Splenic Immune Response Is Down-Regulated in C57BL/6J Mice Fed Eicosapentaenoic Acid and Docosahexaenoic Acid Enriched High Fat Diet

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethical Declaration
2.2. Diet
2.3. Splenic Fatty Acid Profiling
2.4. RNA Isolation, Quality Assurance and Microarray Analysis
2.5. Quantitative Real-Time PCR (qRT-PCR)
2.6. Statistical Analysis
3. Results
3.1. Physiological Changes in Mice—Effects of High Fat Diets Differing in Their Fat Composition
3.2. Spleen Fatty Acid Profiles
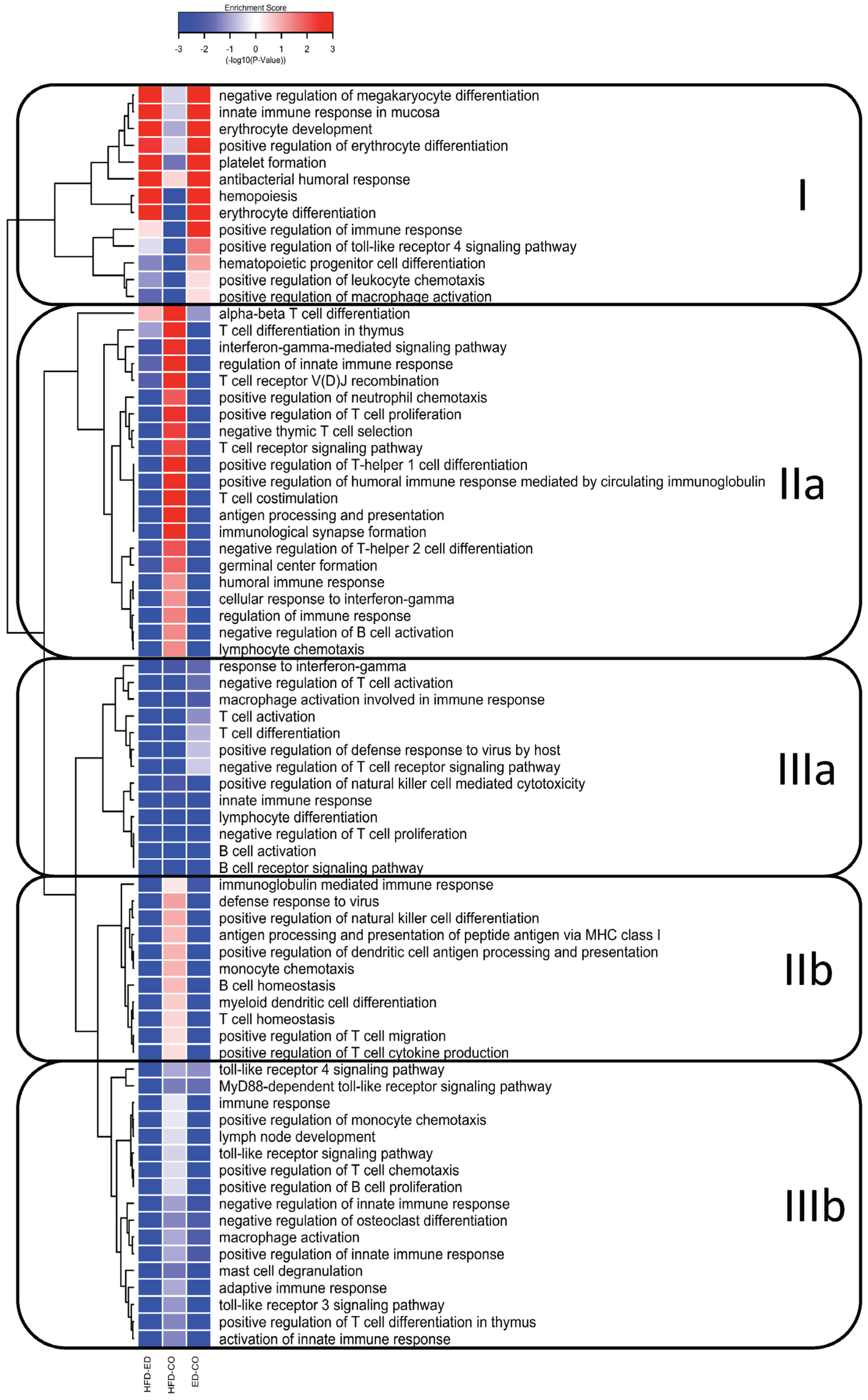
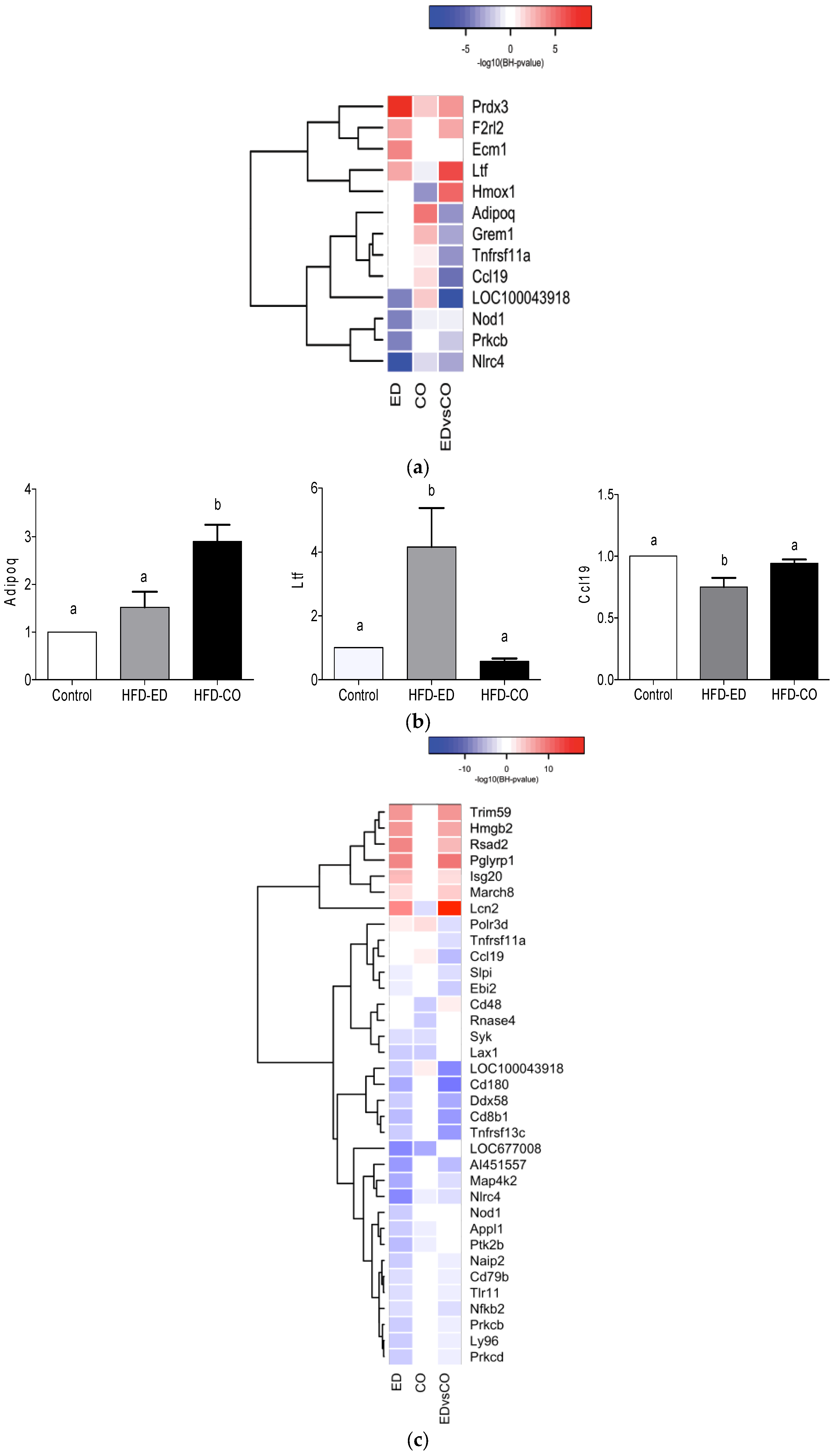
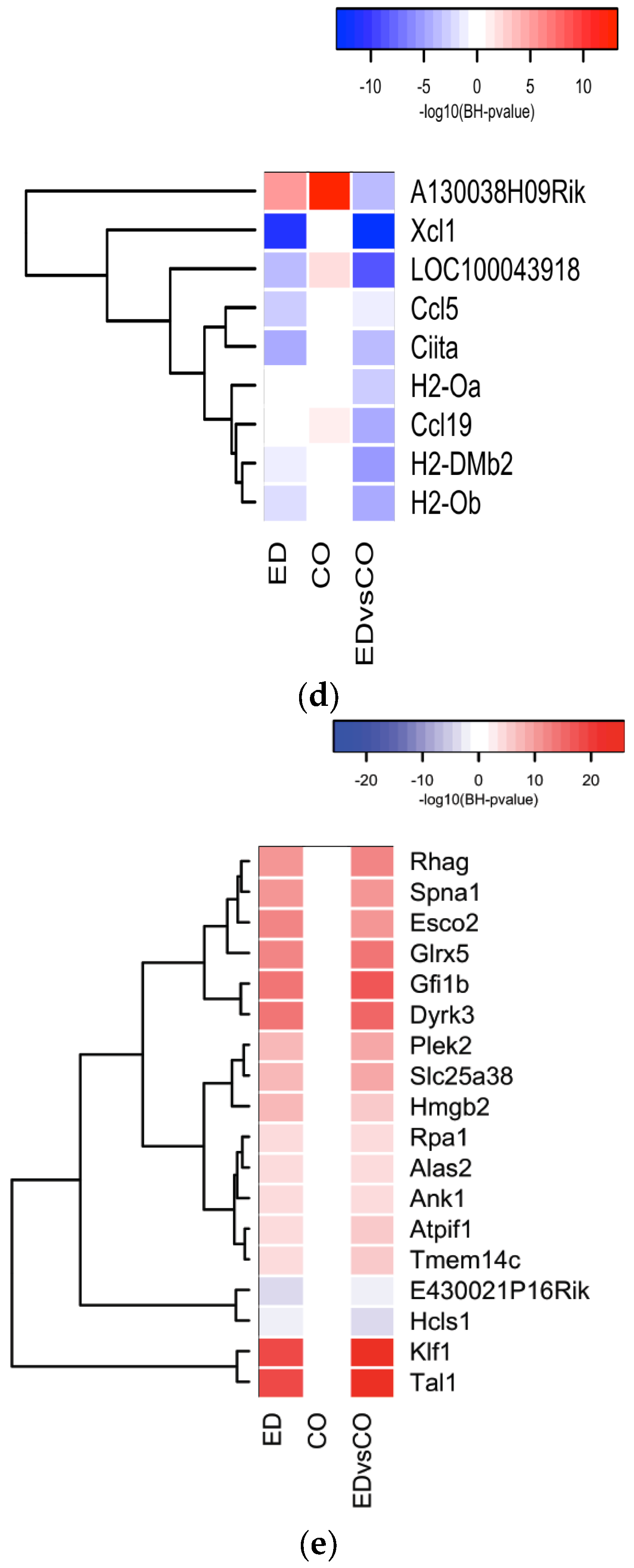
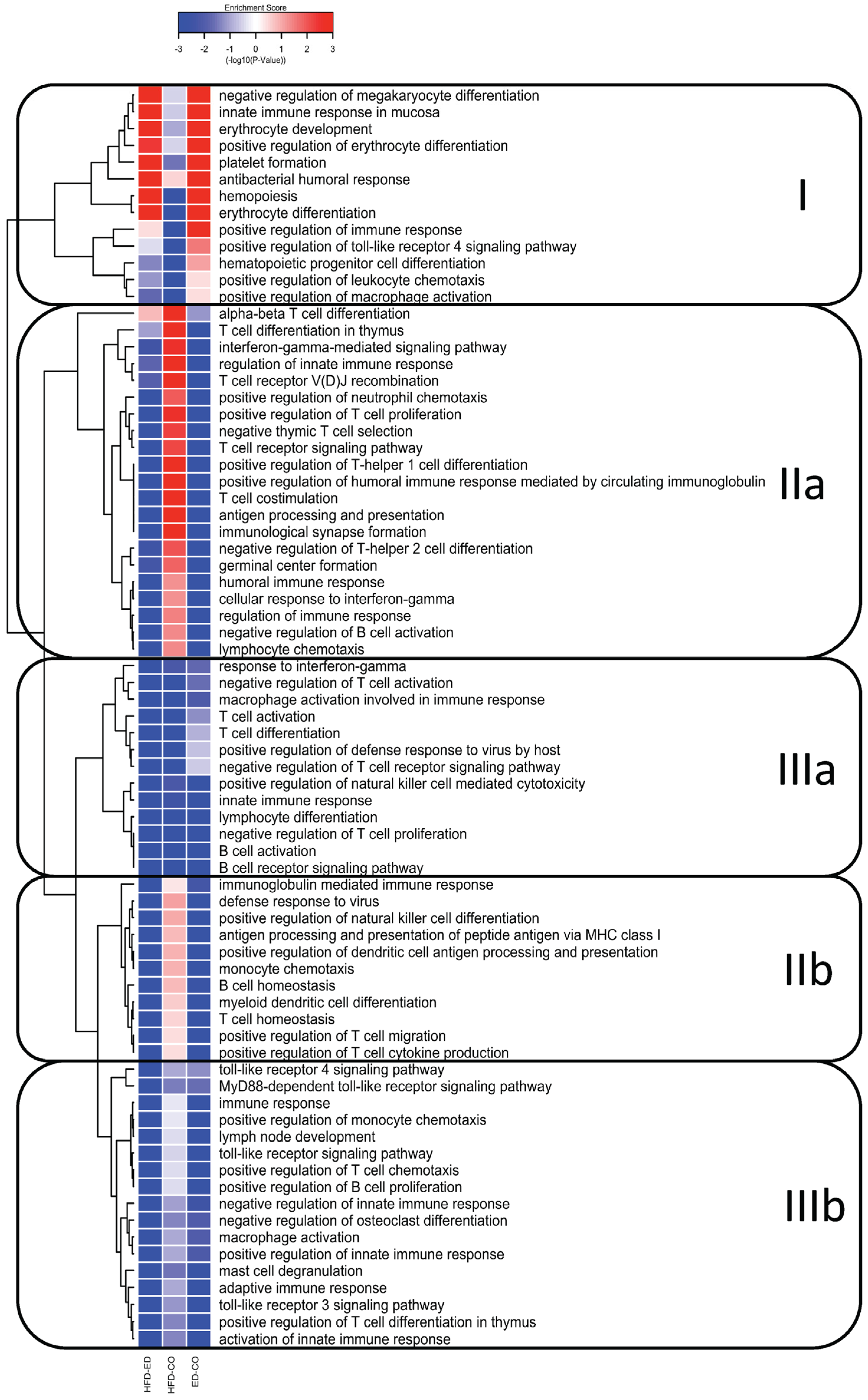
3.3. Global Gene Expression Analysis of the Spleen Transcriptome
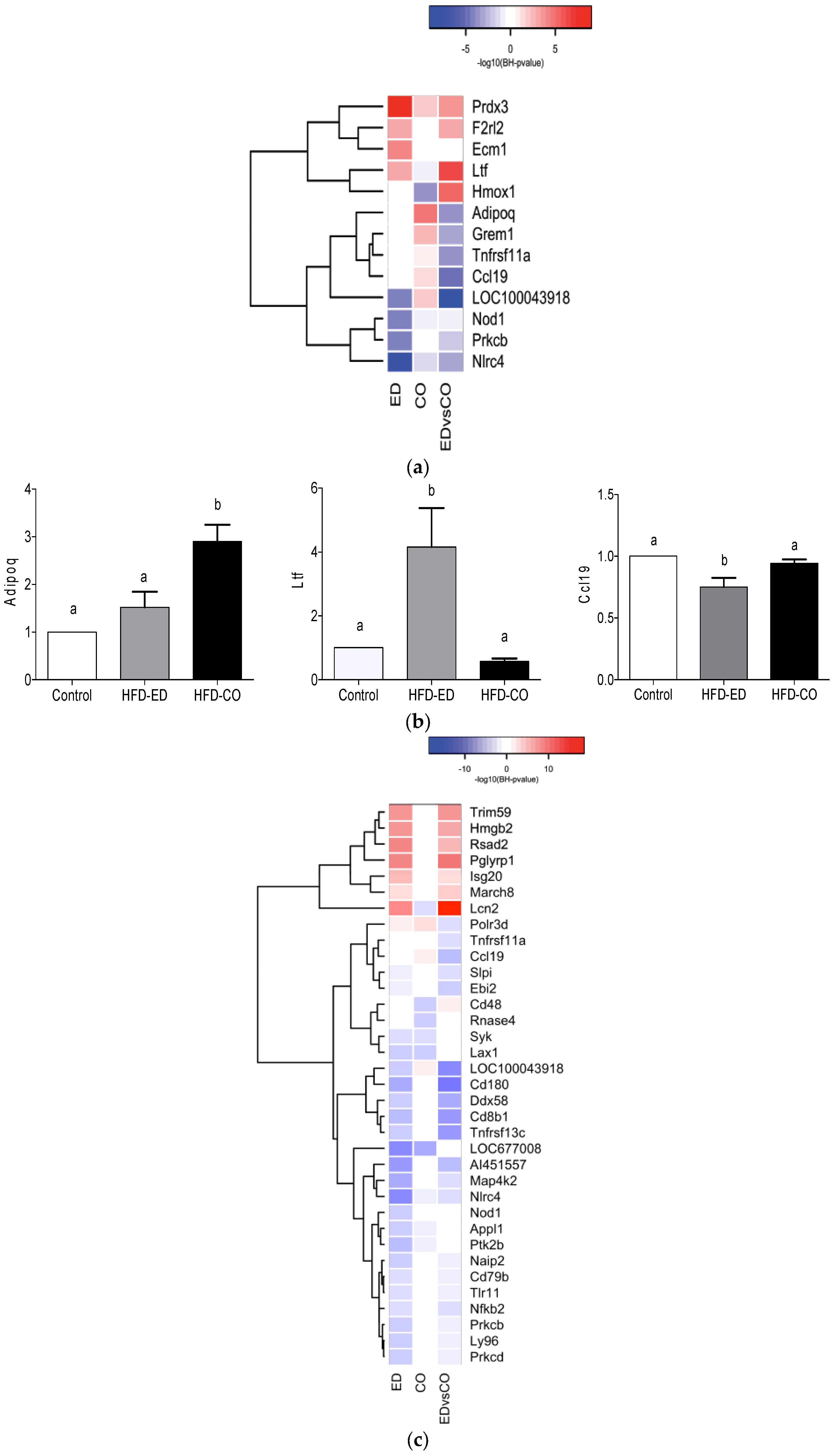
3.4. Immune System Related Gene Expression in the Spleen
3.5. Modulation of NF-κB Related Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nathan, C. Points of control in inflammation. Nature 2002, 420, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Shoelson, S.E.; Lee, J.; Goldfine, A.B. Inflammation and insulin resistance. J. Clin. Investig. 2006, 116, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A.; Nguyen, K.D.; Goh, Y.P. Macrophage-mediated inflammation in metabolic disease. Nat. Rev. Immunol. 2011, 11, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Sowers, J.R. Obesity as a cardiovascular risk factor. Am. J. Med. 2003, 115, 37S–41S. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Noncommunicable Diseases. Fact Sheet fs355. 2015. Available online: http://www.who.int/mediacentre/factsheets/fs355/en/ (accessed on 6 September 2016).
