metab. Dispos. 2002, 30, 897–890. [Google Scholar] [CrossRef] [PubMed]
- Hackman, R.M.; Polagruto, J.A.; Zhu, Q.Y.; Sun, B.; Fujii, H.; Keen, C.L. Flavanols: Digestion, absorption and bioactivity. Phytochem. Rev. 2008, 7, 195–208. [Google Scholar] [CrossRef]
- Garcıá-Ramirez, B.; Fernández-Larrea, J.; Salvado, M.J.; Ardevol, A.; Arola, L.; Blade, C. Tetramethylated dimeric procyanidins are detected in rat plasma and liver early after oral administration of synthetic oligomeric procyanidins. J. Agric. Food Chem. 2006, 54, 2543–2551. [Google Scholar] [CrossRef] [PubMed]
- Deprez, S.; Brezillon, C.; Rabot, S.; Philippe, C.; Mila, I.; Lapierre, C.; Scalbert, A. Polymeric proanthocyanidins are catabolized by human colonic microflora into low-molecular-weight phenolic acids. J. Nutr. 2000, 130, 2733–2738. [Google Scholar] [PubMed]
- Rios, L.Y.; Gonthier, M.P.; Remesy, C.; Mila, I.; Lapierre, C.; Lazarus, S.A.; Williamson, G.; Scalbert, A. Chocolate intake increases urinary excretion of polyphenol-derived phenolic acids in healthy human subjects. Am. J. Clin. Nutr. 2003, 77, 912–918. [Google Scholar] [PubMed]
- Meselhy, M.R.; Nakamura, N.; Hattori, M. Biotransformation of (−)-epicatechin-3-O-gallate by human intestinal bacteria. Chem. Pharm. Bull. 1997, 45, 888–893. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Lambert, J.D.; Ho, C.T.; Yang, C.S. The chemistry and biotransformation of tea constituents. Pharmacol. Res. 2011, 64, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Takagaki, A.; Nanjo, F. Catabolism of (+)-catechin and (−)-epicatechin by rat intestinal microbiota. J. Agric. Food Chem. 2013, 61, 4927–4935. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Q.; Meselhy, M.R.; Li, Y.; Nakamura, N.; Min, B.S.; Qin, G.W.; Hattori, M. The heterocyclic ring fission and dehydroxylation of catechins and related compounds by Eubacterium sp. strain SDG-2, a human intestinal bacterium. Chem. Pharma. Bull. 2001, 49, 1640–1643. [Google Scholar] [CrossRef]
- Kutschera, M.; Engst, W.; Blaut, M.; Braune, A. Isolation of catechin-converting human intestinal bacteria. J. Appl. Microbiol. 2011, 111, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Roowi, S.; Stalmach, A.; Mullen, W.; Lean, M.E.; Edwards, C.A.; Crozier, A. Green tea flavan-3-ols: Colonic degradation and urinary excretion of catabolites by humans. J. Agric. Food. Chem. 2010, 58, 1296–1304. [Google Scholar] [CrossRef] [PubMed]
- Schantz, M.; Erk, T.; Richling, E. Metabolism of green tea catechins by the human small intestine. Biotechnol. J. 2010, 5, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Van’t Slot, G.; Humpf, H.U. Degradation and metabolism of catechin, epigallocatechin-3-gallate (EGCG), and related compounds by the intestinal microbiota in the pig cecum model. J. Agric. Food. Chem. 2009, 57, 8041–8048. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Sang, S.; Zhu, N.; Lu, H.; Sheng, S.; Lee, M.J.; Ho, C.T.; Yang, C.S. Identification and characterization of methylated and ring-fission metabolites of tea catechins formed in humans, mice, and rats. Chem. Res. Toxicol. 2002, 15, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Takagaki, A.; Nanjo, F. Metabolism of (−)-epigallocatechin gallate by rat intestinal flora. J. Agric. Food Chem. 2010, 58, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Kohri, T.; Matsumoto, N.; Yamakawa, M.; Suzuki, M.; Nanjo, F.; Hara, Y.; Oku, N. Metabolic fate of (−)-[4-(3)H]epigallocatechin gallate in rats after oral administration. J. Agric. Food. Chem. 2001, 49, 4102–4112. [Google Scholar] [CrossRef]
