Live Faecalibacterium prausnitzii Does Not Enhance Epithelial Barrier Integrity in an Apical Anaerobic Co-Culture Model of the Large Intestine

1
Food Nutrition & Health Team, Food & Bio-Based Products Group, AgResearch Grasslands, Palmerston North 4442, New Zealand
2
Riddet Institute, Massey University, Palmerston North 4442, New Zealand
3
High-Value Nutrition National Science Challenge, Auckland 1142, New Zealand
*
Author to whom correspondence should be addressed.
Nutrients 2017, 9(12), 1349; https://doi.org/10.3390/nu9121349
Submission received: 27 October 2017
/
Revised: 29 November 2017
/
Accepted: 8 December 2017
/
Published: 12 December 2017
(This article belongs to the Special Issue Prebiotics and Probiotics)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Appropriate intestinal barrier maturation during infancy largely depends on colonization with commensal bacteria. Faecalibacterium prausnitzii is an abundant obligate anaerobe that colonizes during weaning and is thought to maintain colonic health throughout life. We previously showed that F. prausnitzii induced Toll-like receptor 2 (TLR2) activation, which is linked to enhanced tight junction formation. Therefore, we hypothesized that F. prausnitzii enhances barrier integrity, an important factor in appropriate intestinal barrier maturation. In order to test metabolically active bacteria, we used a novel apical anaerobic co-culture system that allows the survival of both obligate anaerobic bacteria and oxygen-requiring intestinal epithelial cells (Caco-2). The first aim was to optimize the culture medium to enable growth and active metabolism of F. prausnitzii while maintaining the viability and barrier integrity, as measured by trans-epithelial electrical resistance (TEER), of the Caco-2 cells. This was achieved by supplementing the apical cell culture medium with bacterial culture medium. The second aim was to test the effect of F. prausnitzii on TEER across Caco-2 cell layers. Live F. prausnitzii did not improve TEER, which indicates that its benefits are not via altering tight junction integrity. The optimization of the novel dual-environment co-culturing system performed in this research will enable the investigation of new probiotics originating from indigenous beneficial bacteria.
1. Introduction
Faecalibacterium prausnitzii is one of the most abundant bacterial species in the colon of healthy human adults [1,2]. This bacterium is thought to be critical for maintaining colonic health because its abundance is reduced in people with gastrointestinal diseases [3,4,5,6,7,8,9,10]. Therefore, increasing the abundance of F. prausnitzii in the colonic microbiota has become the target of much research, either by directly delivering the bacterium as a probiotic [11] or by using food ingredients that preferentially stimulate the growth of endogenous F. prausnitzii [12]. Despite this, little is known about the role of F. prausnitzii in appropriate development of the intestinal barrier during infancy and whether it has the potential to be a probiotic during early life.
Intestinal maturation, including the development of the intestinal barrier integrity and immune function as well as the establishment and stabilization of the microbiota, occurs throughout the first two years of life. Much of this process is regulated by diet (e.g., breast milk versus infant formula), which in turn influences the colonization patterns of the early microbiota and their interactions with the host [13]. F. prausnitzii colonizes the large intestine between six and 12 months of life [14,15,16], so it is likely to have an impact on intestinal maturation during weaning.
One key area of intestinal maturation is the education of the immune system by the resident bacteria. F. prausnitzii has been associated with anti-inflammatory effects in adult gnotobiotic rodents colonized with Escherichia coli [17] or Bacteroides thetaiotaomicron [18]. However, F. prausnitzii is unable to mono-colonize gnotobiotic rodents [17], which means that targeted in vivo studies are not possible. An alternative is to use in vitro techniques to investigate the specific immune-modulatory effects of F. prausnitzii on host cells. Such studies have been limited due to the difficulty of co-culturing obligate anaerobes and human oxygen-requiring cells using conventional culturing systems. Using a novel dual-environment co-culturing system we previously showed that live F. prausnitzii induced TLR2 activation in transfected human embryonic kidney cells (HEK293) [19], which has been implicated in maintaining homeostasis between immunity and tolerance in the intestinal epithelium [20].
Another key to appropriate intestinal maturation is development of the barrier integrity, which is crucial not only for nutrient absorption but also to prevent the entry of bacteria and food antigens from the lumen into underlying tissues [21]. F. prausnitzii improved barrier integrity in mice with DSS-induced colitis [22]. However, our previous study using Caco-2 cell monolayers as a model of the large intestinal epithelium showed that F. prausnitzii did not alter ion permeability, as measured by the trans-epithelial electrical resistance (TEER) assay, and increased small molecule permeability, as measured by the 3H-mannitol flux assay, which could be considered detrimental [23]. In the study described above using the dual-environment co-culturing system the viability of F. prausnitzii in apical anaerobic conditions was improved compared to when cultured in the presence of oxygen, but the bacterium was not actively growing. The discrepancy between the in vivo and in vitro results may be due to this lack of growth, especially since mammalian cells have been shown to respond differently to the same bacterium depending on its growth phases [24].
Therefore, the specific hypothesis of this research was that actively growing F. prausnitzii improves intestinal barrier integrity, as measured by the TEER across Caco-2 cells, indicating that it has potential to be a probiotic to improve intestinal barrier maturation during early life. In order to test the hypothesis the first aim of this study was to optimize the apical medium to suit the requirements of both the bacterium and the intestinal epithelial cells, and in particular to encourage growth and active metabolism of F. prausnitzii. The second aim was then to test the effects of three F. prausnitzii strains, A2-165, American Type Culture Collection (ATCC) 27768, and HTF-F, on TEER across Caco-2 cells to ensure that our results were not limited to one strain.
2. Materials and Methods
2.1. F. prausnitzii Culturing Conditions
The three F. prausnitzii strains A2-165 (DSM 17677), ATCC 27768, and HTF-F (DSM 26943) were kindly provided by Hermie J. M. Harmsen (Department of Medical Microbiology, University of Groningen, Groningen, The Netherlands). Bacteria were cultured anaerobically in Brain Heart Infusion (BHI) broth containing 3.7% (w/v) BHI powder (Becton Dickinson, Auckland, New Zealand) supplemented with 0.5% (w/v) yeast extract, 0.0005% (w/v) hemin, 0.0005% vitamin K and 0.2% l-cysteine (Sigma-Aldrich, Auckland, New Zealand) in an atmosphere of 10% CO2, 10% H2 in N2 at 37 °C (Concept Plus Anaerobic Workstation, Ruskinn Technology Ltd., Bridgend, UK) as previously described [19].