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Duncan, B.B.; Schmidt, M.I.; Pankow, J.S.; Ballantyne, C.M.; Couper, D.; Vigo, A.; Hoogeveen, R.; Folsom, A.R.; Heiss, G.; Atherosclerosis Risk in Communities Study. Low-grade systemic inflammation and the development of type 2 diabetes: The atherosclerosis risk in communities study. Diabetes 2003, 52, 1799–1805. [Google Scholar] [CrossRef] [PubMed]
- Vozarova, B.; Weyer, C.; Lindsay, R.S.; Pratley, R.E.; Bogardus, C.; Tataranni, P.A. High white blood cell count is associated with a worsening of insulin sensitivity and predicts the development of type 2 diabetes. Diabetes 2002, 51, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Spranger, J.; Kroke, A.; Mohlig, M.; Hoffmann, K.; Bergmann, M.M.; Ristow, M.; Boeing, H.; Pfeiffer, A.F. Inflammatory cytokines and the risk to develop type 2 diabetes: Results of the prospective population-based European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam Study. Diabetes 2003, 52, 812–817. [Google Scholar] [CrossRef] [PubMed]
- Herder, C.; Baumert, J.; Thorand, B.; Koenig, W.; de Jager, W.; Meisinger, C.; Illig, T.; Martin, S.; Kolb, H. Chemokines as risk factors for type 2 diabetes: Results from the MONICA/KORA Augsburg study, 1984–2002. Diabetologia 2006, 49, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Sohn, K.H.; Rhee, S.H.; Hwang, D. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through Toll-like receptor 4. J. Biol. Chem. 2001, 276, 16683–16689. [Google Scholar] [CrossRef] [PubMed]
- Nagakura, T.; Matsuda, S.; Shichijyo, K.; Sugimoto, H.; Hata, K. Dietary supplementation with fish oil rich in omega-3 polyunsaturated fatty acids in children with bronchial asthma. Eur. Respir. J. 2000, 16, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.M.; Ma, D.W. Are all n-3 polyunsaturated fatty acids created equal? Lipids Health Dis. 2009, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Duda, M.K.; O’Shea, K.M.; Stanley, W.C. Omega-3 polyunsaturated fatty acid supplementation for the treatment of heart failure: Mechanisms and clinical potential. Cardiovasc. Res. 2009, 84, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Saremi, A.; Arora, R. The utility of omega-3 fatty acids in cardiovascular disease. Am. J. Ther. 2009, 16, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Yashodhara, B.M.; Umakanth, S.; Pappachan, J.M.; Bhat, S.K.; Kamath, R.; Choo, B.H. Omega-3 fatty acids: A comprehensive review of their role in health and disease. Postgrad. Med. J. 2009, 85, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.Y.; McMurray, D.N.; Ly, L.H.; Chapkin, R.S. Dietary (n-3) polyunsaturated fatty acids remodel mouse T-cell lipid rafts. J. Nutr. 2003, 133, 1913–1920. [Google Scholar] [PubMed]
- Fan, Y.Y.; Ly, L.H.; Barhoumi, R.; McMurray, D.N.; Chapkin, R.S. Dietary docosahexaenoic acid suppresses T cell protein kinase C theta lipid raft recruitment and IL-2 production. J. Immunol. 2004, 173, 6151–6160. [Google Scholar] [CrossRef] [PubMed]
- Teague, H.; Harris, M.; Fenton, J.; Lallemand, P.; Shewchuk, B.M.; Shaikh, S.R. Eicosapentaenoic and docosahexaenoic acid ethyl esters differentially enhance B-cell activity in murine obesity. J. Lipid Res. 2014, 55, 1420–1433. [Google Scholar] [CrossRef] [PubMed]
- Svahn, S.L.; Varemo, L.; Gabrielsson, B.G.; Peris, E.; Nookaew, I.; Grahnemo, L.; Sandberg, A.S.; Wernstedt Asterholm, I.; Jansson, J.O.; Nielsen, J.; et al. Six Tissue Transcriptomics Reveals Specific Immune Suppression in Spleen by Dietary Polyunsaturated Fatty Acids. PLoS ONE 2016, 11, e0155099. [Google Scholar] [CrossRef] [PubMed]
- Soni, N.K.; Nookaew, I.; Sandberg, A.S.; Gabrielsson, B.G. Eicosapentaenoic and docosahexaenoic acid-enriched high fat diet delays the development of fatty liver in mice. Lipids Health Dis. 2015, 14, 74. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [PubMed]