- Kohri, T.; Nanjo, F.; Suzuki, M.; Seto, R.; Matsumoto, N.; Yamakawa, M.; Hojo, H.; Hara, Y.; Desai, D.; Amin, S.; et al. Synthesis of (−)-[4–3H]epigallocatechin gallate and its metabolic fate in rats after intravenous administration. J. Agric. Food. Chem. 2001, 49, 1042–1048. [Google Scholar] [CrossRef] [PubMed]
- Fleschhut, J.; Kratzer, F.; Rechkemmer, G.; Kulling, S.E. Stability and biotransformation of various dietary anthocyanins in vitro. Eur. J. Nutr. 2006, 45, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Keppler, K.; Humpf, H.U. Metabolism of anthocyanins and their phenolic degradation products by the intestinal microflora. Bioorg. Med. Chem. 2005, 13, 5195–5205. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Nair, M.G.; Strasburg, G.M.; Chang, Y.C.; Booren, A.M.; Gray, J.I.; DeWitt, D.L. Antioxidant and antiinflammatory activities of anthocyanins and their aglycon, cyanidin, from tart cherries. J. Nat. Prod. 1999, 62, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.X. Potential mechanisms of cancer chemoprevention by anthocyanins. Curr. Mol. Med. 2003, 3, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Katsube, N.; Iwashita, K.; Tsushida, T.; Yamaki, K.; Kobori, M. Induction of apoptosis in cancer cells by Bilberry (Vaccinium. myrtillus) and the anthocyanins. J. Agric. Food Chem. 2003, 51, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.B.; Högger, P. Advance in pharmacokinetics of bioactive polyphenols. Curr. Drug Metab. 2014, 15, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Inaba, H.; Kishi, M.; Tominaga, S.; Hirayama, M.; Tsuda, T. Orally administered delphinidin 3-rutinoside and cyanidin 3-rutinoside are directly absorbed in rats and humans and appear in the blood as the intact forms. J. Agric. Food Chem. 2001, 49, 1546–1551. [Google Scholar] [CrossRef] [PubMed]
- Vitaglione, P.; Donnarumma, G.; Napolitano, A.; Galvano, F.; Gallo, A.; Scalfi, L.; Fogliano, V. Protocatechuic acid is the major human metabolite of cyanidin-glucosides. J. Nutr. 2007, 137, 2043–2048. [Google Scholar] [PubMed]
- Kahle, K.; Kraus, M.; Scheppach, W.; Ackermann, M.; Ridder, F.; Richling, E. Studies on apple and blueberry fruit constituents: Do the polyphenols reach the colon after ingestion? Mol. Nutr. Food Res. 2006, 50, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Aura, A.M.; Martin-Lopez, P.; O’Leary, K.A.; Williamson, G.; Oksman-Caldentey, K.M.; Poutanen, K.; Santos-Buelga, C. In vitro metabolism of anthocyanins by human gut microflora. Eur. J. Nutr. 2005, 44, 133–142. [Google Scholar] [CrossRef] [PubMed]
- El Mohsen, M.A.; Marks, J.; Kuhnle, G.; Moore, K.; Debnam, E.; Srai, S.K.; Rice-Evans, C.; Spencer, J.P. Absorption, tissue distribution and excretion of pelargonidin and its metabolites following oral administration to rats. Br. J. Nutr. 2006, 95, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Hassimotto, N.M.A.; Genovese, M.I.; Lajolo, F.M. Absorption and metabolism of cyanidin-3-glucoside and cyanidin-3-rutinoside extracted from wild mulberry (Morus nigra L.) in rats. Nutr. Res. 2008, 28, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Hanske, L.; Engst, W.; Loh, G.; Sczesny, S.; Blaut, M.; Braune, A. Contribution of gut bacteria to the metabolism of cyanidin 3-glucoside in human microbiota-associated rats. Br. J. Nutr. 2013, 109, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Pittman Lii, H.E.; Hager, T.; Hager, A.; Howard, L.; Prior, R.L. Phenolic acids in black raspberry and in the gastrointestinal tract of pigs following ingestion of black raspberry. Mol. Nutr. Food Res. 2009, 53, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T.; Horio, F.; Osawa, T. Absorption and metabolism of cyanidin 3-O-beta-d-glucoside in rats. FEBS Lett. 1999, 449, 179–182. [Google Scholar] [CrossRef]