2.2. Caco-2 Cell Culturing Conditions
Caco-2 cells (HTB37) were obtained from the ATCC and used in experiments between passage 25 and 35. Caco-2 cells were maintained in cell culture flasks (Corning, New York, NY, USA) in Medium 199 (M199) cell culture medium supplemented with 10% fetal bovine serum (FBS), 1% non-essential amino acids (NEAA; 100× solution), and 1% penicillin-streptomycin (10,000 units/mL penicillin, and 10 mg/mL streptomycin (Life Technologies, Auckland, New Zealand), referred to as M199 Standard medium (M199 Std) and incubated at 37 °C in 5% CO2. Medium was replaced twice a week. For co-culture experiments Caco-2 cells were seeded on semi-permeable polyester membranes (Transwell inserts, 6.5 mm diameter, 0.4 μm pore size, Corning, New York, NY, USA) at a density of 8 × 104 cells per insert and cultured for 16–18 days to attain differentiated cell monolayers (TEER over 400 Ω·cm2).
2.3. Apical Anaerobic Co-Culture Model
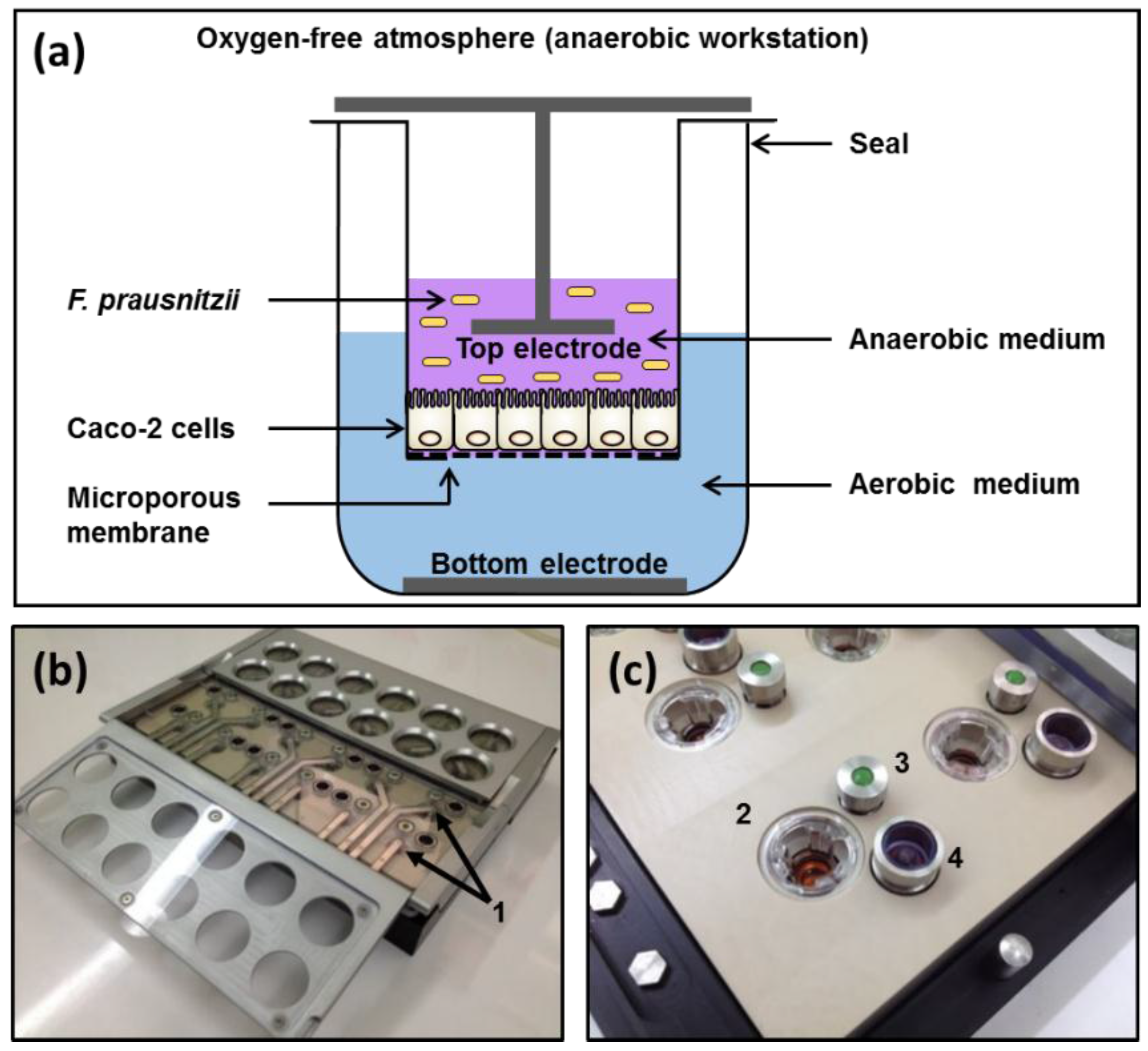
The apical anaerobic co-culture model utilized an in-house designed and built dual-environment co-culture chamber inside an anaerobic workstation (Figure 1). The Caco-2 cells received oxygen through the semi-permeable membrane from the oxygenated medium in the basal compartment and the bacterial cells were maintained in the apical anaerobic medium. The chamber was sterilized and consumables and solutions were made anaerobic before each experiment as previously described [19].
2.4. TEER Experiment Protocol
Twenty-four hours before the experiments, the apical and basal M199 Std was removed from the Caco-2 cell monolayers grown on Transwell inserts and replaced with M199 supplemented only with 10% FBS and 1% NEAAs, referred to as M199 TEER, in order to remove the antibiotics before co-culturing the cells with bacteria. On the day of the experiment, the basal wells of the co-culture chamber were filled with 3 mL of M199 TEER. The Transwell inserts containing the cell monolayers were carefully inserted into the co-culture chamber using a twisting motion. The co-culture chamber was transferred into the interlock chamber of the anaerobic workstation, purged with nitrogen and after finishing the interlock cycle moved into the anaerobic workstation. The co-culture chamber was connected to the commercially available CellZscope automated TEER monitoring system (CellZscope controller and CellZscope software version 2.2.3; nanoAnalytics, Münster, Germany). The TEER across each Caco-2 cell monolayer was measured twice and the second reading (after 1 h adaptation to the environment) was used as a baseline reading. The apical aerobic medium was removed and replaced with 260 μL of anaerobic medium or treatments. The TEER measurements were resumed and recorded hourly over 12 h. Since the initial TEER for each insert was different, the effect on TEER over time was expressed as the change in TEER compared to the initial TEER for each insert using the following Equation (1):
2.5. Survival of F. prausnitzii in Anaerobic Cell Culture Media
The three F. prausnitzii strains were incubated in M199 TEER medium to monitor their survival. Secondary bacterial cultures in stationary phase were pelleted by centrifugation at 2500× g for 6 min (11180/13190 rotor, Sigma 3-18K centrifuge, Osterode am Harz, Germany) and resuspended in anaerobic media inside the anaerobic workstation. Bacterial number was estimated using a Petroff-Hauser counting chamber and solutions diluted to yield a concentration of 2.4 × 107 bacterial cells/mL. This bacterial density was chosen so that when used in the co-culture experiments described below it resulted in a multiplicity of infection (MOI) of 100 bacterial cells per intestinal epithelial cell. Triplicates of the bacterial solutions were incubated at 37 °C and the optical density at a wavelength of 600 nm (OD600nm) was recorded (Implen OD600 DiluPhotometer with DC10 DiluCell cuvettes; Total Lab Solutions, Auckland, New Zealand) at 2-h intervals over 24 h.