- Nookaew, I.; Gabrielsson, B.G.; Holmäng, A.; Sandberg, A.-S.; Nielsen, J. Identifying Molecular Effects of Diet through Systems Biology: Influence of Herring Diet on Sterol Metabolism and Protein Turnover in Mice. PLoS ONE 2010, 5, e12361. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsson, B.G.; Wikstrom, J.; Jakubowicz, R.; Marmon, S.K.; Carlsson, N.G.; Jansson, N.; Gan, L.M.; Undeland, I.; Lonn, M.; Holmang, A.; et al. Dietary herring improves plasma lipid profiles and reduces atherosclerosis in obese low-density lipoprotein receptor-deficient mice. Int. J. Mol. Med. 2012, 29, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Soni, N.; Ross, A.; Scheers, N.; Savolainen, O.; Nookaew, I.; Gabrielsson, B.; Sandberg, A.-S. Eicosapentaenoic and Docosahexaenoic Acid-Enriched High Fat Diet Delays Skeletal Muscle Degradation in Mice. Nutrients 2016, 8, 543. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Kibbe, W.A.; Lin, S.M. Lumi: A pipeline for processing Illumina microarray. Bioinformatics 2008, 24, 1547–1548. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Varemo, L.; Nielsen, J.; Nookaew, I. Enriching the gene set analysis of genome-wide data by incorporating directionality of gene expression and combining statistical hypotheses and methods. Nucleic Acids Res. 2013, 41, 4378–4391. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013. [Google Scholar] [CrossRef] [PubMed]
- Cesta, M.F. Normal structure, function, and histology of the spleen. Toxicol. Pathol. 2006, 34, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Oarada, M.; Furukawa, H.; Majima, T.; Miyazawa, T. Fish oil diet affects on oxidative senescence of red blood cells linked to degeneration of spleen cells in mice. Biochim. Biophys. Acta 2000, 1487, 1–14. [Google Scholar] [CrossRef]
- Fowler, K.H.; Chapkin, R.S.; McMurray, D.N. Effects of purified dietary n-3 ethyl esters on murine T lymphocyte function. J. Immunol. 1993, 151, 5186–5197. [Google Scholar] [PubMed]
- Yaqoob, P.; Newsholme, E.A.; Calder, P.C. The effect of dietary lipid manipulation on rat lymphocyte subsets and proliferation. Immunology 1994, 82, 603–610. [Google Scholar] [PubMed]
- Yaqoob, P.; Calder, P. Effects of dietary lipid manipulation upon inflammatory mediator production by murine macrophages. Cell. Immunol. 1995, 163, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Teague, H.; Fhaner, C.J.; Harris, M.; Duriancik, D.M.; Reid, G.E.; Shaikh, S.R. N-3 PUFAs enhance the frequency of murine B-cell subsets and restore the impairment of antibody production to a T-independent antigen in obesity. J. Lipid Res. 2013, 54, 3130–3138. [Google Scholar] [CrossRef] [PubMed]
- Gordon, T.; Kannel, W.B.; Castelli, W.P.; Dawber, T.R. Lipoproteins, cardiovascular disease, and death. The Framingham study. Arch. Intern. Med. 1981, 141, 1128–1131. [Google Scholar] [CrossRef] [PubMed]
- Rahilly-Tierney, C.R.; Lawler, E.V.; Scranton, R.E.; Michael Gaziano, J. Low-density lipoprotein reduction and magnitude of cardiovascular risk reduction. Prev. Cardiol. 2009, 12, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Wassall, S.R.; Stillwell, W. Docosahexaenoic acid domains: The ultimate non-raft membrane domain. Chem. Phys. Lipids 2008, 153, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Mullen, A.; Loscher, C.E.; Roche, H.M. Anti-inflammatory effects of EPA and DHA are dependent upon time and dose-response elements associated with LPS stimulation in THP-1-derived macrophages. J. Nutr. Biochem. 2010, 21, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Chaudhary, A.; Sethi, S. Oxidized omega-3 fatty acids inhibit NF-kappaB activation via a PPARalpha-dependent pathway. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1621–1627. [Google Scholar] [CrossRef] [PubMed]
- Svahn, S.L.; Grahnemo, L.; Palsdottir, V.; Nookaew, I.; Wendt, K.; Gabrielsson, B.; Schele, E.; Benrick, A.; Andersson, N.; Nilsson, S.; et al. Dietary polyunsaturated fatty acids increase survival and decrease bacterial load during septic Staphylococcus aureus infection and improve neutrophil function in mice. Infect. Immun. 2015, 83, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Hong, S.; Gronert, K.; Colgan, S.P.; Devchand, P.R.; Mirick, G.; Moussignac, R.L. Resolvins: A family of bioactive products of omega-3 fatty acid transformation circuits initiated by aspirin treatment that counter proinflammation signals. J. Exp. Med. 2002, 196, 1025–1037. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Takada, Y.; Boriek, A.M.; Aggarwal, B.B. Nuclear factor-kappaB: Its role in health and disease. J. Mol. Med. 2004, 82, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Shin, Y.J.; Cho, S.C.; Lee, S.M.; Bahn, Y.J.; Kim, J.Y.; Kwon, E.S.; Jeong do, Y.; Park, S.C.; Rhee, S.G.; et al. Peroxiredoxin 3 has a crucial role in the contractile function of skeletal muscle by regulating mitochondrial homeostasis. Free Radic Biol. Med. 2014, 77, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.Y.; Kim, Y.; Jeong, J.; Park, J.; Kim, I.; Huh, K.H.; Kim, Y.S.; Woo, H.A.; Rhee, S.G.; Lee, K.J.; et al. Peroxiredoxin 3 is a key molecule regulating adipocyte oxidative stress, mitochondrial biogenesis, and adipokine expression. Antioxid. Redox Signal. 