- Han, S.J.; Ryu, S.N.; Trinh, H.T.; Joh, E.H.; Jang, S.Y.; Han, M.J.; Kim, D.H. Metabolism of cyanidin-3-O-beta-d-glucoside isolated from black colored rice and its antiscratching behavioral effect in mice. J. Food Sci. 2009, 74, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Clifford, M.N. Colonic metabolites of berry polyphenols: The missing link to biological activity? Br. J. Nutr. 2010, 104, 48–66. [Google Scholar] [CrossRef] [PubMed]
- Andreasen, M.F.; Kroon, P.A.; Williamson, G.; Garcia-Conesa, M.-T. Intestinal release and uptake of phenolic antioxidant diferulic acids. Free Radic. Biol. Med. 2001, 31, 304–314. [Google Scholar] [CrossRef]
- Rondini, L.; Peyrat-Maillard, M.-N.; Marsset-Baglieri, A.; Fromentin, G.; Durand, P.; Tome, P.; Prost, M.; Berset, C. Bound ferulic acid from bran is more bioavailable than the free compound in rat. J. Agric. Food Chem. 2004, 52, 4338–4343. [Google Scholar] [CrossRef] [PubMed]
- Andreasen, M.F.; Kroon, P.A.; Williamson, G.; Garcia-Conesa, M.T. Esterase activity able to hydrolyze dietary antioxidant hydroxycinnamates is distributed along the intestine of mammals. J. Agric. Food Chem. 2001, 49, 5679–5684. [Google Scholar] [CrossRef] [PubMed]
- Gonthier, M.P.; Remesy, C.; Scalbert, A.; Cheynier, V.; Souquet, J.M.; Poutanen, K.; Aura, A.M. Microbial metabolism of caffeic acid and its esters chlorogenic and caftaric acids by human faecal microbiota in vitro. Biomed. Pharmacotherapy 2006, 60, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Peppercorn, M.A.; Goldman, P. Caffeic acid metabolism by bacteria of the human gastrointestinal tract. J. Bacteriol. 1971, 108, 996–1000. [Google Scholar] [PubMed]
- Andreasen, M.F.; Christensen, L.P.; Meyer, A.S.; Hansen, A. Content of phenolic acids and ferulic acid dehydrodimers in 17 Rye (Secale cereale L.) varieties. J. Agric. Food Chem. 2000, 48, 2837–2842. [Google Scholar] [CrossRef] [PubMed]
- Braune, A.; Bunzel, M.; Yonekura, R.; Blaut, M. Conversion of dehydrodiferulic acids by human intestinal microbiota. J. Agric. Food Chem. 2009, 57, 3356–3362. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.Y.; Bolling, B.W. Characterisation of stilbenes in California almonds (Prunus dulcis) by UHPLC-MS. Food Chem. 2015, 148, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Mulat, D.G.; Latva-Maenpaa, H.; Koskela, H.; Saranpaa, P.; Wahala, K. Rapid chemical characterisation of stilbenes in the root bark of Norway spruce by off-line HPLC/DAD-NMR. Phytochem. Anal. 2014, 25, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Folmer, F.; Basavaraju, U.; Jaspars, M.; Hold, G.; El-Omar, E.; Dicato, M.; Diederich, M. Anticancer effects of bioactive berry compounds. Phytochem. Rev. 2014, 13, 295–322. [Google Scholar] [CrossRef]
- Fu, Y.J.; Kadioglu, O.; Wiench, B.; Wei, Z.F.; Gao, C.; Luo, M.; Gu, C.B.; Zu, Y.G.; Efferth, T. Cell cycle arrest and induction of apoptosis by cajanin stilbene acid from Cajanus cajun in breast cancer cells. Phytomedicine 2015, 22, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wang, Y.R.; Gao, J.Y.; Lu, Z.Y.; Yin, W.P.; Deng, R.X. Resveratrol trimers from seed cake of Paeonia rockii. Molecules 2014, 19, 19549–19556. [Google Scholar] [CrossRef] [PubMed]
- Bode, L.M.; Bunzel, D.; Huch, M.; Cho, G.S.; Ruhland, D.; Bunzel, M.; Bub, A.; Franz, C.M.A.P.; Kulling, S.E. In vivo and in vitro metabolism of trans-resveratrol by human gut microbiota. Am. J. Clin. Nutr. 2013, 97, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.G.; Zhang, Z.W.; Ju, J.F.; Wang, X.Y.; Qiu, W.J. Investigation of piceid metabolites in rat by liquid chromatography tandem mass spectrometry. J. Chromatogr. B 2011, 879, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; Hanley, B.; Lamuela-Raventos, R.M. Isoflavones: Lignans and stilbenes; origins, metabolism and potential importance to human health. J. Sci. Food Agric. 2000, 80, 1044–1062. [Google Scholar] [CrossRef]