2.6. TEER and Viability of Caco-2 Cells Using a Combination of Cell and Bacterial Culture Media
The effect of combining M199 TEER medium and BHI medium on the TEER and viability of Caco-2 cells was examined using the apical anaerobic co-culture model. Following the measurement of the initial resistance after two hours the apical medium was removed and replaced with 260 μL of anaerobic M199 TEER medium supplemented with increasing concentrations of anaerobic BHI (0%, 25%, 50%, 75%, or 100% BHI). After incubation for 12 h with automated hourly TEER measurements, the inserts were removed and the viability of the Caco-2 cells was determined using the neutral red uptake assay. Neutral red (3-amino-7-dimethylamino-2-methyl-phenazine hydrochloride; Sigma-Aldrich) was dissolved in phosphate-buffered saline (PBS) at 5 mg/mL, filter sterilized (0.22 μm filter), and diluted with M199 Std to a concentration of 50 μg/mL (referred to as neutral red medium). The medium was removed from all the Caco-2 cell monolayers and replenished with 200 μL neutral red medium. After incubation for 2 h at 37 °C in a CO2 incubator, the neutral red medium was removed and cell monolayers washed twice with PBS. The dye was extracted from viable cells by adding 200 μL solubilization solution (1% acetic acid–50% ethanol) and incubating at room temperature on a plate shaker at 200 rpm for 7 min. 150 μL of extract was transferred to a 96-well plate and the absorbance of neutral red was determined on a microplate reader at 540 nm (FlexStation 3, Molecular Devices, Sunnyvale, CA, USA). The background absorbance of the 96-well plate was measured at 690 nm and subtracted from the 540 nm measurement. The experiment was completed in five blocks with four replicates per treatment group in each block.
2.7. Viability of the F. prausnitzii Strains in the Apical Anaerobic Co-Culture Model Using a Combination of Cell and Bacterial Culture Media
Combinations of anaerobic M199 TEER medium and BHI medium were used for co-culturing Caco-2 cells and the three F. prausnitzii strains in the apical anaerobic co-culture model and it was determined if the bacteria were growing in these adapted conditions. The media compositions chosen were anaerobic M199 TEER supplemented with 25% or 50% of anaerobic BHI (referred to as 25% and 50% BHI). This adapted medium was only used for the apical compartment of the co-culture chamber; the basal compartments had aerobic M199 TEER medium. The bacterial suspensions were diluted with 25% and 50% BHI to a concentration of 2.4 × 107 bacterial cells/mL as described previously. The viability of the F. prausnitzii strains in the co-culture model was assessed by determining the viable colony forming units (CFU) of the bacteria at 0 h and 12 h after co-culture with the Caco-2 cells in both the cell supernatant and cell lysate. Duplicate ten-fold serial dilutions of the bacterial solutions were made in 96-well plates for each sample. Three 20-μL spots of each dilution were pipetted onto anaerobic BHI agar, allowed to dry, and then incubated anaerobically for 48 h at 37 °C. Spots with between 10 and 100 colonies were counted and the CFU were calculated. This experiment was done in five blocks, with three replicates per treatment group per block.
2.8. TEER Assay Using F. prausnitzii in Different Apical Media
To assess the effects of F. prausnitzii on TEER, each of the strains (A2-165, ATCC 27768, or HTF-F) was co-cultured with differentiated Caco-2 monolayers in the apical anaerobic co-culture model using 25% or 50% BHI as apical medium. There were eight treatment groups: the two control media (25% and 50% BHI) and the three F. prausnitzii strains diluted with the two different media. The TEER was recorded hourly for 12 h. The experiment was done in five blocks, with three replicates per treatment in each block.
2.9. TEER Assay Using Live and UV-Killed F. prausnitzii
The effect of both live and UV-killed F. prausnitzii on TEER across Caco-2 monolayers in the apical anaerobic co-culture model was determined using 50% BHI as the apical medium. The bacterial suspensions of the three F. prausnitzii strains were prepared for the co-culture experiments as follows. A 2-mL aliquot was removed from each bacterial suspension, and transferred to wells of a six-well plate. With the lid removed, the plate was placed on ice and the bacterial suspension treated with a UV lamp (UVP 3UV-38, Bio-Strategy Ltd., Auckland, New Zealand). The bacteria were exposed for 15 min to UVC light. The UV-treated bacteria were plated on anaerobic BHI agar plates and incubated anaerobically for 48 h at 37 °C to confirm that the bacteria were dead. For the TEER experiment there were seven treatment groups: the control medium (50% BHI), the three live F. prausnitzii strains and the three UV-killed F. prausnitzii strains. The experiment was done in three blocks, with three replicates per treatment per block.
2.10. Statistical Analysis
The statistical analyses were performed using SAS (SAS/STAT version 9.3; SAS Institute Inc., Cary, NC, USA). An analysis of repeated measures was conducted to test the effect of the treatment and time and their interaction on the response variables (change in TEER or change in OD600nm). The most appropriate covariance structure of the mixed model for each response variable was selected after fitting the models by restricted maximum likelihood method and comparing them using the log-likelihood ratio test. When an interaction was not significant it was removed from the model. An analysis of variance (ANOVA) was also conducted to test the effect of the BHI concentration on the viability of Caco-2 cells in apical anaerobic conditions. When the F-value of the analyses were significant (p < 0.05), the means were compared using Tukey tests. A two independent samples t-test procedure was performed to compare the viability of the three F. prausnitzii strains before and after the co-culture with Caco-2 cells in the apical anaerobic co-culture model. Additionally, a paired t-test was conducted to compare the viability of the three F. prausnitzii strains before and after the incubation in 50% BHI. For all the analyses the model assumptions (e.g., normal distribution and the homogeneity of variance) was evaluated using the Output Delivery System (ODS) graphics in SAS. When the response variable did not fulfil these assumptions a log10 transformation was performed to reach these assumptions.