2012, 16, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Polyunsaturated fatty acids and inflammatory processes: New twists in an old tale. Biochimie 2009, 91, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Vessby, B. Dietary fat, fatty acid composition in plasma and the metabolic syndrome. Curr. Opin. Lipidol. 2003, 14, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Brydon, E.W.; Morris, S.J.; Sweet, C. Role of apoptosis and cytokines in influenza virus morbidity. FEMS Microbiol. Rev. 2005, 29, 837–850. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Takahama, Y. XCL1 and XCR1 in the immune system. Microbes Infect. 2012, 14, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A. The chemokine system: Redundancy for robust outputs. Immunol. Today 1999, 20, 254–257. [Google Scholar] [CrossRef]
- Ren, M.; Zeng, J.; De Lemos-Chiarandini, C.; Rosenfeld, M.; Adesnik, M.; Sabatini, D.D. In its active form, the GTP-binding protein rab8 interacts with a stress-activated protein kinase. Proc. Natl. Acad. Sci. USA 1996, 93, 5151–5155. [Google Scholar] [CrossRef] [PubMed]
- Kushiyama, A.; hojima, N.; Ogihara, T.; Inukai, K.; Sakoda, H.; Fujishiro, M.; Fukushima, Y.; Anai, M.; Ono, H.; Horike, N.; et al. Resistin-like molecule beta activates MAPKs, suppresses insulin signaling in hepatocytes, and induces diabetes, hyperlipidemia, and fatty liver in transgenic mice on a high fat diet. J. Biol. Chem. 2005, 280, 42016–42025. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Zhong, J.; Xu, X.; Jordan, B.; Maurya, S.; Braunstein, Z.; Wang, T.Y.; Huang, W.; Aggarwal, S.; Periasamy, M.; et al. Exercise protects against diet-induced insulin resistance through downregulation of protein kinase Cbeta in mice. PLoS ONE 2013, 8, e81364. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Vienberg, S.G.; Bezy, O.; O’Neill, B.T.; Kahn, C.R. Role of PKCdelta in Insulin Sensitivity and Skeletal Muscle Metabolism. Diabetes 2015, 64, 4023–4032. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Efsa Panel on Dietetic Products, Nutrients and Allergies. Scientific Opinion on the substantiation of health claims related to eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA), docosapentaenoic acid (DPA) and maintenance of normal cardiac function (ID 504, 506, 516, 527, 538, 703, 1128, 1317, 1324, 1325), maintenance of normal blood glucose concentrations (ID 566), maintenance of normal blood pressure (ID 506, 516, 703, 1317, 1324), maintenance of normal blood HDL-cholesterol concentrations (ID 506), maintenance of normal (fasting) blood concentrations of triglycerides (ID 506, 527, 538, 1317, 1324, 1325), maintenance of normal blood LDL-cholesterol concentrations (ID 527, 538, 1317, 1325, 4689), protection of the skin from photo-oxidative (UV-induced) damage (ID 530), improved absorption of EPA and DHA (ID 522, 523), contribution to the normal function of the immune system by decreasing the levels of eicosanoids, arachidonic acid-derived mediators and pro-inflammatory cytokines (ID 520, 2914), and “immunomodulating agent” (4690) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2010, 8, 1796–1828. [Google Scholar]
- Efsa Panel on Dietetic Products, Nutrients and Allergies. Scientific Opinion on the Tolerable Upper Intake Level of eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA) and docosapentaenoic acid (DPA). EFSA J. 2012, 10, 2815. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Ingredient (g/100 g Diet) | Control | HFD-ED | HFD-CO | |
|---|---|---|---|---|
| Protein | Casein | 22.20 | 25.60 | 25.60 |
| Carbohydrates | Sucrose | 5.00 | 10.00 | 10.00 |
| Corn starch | 56.00 | 34.80 | 34.80 | |
| Cellulose | 5.00 | 5.80 | 5.80 | |
| Fat | Total | 5.00 | 15.00 | 15.00 |
| Corn oil | 2.50 | 3.00 | 5.00 | |
| Coconut oil | 2.50 | 10.00 | 10.00 | |
| EPAX oils a | 0.00 | 2.00 | 0.00 | |
| Minerals b | 2.00 | 2.50 | 2.50 | |
| Miconutrients c | 3.00 | 3.00 | 3.00 | |
| Choline bitartrate | 1.60 | 2.00 | 2.00 | |