- Touillaud, M.S.; Thiebaut, A.C.M.; Fournier, A.; Niravong, M.; Boutron-Ruault, M.C.; Clavel-Chapelon, F. Dietary lignan intake and postmenopausal breast cancer risk by estrogen and progesterone receptor status. J. Natl. Cancer Inst. 2007, 99, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Aura, A.-M.; Oikarinen, S.; Mutanen, M.; Heinonen, S.-M.; Ad-lercreutz, H.C.T.; Virtanen, H.; Poutanen, K.S. Suitability of a batch in vitro fermentation model using human faecal microbiota for prediction of conversion of flaxseed lignans to enterolactone with reference to an in vivo rat model. Eur. J. Nutr. 2006, 45, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Possemiers, S.; Bolca, S.; Eeckhaut, E.; Depypere, H.; Verstraete, W. Metabolism of isoflavones, lignans and prenylflavonoids by intestinal bacteria: Producer phenotyping and relation with intestinal community. FEMS Microbiol. Ecol. 2007, 6, 1372–1383. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Faughnan, M.; Hoey, L.; Wahala, K.; Williamson, G.; Cassidy, A. Bioavailability of phytoestrogens. Br. J. Nutr. 2003, 89, 45–58. [Google Scholar]
- Bowey, E.; Adlercreutz, H.; Rowland, I. Metabolism of isoflavones and lignans by the gut microflora: A study in germ-free and human flora associated rats. Food Chem. Toxicol. 2003, 41, 631–636. [Google Scholar] [CrossRef]
- Eeckhaut, E.; Struijs, K.; Possemiers, S.; Vincken, J.P.; Keukeleire, D.D.; Verstraete, W. Metabolism of the lignan macromolecule into enterolignans in the gastrointestinal lumen as determined in the simulator of the human intestinal microbial ecosystem. J. Agric. Food Chem. 2008, 56, 4806–4812. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Q.; Meselhy, M.R.; Li, Y.; Qin, G.W.; Hattori, M. Human intestinal bacteria capable of transforming secoisolariciresinol diglucoside to mammalian lignans, enterodiol and enterolactone. Chem. Pharm. Bull. 2000, 48, 1606–1610. [Google Scholar] [CrossRef] [PubMed]
- Blaut, M.; Clavel, T. Metabolic diversity of the intestinal microbiota: Implications for health and disease. J. Nutr. 2007, 137, 751–755. [Google Scholar]
- Bohn, T. Bioavailability of non-provitamin a carotenoids. Curr. Nutr. Food Sci. 2008, 4, 240–258. [Google Scholar] [CrossRef]
- Bouayed, J.; Hoffmann, L.; Bohn, T. Total phenolics, flavonoids, anthocyanins and antioxidant activity following simulated gastro-intestinal digestion and dialysis of apple varieties: Bioaccessibility and potential uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Kemperman, R.A.; Bolca, S.; Roger, L.C.; Vaughan, E.E. Novel approaches for analysing gut microbes and dietary polyphenols: Challenges and opportunities. Microbiology 2010, 156, 3224–3231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, G.; Manach, C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr. 2005, 81, 243–255. [Google Scholar]
- Hollman, P.C.; Katan, M.B. Absorption, metabolism and health effects of dietary flavonoids in man. Biomed. Pharmacother. 1997, 51, 305–310. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [PubMed]
- Lampe, J.W. Interindividual differences in response to plant-based diets: Implications for cancer risk. Am. J. Clin. Nutr. 2009, 89, 1553–1557. [Google Scholar] [CrossRef] [PubMed]
- Padayachee, A.; Netzel, G.; Netzel, M.; Day, L.; Zabaras, D.; Mikkelsen, D.; Gidley, M.J. Binding of polyphenols to plant cell wall analogues—Part 2: Phenolic acids. Food Chem. 2012, 135, 2287–2292. [Google Scholar] [CrossRef] [PubMed]