3. Results and Discussion
3.1. F. prausnitzii Did Not Grow in an Anaerobic Cell Culture Medium
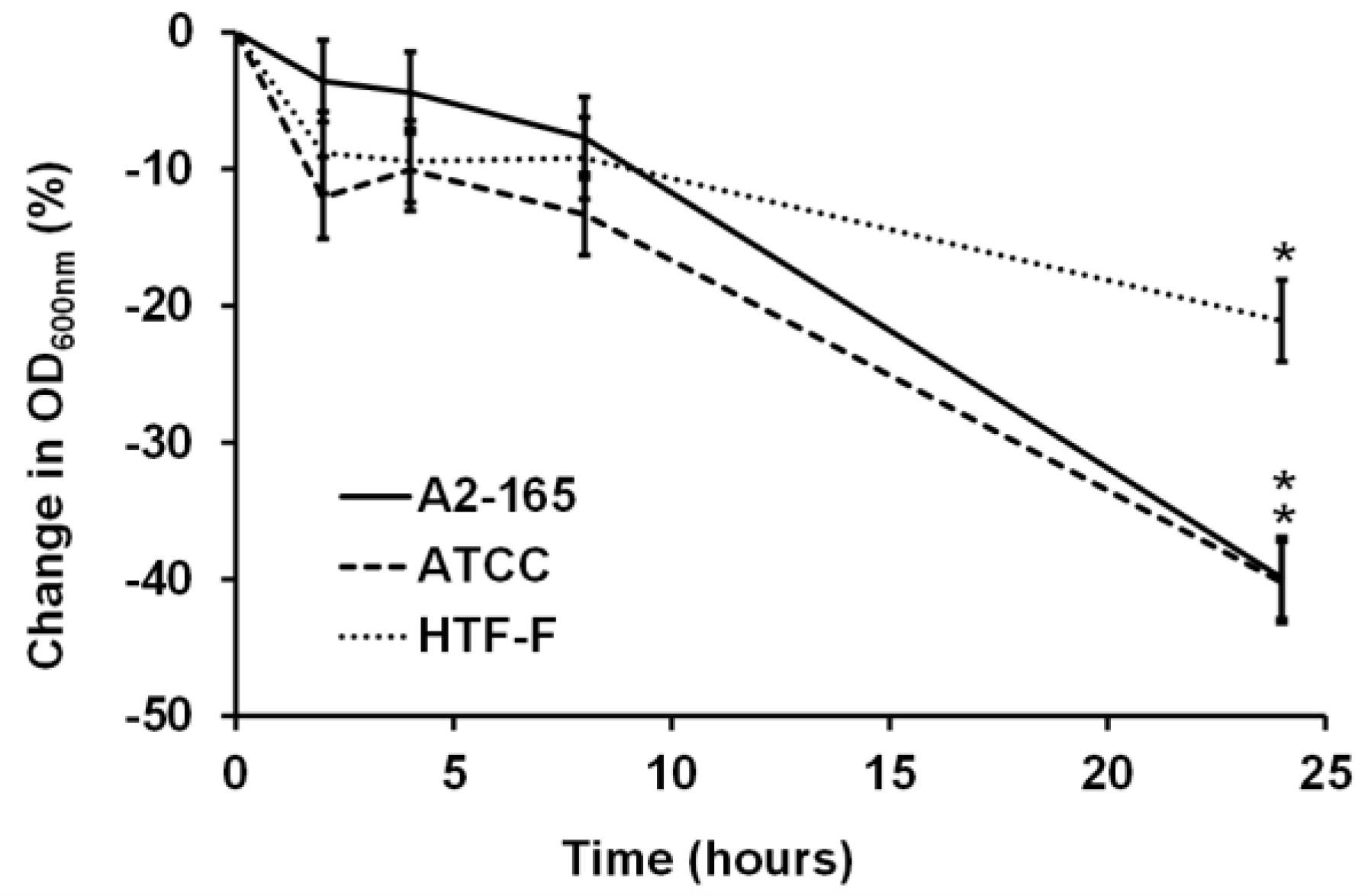
For bacterial–mammalian cell co-culture experiments, it is common to harvest bacterial cells by centrifugation and then resuspend the bacteria in mammalian cell culture media [23,25,26]. Studies have shown that there was no difference in the viability for strains of Escherichia coli, Salmonella typhimurium, and Lactobacillus fructosus in mammalian and bacterial culture medium [25]. In contrast to this, our results showed that the three F. prausnitzii strains did not grow in the anaerobic cell culture medium over the period of 24 h of incubation at 37 °C, as shown in the change in the OD600nm graph (Figure 2). There were no significant differences in the change of the OD600nm until 8 h of incubation for each of the three strains. However, after 24 h of incubation, all three strains showed a decrease in the OD600nm compared to the previous time points (0 to 8 h; p < 0.05). Therefore, it concluded it was necessary to supplement the cell culture medium to stimulate the growth and metabolic activity of F. prausnitzii.
When considering which supplements should be added to the medium first the nutrient composition of M199 TEER was investigated. Iron and vitamin K1 were identified as nutrients that may be lacking in this medium. Bacterial culture media often contain hemin as the iron source [1,23,27], whereas ferric nitrate is the only iron source in the M199 cell culture medium. This may not be optimum for F. prausnitzii since bacteria have developed heme acquisition systems to obtain iron from host heme-sequestering proteins [28]. Similarly, bacterial culture media often contain vitamin K1 [27,29], which bacteria convert into the vitamin K2 required for electron transport processes during anaerobic respiration [30]; whereas M199 only contains vitamin K3 (menadione), a synthetic type of vitamin K, which is a provitamin that requires conversion to menaquinone-4 in order to be active [23,31]. The bioavailability of vitamin K3 may therefore be lower for F. prausnitzii compared to other vitamin K sources.
Further investigation in the literature indicated that F. prausnitzii likely requires a complex medium to grow. A recent study used a combined approach of computational modeling, in vitro experiments, metabolomic analysis and genomic analysis to identify the metabolic capabilities of F. prausnitzii A2-165 and to develop a chemically defined medium, CDM1, for this bacterium [32]. However, CDM1 did not enable growth of F. prausnitzii A2-165, and even when enriched with additional vitamins, amino acids, and bases, F. prausnitzii A2-165 growth was still less than on the bacterial medium YCFAG [32]. Therefore, it was decided to supplement cell culture medium with a complex bacterial culture medium.