| Cholesterol | 0.00 | 1.00 | 1.00 | |
| Methionine | 0.20 | 0.30 | 0.30 | |
| Energy content (kJ/100 g) | 1599 | 1752 | 1752 | |
| Protein E% | 24 | 25 | 25 | |
| Carbohydrate E% | 65 | 44 | 44 | |
| Fat E% | 12 | 32 | 32 | |
| Fatty acid composition d (mg/g diet) | ||||
| C10:0 | 0.20 | 1.47 | 1.33 | |
| C12:0 | 2.37 | 7.58 | 7.72 | |
| C14:0 | 1.54 | 4.58 | 4.78 | |
| C16:0 | 1.90 | 3.44 | 3.59 | |
| C18:0 | 0.68 | 2.26 | 2.49 | |
| SFA | 6.70 | 19.33 | 19.91 | |
| C18:1 n-9 | 2.82 | 4.80 | 5.26 | |
| MUFA | 2.82 | 4.80 | 5.26 | |
| C18:2 n-6 | 3.62 | 5.03 | 7.36 | |
| C18:3 n-6 | 0.12 | 0.22 | 0.26 | |
| Total n-6 PUFA | 3.74 | 5.26 | 7.62 | |
| C20:5 n-3 (EPA) | 0.00 | 2.03 | 0.01 | |
| C22:6 n-3 (DHA) | 0.00 | 4.58 | 0.01 | |
| Total n-3 PUFA | 0.00 | 6.61 | 0.02 | |
| Parameter | Control | HFD-ED | HFD-CO |
|---|---|---|---|
| Total number of animals; n | 9 | 12 | 12 |
| Initial body weight (g) | 27.50 ± 0.80 | 28.60 ± 0.80 | 24.30 ± 0.50 |
| Final body weight (g) | 31.40 ± 1.00 | 33.80 ± 0.80 | 36.20 ± 0.90 |
| Change in body weight (g) | 3.90 ± 0.50 a | 5.20 ± 0.30 a | 8.60 ± 0.50 b |
| Absolute spleen weight (g) | 0.09 ± 0.01 | 0.10 ± 0.00 | 0.09 ± 0.00 |
| Spleen/body weight ratio (g/100 g) | 0.29 ± 0.02 a,b | 0.29 ± 0.01 a | 0.24 ± 0.01 b |
| Neutral Lipids | Control n = 5 | HFD-ED n = 8 | HFD-CO n = 8 |
|---|---|---|---|
| C12:0 | 0.00 ± 0.00 | 0.01 ± 0.00 | 0.06 ± 0.02 |
| C14:0 | 0.01 ± 0.00 | 0.01 ± 0.00 a | 0.14 ± 0.06 b |
| C16:0 | 1.03 ± 0.20 | 1.46 ± 0.22 | 3.34 ± 1.05 |
| C18:0 | 0.21 ± 0.02 | 0.19 ± 0.04 | 0.13 ± 0.03 |
| C18:1 n-9 | 1.78 ± 0.46 | 1.05 ± 0.17 | 1.43 ± 0.33 |
| C18:2 n-6 | 0.03 ± 0.02 a | 0.2 ± 0.08 a | 0.86 ± 0.25 b |
| C20:3 n-6 | 0.11 ± 0.01 a | 0.03 ± 0.01 b | 0.05 ± 0.02 |
| C20:4 n-6 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.24 ± 0.14 |
| C22:6 n-3 | 0.00 ± 0.00 a | 0.16 ± 0.04 b | 0.00 ± 0.00 a |
| Total | 3.16 ± 0.58 | 3.12 ± 0.43 | 6.25 ± 1.37 |
| Free fatty acids | |||
| C12:0 | 0.01 ± 0.00 a | 0.02 ± 0.00 | 0.04 ± 0.01 b |
| C14:0 | 0.01 ± 0.00 | 0.02 ± 0.00 | 0.05 ± 0.01 |
| C16:0 | 0.20 ± 0.02 | 0.20 ± 0.01 | 0.35 ± 0.07 |
| C18:1 n-9 | 0.05 ± 0.00 | 0.05 ± 0.00 a | 0.06 ± 0.00 b |
| C18:3 n-3 | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.03 ± 0.00 |
| C20:3 n-6 | 0.06 ± 0.01 | 0.02 ± 0.00 a | 0.07 ± 0.01 b |
| C22:6 n-3 | 0.00 ± 0.00 a | 0.05 ± 0.00 b | 0.00 ± 0.00 a |
| Total | 0.35 ± 0.03 | 0.38 ± 0.02 | 0.60 ± 0.11 |
| Phospholipids | |||
| C12:0 | 0.00 ± 0.00 a | 0.01 ± 0.00 | 0.01 ± 0.00 b |
| C14:0 | 0.07 ± 0.00 a | 0.15 ± 0.01 b | 0.13 ± 0.00 c |
| C16:0 | 1.69 ± 0.05 | 1.70 ± 0.03 | 1.69 ± 0.05 |
| C18:0 | 0.61 ± 0.02 | 0.59 ± 0.01 | 0.67 ± 0.04 |
| C18:1 n-9 | 0.32 ± 0.01 a | 0.28 ± 0.01 b | 0.31 ± 0.01 a |
| C18:2 n-6 | 0.12 ± 0.00 a | 0.22 ± 0.02 b | 0.14 ± 0.01 a |
| C20:0 | 0.00 ± 0.00 | 0.01 ± 0.00 | 0.00 ± 0.00 |
| C20:1 n-7 | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.02 ± 0.00 |
| C20:2 n-6 | 0.02 ± 0.00 a | 0.05 ± 0.01 b | 0.03 ± 0.00 a |
| C20:3 n-6 | 0.17 ± 0.01 a | 0.18 ± 0.01 a | 0.25 ± 0.02 b |
| C20:4 n-6 | 4.41 ± 0.10 a | 1.92 ± 0.05 b | 4.33 ± 0.08 a |
| C20:5 n-3 | 0.00 ± 0.00 a | 0.41 ± 0.01 b | 0.00 ± 0.00 a |
| C22:0 | 0.02 ± 0.01 | 0.02 ± 0.00 | 0.01 ± 0.00 |
| C22:5 n-3 | 0.00 ± 0.00 a | 0.34 ± 0.01 b | 0.00 ± 0.00 a |
| C22:6 n-3 | 0.20 ± 0.02 a | 0.82 ± 0.02 b | 0.19 ± 0.01 a |
| Total | 7.67 ± 0.19 a | 6.71 ± 0.13 b | 7.78 ± 0.22 a |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soni, N.K.; Ross, A.B.; Scheers, N.; Savolainen, O.I.; Nookaew, I.; Gabrielsson, B.G.; Sandberg, A.-S. Splenic Immune Response Is Down-Regulated in C57BL/6J Mice Fed Eicosapentaenoic Acid and Docosahexaenoic Acid Enriched High Fat Diet. Nutrients 2017, 9, 50. https://doi.org/10.3390/nu9010050
Soni NK, Ross AB, Scheers N, Savolainen OI, Nookaew I, Gabrielsson BG, Sandberg A-S. Splenic Immune Response Is Down-Regulated in C57BL/6J Mice Fed Eicosapentaenoic Acid and Docosahexaenoic Acid Enriched High Fat Diet. Nutrients. 2017; 9(1):50. https://doi.org/10.3390/nu9010050
Chicago/Turabian StyleSoni, Nikul K., Alastair B. Ross, Nathalie Scheers, Otto I. Savolainen, Intawat Nookaew, Britt G. Gabrielsson, and Ann-Sofie Sandberg. 2017. "Splenic Immune Response Is Down-Regulated in C57BL/6J Mice Fed Eicosapentaenoic Acid and Docosahexaenoic Acid Enriched High Fat Diet" Nutrients 9, no. 1: 50. https://doi.org/10.3390/nu9010050