- Gil-Izquierdo, A.; Gil, M.I.; Ferreres, F.; Tomas-Barberan, F.A. In vitro availability of flavonoids and other phenolics in orange juice. J. Agric. Food Chem. 2001, 49, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Mandalaria, G.; Tomaino, A.; Rich, G.T.; Lo Curto, R.; Arcoraci, T.; Martorana, M.; Bisignano, C.; Saija, A.; Parker, M.L.; Waldron, K.W.; et al. Polyphenol and nutrient release from skin of almonds during simulated human digestion. Food Chem. 2010, 122, 1083–1088. [Google Scholar] [CrossRef]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein–phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Saura-Calixto, F. Dietary fiber as a carrier of dietary antioxidants: An essential physiological function. J. Agric. Food Chem. 2011, 59, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Ortega, N.; Reguant, J.; Romero, M.P.; Macia, A.; Motilva, M.J. Effect of fat content on the digestibility and bioaccessibility of cocoa polyphenol by an in vitro digestion model. J. Agric. Food Chem. 2009, 57, 5743–5749. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Urpi-Sarda, M.; Boto-Ordonez, M.; Knox, C.; Llorach, R.; Eisner, R.; Cruz, J.; Neveu, V.; Wishart, D.; Manach, C.; et al. Phenol-Explorer 2.0: A major update of the Phenol-Explorer database integrating data on polyphenol metabolism and pharmacokinetics in humans and experimental animals. Database 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Bjursell, M.K.; Himrod, J.; Deng, S.; Carmichael, L.K.; Chiang, H.C.; Hooper, L.V.; Gordon, J.I. A genomic view of the human-Bacteroides thetaiotaomicron symbiosis. Science 2003, 299, 2074–2076. [Google Scholar] [CrossRef] [PubMed]
- Martens, E.C.; Lowe, E.C.; Chiang, H.; Pudlo, N.A.; Wu, M.; McNulty, N.P.; Abbott, D.W.; Henrissat, B.; Gilbert, H.J.; Bolam, D.N.; et al. Recognition and degradation of plant cell wall polysaccharides by two human gut symbionts. PLoS Biol. 2011, 9, 2492. [Google Scholar] [CrossRef] [PubMed]
- Donohoe, D.R.; Garge, N.; Zhang, X.; Sun, W.; O’Connell, T.M.; Bunger, M.K.; Bultman, S.J. The microbiome and butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell Metab. 2011, 13, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Manchester, J.K.; Semenkovich, C.F.; Gordon, J.I. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci. USA 2007, 104, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Esposito, D.; Damsud, T.; Wilson, M.; Grace, M.H.; Strauch, R.; Li, X.; Lila, M.A.; Komarnytsky, S. Black currant anthocyanins attenuate weight gain and improve glucose metabolism in diet-induced obese mice with intact, but not disrupted, gut microbiome. J. Agric. Food Chem. 2015, 63, 6172–6180. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, I.A.; Mena, P.; Calani, L.; Borges, G.; Pereira-Caro, G.; Bresciani, L.; del Rio, D.; Lean, M.E.J.; Crozier, A. New insights into the bioavailability of red raspberry anthocyanins and ellagitannins. Free Radic. Biol. Med. 2015, 89, 758–769. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Tomás-Barbéran, F.A.; Espín, J.C. Metabolism of antioxidant and chemopreventive ellagitannins from strawberries, raspberries, walnuts, and oak-aged wine in humans: Identification of biomarkers and individual variability. J. Agric. Food Chem. 2005, 53, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Henning, S.M.; Zhang, Y.; Suchard, M.; Li, Z.; Heber, D. Pomegranate juice ellagitannin metabolites are present in human plasma and some persist in urine for up to 48 h. J. Nutr. 2006, 136, 2481–2485. [Google Scholar] [PubMed]