3.2. Bacterial Medium Did Not Affect Caco-2 Viability, but Reduced TEER at High Concentrations
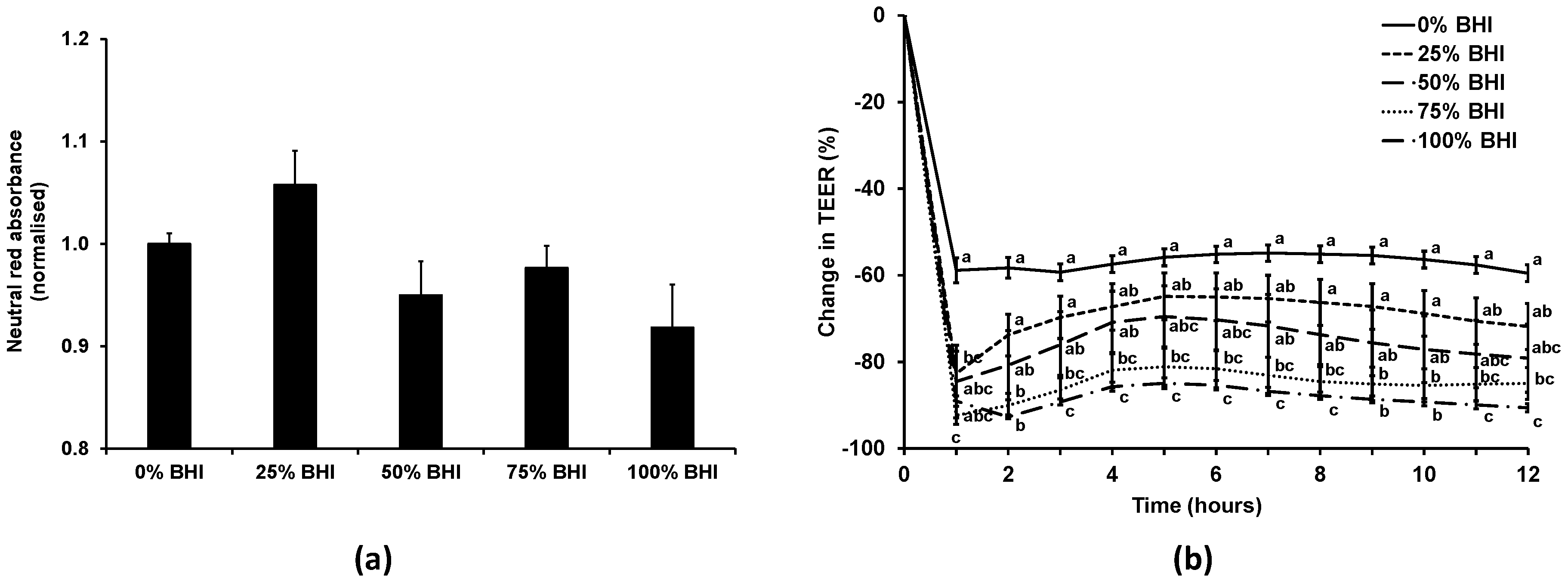
As our previous experience indicated that bacterial medium can be detrimental to mammalian cells, initially the effect of the bacterial medium on the Caco-2 cell monolayers was investigated. The viability of the Caco-2 cells treated with anaerobic M199 TEER mixed with anaerobic BHI in different ratios (0%, 25%, 50%, 75%, or 100% BHI) in apical anaerobic conditions (co-culture chamber inside the anaerobic workstation) over 12 h was determined using the neutral red viability assay (Figure 3a). There was no significant difference (p = 0.07) between the viability of Caco-2 cell monolayers treated with 0%, 25%, 50%, 75%, or 100% BHI in the apical anaerobic co-culture model.
The effect of the different M199 TEER-BHI media combinations on TEER across Caco-2 monolayers in apical anaerobic conditions was tested over 12 h (Figure 3b). There was a significant interaction between the time and the BHI concentration on the change in TEER (p < 0.01). When treated with 25% BHI, the normalized TEER across Caco-2 cell monolayers was significantly lower at one hour after adding the treatments compared to 0% BHI, however after that time point onwards there was no difference between 0% and 25% BHI. Caco-2 cell monolayers treated with 50% BHI showed no difference in TEER to Caco-2 cells treated with 0% BHI across all time points. In contrast, when 75% or 100% BHI were added to the apical side of Caco-2 cell monolayers, the normalized TEER values were significantly lower compared to cells treated with 0% BHI across all time points (p < 0.05).
The TEER of the Caco-2 cells treated with 50% BHI reached a plateau of approximately 300 Ω·cm2 following the initial drop in TEER, whereas those with cell culture medium had TEER values that plateaued at approximately 600 Ω·cm2. The Caco-2 cell monolayers treated with this combination of bacterial and cell culture medium had TEER values that were more comparable to those reported for human colon tissues (100 to 300 Ω·cm2) [33,34].
3.3. Two F. prausnitzii Strains Grew in 50% Bacterial Medium When Co-Cultured with Caco-2 Cells
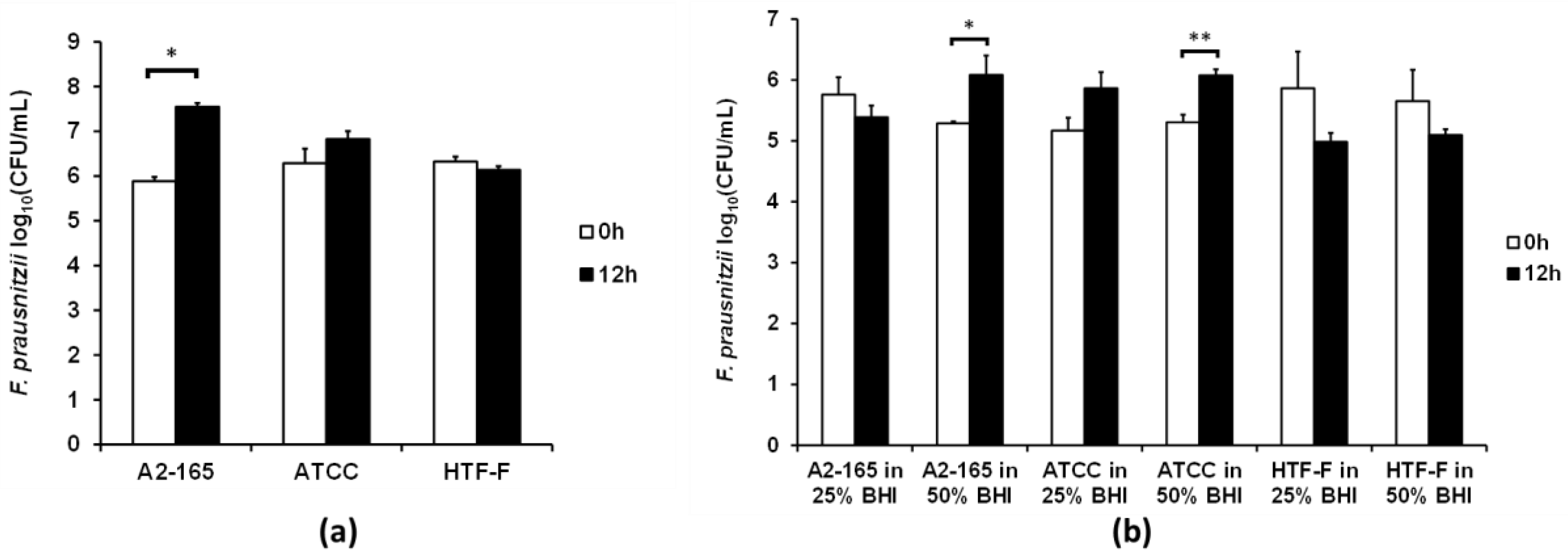
Based on the results above, the viability of the F. prausnitzii strains was tested using the anaerobic M199 TEER: BHI ratio (1:1), which was referred to as 50% BHI. The CFUs of the three F. prausnitzii strains were determined before and after their incubation in 50% BHI over 12 h (Figure 4a). The number of CFU of F. prausnitzii A2-165 increased by 1.7 log (p < 0.001), while the other two strains showed no significant differences between the CFU before and after 12 h of incubation in 50% BHI.
It was proposed that the F. prausnitzii strains may grow better in the presence of intestinal epithelial cells. Therefore the three strains were co-cultured with differentiated Caco-2 cell monolayers in the apical anaerobic co-culture model using M199 TEER supplemented with 25% or 50% BHI medium on the apical side of the cell monolayers. These medium compositions were chosen as they did not compromise the TEER and viability of the Caco-2 cell monolayers. The viability of the bacterial strains was determined by comparing the CFU before and after 12 h of co-culture with Caco-2 cells. No bacteria were able to be cultured from the Caco-2 cell lysate, therefore indicating that none of the three strains adhered to the Caco-2 cells. When 25% BHI was used at the apical side of the Caco-2 cell monolayer none of the three F. prausnitzii strains had an increase in CFU (Figure 4b). The number of CFU of both F. prausnitzii A2-165 and F. prausnitzii ATCC 27768 increased by 0.8 log after the 12 h of incubation (p < 0.05) when 50% BHI was used as the apical culture medium. However, there was no significant difference in CFU of F. prausnitzii HTF-F at 0 and 12 h of co-culture with Caco-2 cells when 50% BHI was used as the apical culture medium.
In agreement with published results using a simple dual-environment co-culture model [35], the presence of the Caco-2 cells improved the growth of the F. prausnitzii strains. This may be due to the presence of mucins. Though Caco-2 cells do not express mucin-2, the predominant mucin in the gastrointestinal tract, they express mucins 3 and 5A/C [36,37]. Furthermore, F. prausnitzii may benefit from the oxygen gradient close to the Caco-2 cell monolayer. F. prausnitzii uses an extracellular electron shuttle of flavins and thiols to transfer electrons to oxygen [38]. Small amounts of oxygen may diffuse from the aerobic basal compartment of the apical anaerobic co-culture model through the Caco-2 cell monolayer to the apical side. The F. prausnitzii strains may be able to use riboflavin (vitamin B2), one component of M199, for its extracellular electron transfer, which may benefit growth at this oxic–anoxic interphase [39].
3.4. Live F. prausnitzii Did Not Alter TEER across Caco-2 Cells
To determine whether live F. prausnitzii was able to improve TEER, differentiated Caco-2 monolayers were co-cultured with the three F. prausnitzii strains (A2-165, ATCC 27768, or HTF-F) in the apical anaerobic co-culture model using 25% or 50% BHI as apical medium. These apical media were chosen as one of them enabled growth of F. prausnitzii A2-165 and ATCC 27768 in co-culture with Caco-2 cells (50% BHI), whereas the other medium did not enable growth of any of the three strains (25% BHI) in the previous experiment. It could therefore be determined if change in TEER over time across Caco-2 monolayers differed when co-cultured with growing or non-growing bacteria. The interaction between the bacterial treatment and time was significant for both media (Figure 5; p < 0.001). However, there were no differences between the TEER of Caco-2 cells treated with either the 25% or 50% BHI medium or the three F. prausnitzii strains in the respective medium for each time point (p > 0.05).