- González-Barrio, R.; Borges, G.; Mullen, W.; Crozier, A. Bioavailability of anthocyanins and ellagitannins following consumption of raspberries by healthy humans and subjects with an ileostomy. J. Agric. Food. Chem. 2010, 58, 3933–3939. [Google Scholar] [CrossRef] [PubMed]
- González-Barrio, R.; Edwards, C.A.; Crozier, A. Colonic catabolism of ellagitannins, ellagic acid, and raspberry anthocyanins: In vivo and in vitro studies. Drug Metab. Dispos. 2011, 39, 1680–1688. [Google Scholar] [CrossRef] [PubMed]
- Dudonné, S.; Varin, T.V.; Anhê, F.F.; Dubé, P.; Roy, D.; Pilon, G.; Marette, A.; Levy, E.; Jacquot, C.; Urdaci, M.; et al. Modulatory effects of a cranberry extract co-supplementation with Bacillus subtilis CU1 probiotic on phenolic compounds bioavailability and gut microbiota composition in high-fat diet-fed mice. PharmaNutrition 2015, 3, 89–100. [Google Scholar] [CrossRef]
- Halliwell, B.; Rafter, J.; Jenner, A. Health promotion by flavonoids, tocopherols, tocotrienols, and other phenols: Direct or indirect effects? Antioxidant or not? Am. J. Clin. Nutr. 2005, 81, 268–276. [Google Scholar]
- Choy, Y.Y.; Jaggers, G.K.; Oteiza, P.I.; Waterhouse, A.L. Bioavailability of intact proanthocyanidins in the rat colon after ingestion of grape seed extract. J. Agric. Food Chem. 2013, 61, 121–127. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Magnuson, B.A.; Giusti, M.M. Analysis of anthocyanins in rat intestinal contentsimpact of anthocyanin chemical structure on fecal excretion. J. Agric. Food Chem. 2005, 53, 2859–2866. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R. Prebiotics as gut microflora management tools. J. Clin. Gastroenterol. 2008, 42, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Burcelin, R.; Garidou, L.; Pomie, C. Immuno-microbiota cross and talk: The new paradigm of metabolic diseases. Semin. Immunol. 2012, 24, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Rastall, R.A.; Gibson, G.R.; Gill, H.S.; Guarner, F.; Klaenhammer, T.R.; Pot, B.; Reid, G.; Rowland, I.R.; Sanders, M.E. Modulation of the microbial ecology of the human colon by probiotics, prebiotics and synbiotics to enhance human health: An overview of enabling science and potential applications. FEMS Microbiol. Ecol. 2005, 52, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Duda-Chodak, A. The inhibitory effect of polyphenols on human gut microbiota. J. Physiol. Pharmacol. 2012, 63, 497–503. [Google Scholar] [PubMed]
- Etxeberria, U.; Arias, N.; Boqué, N.; Macarulla, M.T.; Portillo, M.P.; Martínez, J.A.; Milagro, F.I. Reshaping faecal gut microbiota composition by the intake of trans-resveratrol and quercetin in high-fat sucrose diet-fed rats. J. Nutr. Biochem. 2015, 26, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Sugiyama, Y.; Sakano, T.; Ohigashi, H. Flavonols enhanced production of anti-inflammatory substance(s) by Bifidobacterium adolescentis: Prebiotic actions of galangin, quercetin, and fisetin. Biofactors 2013, 39, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Parkar, S.G.; Stevenson, D.E.; Skinner, M.A. The potential influence of fruit polyphenols on colonic microflora and human gut health. Int. J. Food Microbiol. 2008, 124, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Clavel, T.; Fallani, M.; Lepage, P.; Levenez, F.; Mathey, J.; Rochet, V.; Serezat, M.; Sutren, M.; Henderson, G.; Bennetau-Pelissero, C.; et al. Isoflavones and functional foods alter the dominant intestinal microbiota in postmenopausal women. J. Nutr. 2005, 135, 2786–2792. [Google Scholar] [PubMed]