Based on these results, it is likely that the maintenance of colonic health by F. prausnitzii is not mediated through enhancement of epithelial barrier integrity. Instead, the beneficial effects of F. prausnitzii may be due to it supporting immune homeostasis, as previously shown [19]. However, it is possible that metabolites secreted by live F. prausnitzii may require an increased treatment time than that undertaken here to exert their effects on intestinal barrier integrity. Specific probiotics and commensal bacteria influence intestinal barrier integrity through secreted metabolites [40,41,42], for example butyrate, a short chain fatty acid produced during bacterial fermentation, enhanced the barrier integrity through the regulation of tight junction assembly [43]. However, the barrier enhancing properties of butyrate occurred only after 24 h of incubation and TEER values reached maximum levels between 48 to 72 h [43]. In order to determine the effects of F. prausnitzii on TEER over a prolonged incubation time in this model, further validation studies would be necessary to ensure survival of the Caco-2 cells since the initial validation studies were performed for 12 h [23]. In addition, although F. prausnitzii is known as one of the major butyrate producers in the colon [1], it is unknown whether it produced butyrate when using 50% BHI as apical medium; therefore, further studies could analyze the composition of the apical medium after the co-culture of F. prausnitzii with Caco-2 cells.
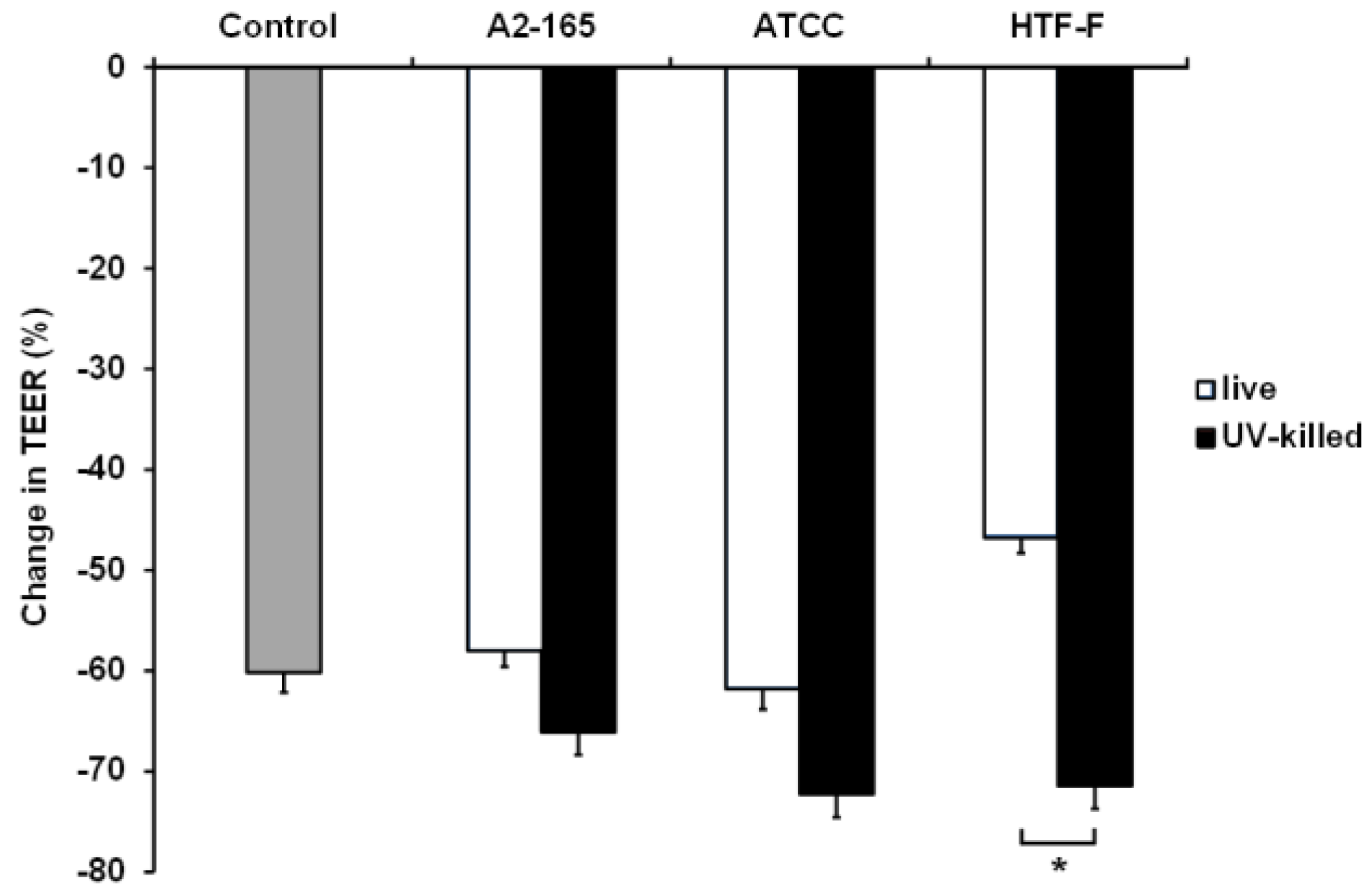
3.5. UV-Killed F. prausnitzii Decreased TEER
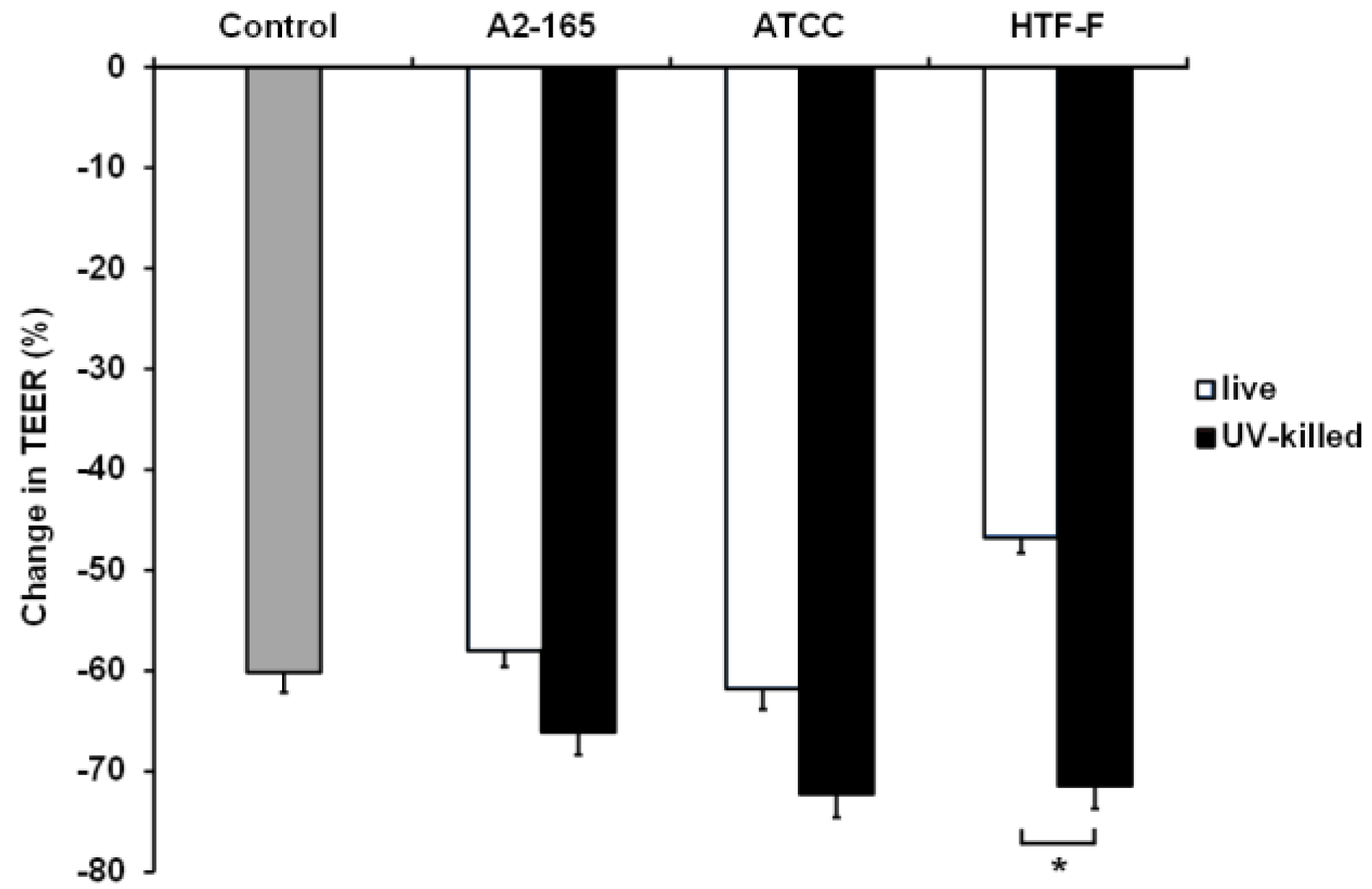
Caco-2 cells in the apical anaerobic co-culture model were co-cultured with live or UV-killed F. prausnitzii (strains A2-165, ATCC 27768, or HTF-F) in 50% BHI for 12 h (Figure 6). The interaction between the treatment and time was not significant, so it was removed from the statistical analysis. There was a significant treatment effect (p = 0.002), so between-treatment comparisons were warranted. No differences were observed between the TEER across Caco-2 monolayers exposed to the bacterial treatments and the untreated controls (p > 0.05). However, the TEER across Caco-2 monolayers co-cultured with live or UV-killed F. prausnitzii HTF-F was significantly different (p < 0.05) with higher TEER values recorded for cells treated with live bacteria. It is likely that this detrimental effect of the UV-killed bacteria is due to bacteria surface proteins interacting with the host cells. It is also possible that this negative effect is mitigated by metabolites produced by the live bacterium. Live F. prausnitzii may also maintain barrier integrity of Caco-2 monolayers through the activation of innate signaling. For example, commensal induced TLR2 signaling was shown to enhance intestinal barrier function and thereby limit mucosal inflammation [44,45]. We have previously shown that live F. prausnitzii induced higher TLR2 activation compared to dead F. prausnitzii [19], which may cause the barrier-protecting properties.
4. Conclusions
In conclusion, this research resulted in further optimization of the novel dual-environment co-culturing system, which will enable the investigation of new probiotics originating from indigenous beneficial bacteria. Contrary to our hypothesis, actively growing F. prausnitzii (strains A2-165, ATCC 27768, or HTF-F) did not improve intestinal barrier integrity, as measured by the TEER of Caco-2 cells. This result indicates that the benefits of F. prausnitzii are likely not due to it altering intracellular tight junction integrity.