- Tzounis, X.; Vulevic, J.; Kuhnle, G.G.C.; George, T.; Leonczak, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P.M. Flavanol monomer-induced changes to the human faecal microflora. Br. J. Nutr. 2008, 99, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Cueva, C.; Sánchez-Patán, F.; Monagas, M.; Walton, G.E.; Gibson, G.R.; Martín-Álvarez, P.J.; Bartolome, B.; Moreno-Arribas, M.V. In vitro fermentation of grape seed flavan-3-ol fractions by human faecal microbiota: Changes in microbial groups and phenolic metabolites. FEMS Microbiol. Ecol. 2013, 83, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Choy, Y.Y.; Quifer-Rada, P.; Holstege, D.M.; Frese, S.A.; Calvert, C.C.; Mills, D.A.; Lamuela-Raventos, R.M.; Waterhouse, A.L. Phenolic metabolites and substantial microbiome changes in pig feces by ingesting grape seed proanthocyanidins. Food. Funct. 2014, 5, 2298–2308. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.; de Pascual-Teresa, S. Metabolism of anthocyanins by human gut microflora and their influence on gut bacterial growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef] [PubMed]
- Bialonska, D.; Kasimsetty, S.G.; Schrader, K.K.; Ferreira, D. The effect of pomegranate (Punica granatum L.) byproducts and ellagitannins on the growth of human gut bacteria. J. Agric. Food Chem. 2009, 57, 8344–8349. [Google Scholar] [CrossRef] [PubMed]
- Bialonska, D.; Ramnani, P.; Kasimsetty, S.G.; Muntha, K.R.; Gibson, G.R.; Ferreira, D. The influence of pomegranate by-product and punicalagins on selected groups of human intestinal microbiota. Int. J. Food Microbiol. 2010, 140, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; Gonzalez-Sarrıas, A.; Yanez-Gascon, M.J.; Selma, M.V.; Azorın-Ortuno, M.; Toti, S.; Tomas-Barberan, F.A.; Dolara, P.; Espın, J.C. Anti-inflammatory properties of a pome-granate extract and its metabolite urolithin-A in a colitis rat model and the effect of colon inflammation on the phenolic metabolism. J. Nutr. Biochem. 2010, 21, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Henning, S.M.; Lee, R.-P.; Lu, Q.-Y.; Summanen, P.H.; Thames, G.; Corbett, K.; Downes, J.; Tsena, C.-H.; Finegold, S.M.; et al. Pomegranate extract induces metabolite formation and changes stool microbiota in healthy volunteers. Food Funct. 2015, 6, 2487–2495. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; Yanez-Gascon, M.J.; Selma, M.V.; Gonzalez-Sarrıas, A.; Toti, S.; Ceron, J.J.; Tomas-Barberan, F.A.; Dolara, P.; Espın, J.C. Effect of a low dose of dietary resveratrol on colon microbiota, inflammation and tissue damage in a DSS-induced colitis rat model. J. Agric. Food Chem. 2009, 57, 2211–2220. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Sun, J.; Xia, S.; Tang, X.; Shi, Y.; Le, G. Effects of resveratrol on gut microbiota and fat storage in a mouse model with high-fat-induced obesity. Food Funct. 2014, 5, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Niemi, P.; Aura, A.M.; Maukonen, J.; Smeds, A.I.; Mattila, I.; Niemelä, K.; Tamminen, T.; Fauulds, C.B.; Buchert, J.; Poutanen, K. Interactions of a lignin-rich fraction from Brewer’s spent grain with gut microbiota in vitro. J. Agric. Food Chem. 2013, 61, 6754–6762. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The Reciprocal Interactions between Polyphenols and Gut Microbiota and Effects on Bioaccessibility. Nutrients 2016, 8, 78. https://doi.org/10.3390/nu8020078
Ozdal T, Sela DA, Xiao J, Boyacioglu D, Chen F, Capanoglu E. The Reciprocal Interactions between Polyphenols and Gut Microbiota and Effects on Bioaccessibility. Nutrients. 2016; 8(2):78. https://doi.org/10.3390/nu8020078
Chicago/Turabian StyleOzdal, Tugba, David A. Sela, Jianbo Xiao, Dilek Boyacioglu, Fang Chen, and Esra Capanoglu. 2016. "The Reciprocal Interactions between Polyphenols and Gut Microbiota and Effects on Bioaccessibility" Nutrients 8, no. 2: 78. https://doi.org/10.3390/nu8020078