Acknowledgments
This research was funded by the Marsden Fund, administered by the Royal Society of New Zealand (Grant AGR1102) and the New Zealand Ministry of Business, Innovation and Employment (Grant C10X1003 Dairy-based solutions for improved intestinal function). Eva Maier was supported by a Ph.D. stipend from the Riddet Institute, a New Zealand Centre of Research Excellence, funded by the Tertiary Education Commission. The authors also acknowledge the statistical advice of Carlos Montoya.
Author Contributions
Eva Maier, Rachel C. Anderson and Nicole C. Roy conceived and designed the experiments; Eva Maier performed the experiments and analyzed the data; Eva Maier and Rachel C. Anderson wrote the paper. All authors commented, edited, approved and are responsible for the final version of the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Duncan, S.H.; Hold, G.L.; Harmsen, H.J.M.; Stewart, C.S.; Flint, H.J. Growth requirements and fermentation products of Fusobacterium prausnitzii, and a proposal to reclassify it as Faecalibacterium prausnitzii gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 2141–2146. [Google Scholar] [PubMed]
- Hold, G.L.; Schwiertz, A.; Aminov, R.I.; Blaut, M.; Flint, H.J. Oligonucleotide probes that detect quantitatively significant groups of butyrate-producing bacteria in human feces. Appl. Environ. Microbiol. 2003, 69, 4320–4324. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Loening-Baucke, V.; Vaneechoutte, M.; Doerffel, Y. Active Crohn’s disease and ulcerative colitis can be specifically diagnosed and monitored based on the biostructure of the fecal flora. Inflamm. Bowel Dis. 2008, 14, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Seksik, P.; Furet, J.P.; Firmesse, O.; Nion-Larmurier, I.; Beaugerie, L.; Cosnes, J.; Corthier, G.; Marteau, P.; Doraé, J. Low counts of Faecalibacterium prausnitzii in colitis microbiota. Inflamm. Bowel Dis. 2009, 15, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Mondot, S.; Kang, S.; Furet, J.P.; Aguirre De Carcer, D.; McSweeney, C.; Morrison, M.; Marteau, P.; Doré, J.; Leclerc, M. Highlighting new phylogenetic specificities of Crohn’s disease microbiota. Inflamm. Bowel Dis. 2011, 17, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Kabeerdoss, J.; Sankaran, V.; Pugazhendhi, S.; Ramakrishna, B.S. Clostridium leptum group bacteria abundance and diversity in the fecal microbiota of patients with inflammatory bowel disease: A case-control study in India. BMC Gastroenterol. 2013, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, T.; Imaeda, H.; Takahashi, K.; Kasumi, E.; Bamba, S.; Fujiyama, Y.; Andoh, A. Decreased abundance of Faecalibacterium prausnitzii in the gut microbiota of Crohn’s disease. J. Gastroenterol. Hepatol. 2013, 28, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Rajilic-Stojanovic, M.; Biagi, E.; Heilig, H.G.; Kajander, K.; Kekkonen, R.A.; Tims, S.; de Vos, W.M. Global and deep molecular analysis of microbiota signatures in fecal samples from patients with irritable bowel syndrome. Gastroenterology 2011, 141, 1792–1801. [Google Scholar] [CrossRef] [PubMed]
- Balamurugan, R.; Rajendiran, E.; George, S.; Samuel, G.V.; Ramakrishna, B.S. Real-time polymerase chain reaction quantification of specific butyrate-producing bacteria, Desulfovibrio and Enterococcus faecalis in the feces of patients with colorectal cancer. J. Gastroenterol. Hepatol. 2008, 23, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- De Palma, G.; Nadal, I.; Medina, M.; Donat, E.; Ribes-Koninckx, C.; Calabuig, M.; Sanz, Y. Intestinal dysbiosis and reduced immunoglobulin-coated bacteria associated with coeliac disease in children. BMC Microbiol. 2010, 10, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, R.; Miquel, S.; Benevides, L.; Bridonneau, C.; Robert, V.; Hudault, S.; Chain, F.; Berteau, O.; Azevedo, V.; Chatel, J.M.; et al. Functional characterization of novel Faecalibacterium prausnitzii strains isolated from healthy volunteers: A step forward in the use of F. prausnitzii as a next-generation probiotic. Front. Microbiol. 2017, 8, 1226. [Google Scholar] [CrossRef] [PubMed]
- Blatchford, P.; Stoklosinski, H.; Walton, G.; Swann, J.; Gibson, G.; Gearry, R.; Ansell, J. Kiwifruit fermentation drives positive gut microbial and metabolic changes irrespective of initial microbiota composition. Bioact. Carbohydr. Diet. Fibre 2015, 6, 37–45. [Google Scholar] [CrossRef]
- Neu, J. Gastrointestinal maturation and implications for infant feeding. Early Hum. Dev. 2007, 83, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Talarico, S.T.; Santos, F.E.; Brandt, K.G.; Martinez, M.B.; Taddei, C.R. Anaerobic bacteria in the intestinal microbiota of Brazilian children. Clinics (Sao Paulo) 2017, 72, 154–160. [Google Scholar] [CrossRef]
- Hopkins, M.J.; Macfarlane, G.T.; Furrie, E.; Fite, A.; Macfarlane, S. Characterisation of intestinal bacteria in infant stools using real-time PCR and northern hybridisation analyses. FEMS Microbiol. Ecol. 2005, 54, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Pop, M.; Walker, A.W.; Paulson, J.; Lindsay, B.; Antonio, M.; Hossain, M.A.; Oundo, J.; Tamboura, B.; Mai, V.; Astrovskaya, I.; et al. Diarrhea in young children from low-income countries leads to large-scale alterations in intestinal microbiota composition. Genome Biol. 2014, 15, R76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miquel, S.; Leclerc, M.; Martin, R.; Chain, F.; Lenoir, M.; Raguideau, S.; Hudault, S.; Bridonneau, C.; Northen, T.; Bowen, B.; et al. Identification of metabolic signatures linked to anti-inflammatory effects of Faecalibacterium prausnitzii. MBio 2015, 6, e00300-15. [Google Scholar] [CrossRef] [PubMed]
- Wrzosek, L.; Miquel, S.; Noordine, M.-L.; Bouet, S.; Chevalier-Curt, M.J.; Robert, V.; Philippe, C.; Bridonneau, C.; Cherbuy, C.; Robbe-Masselot, C.; et al. Bacteroides thetaiotaomicron and Faecalibacterium prausnitzii influence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biol. 2013, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Maier, E.; Anderson, R.C.; Altermann, E.; Roy, N.C. Live Faecalibacterium prausnitzii induces greater TLR2 and TLR2/6 activation than the dead bacterium in an apical anaerobic co-culture system. Cell. Microbiol. 2017, e12805. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.M.; Loonen, L.M.; Karczewski, J.M. The role of innate signaling in the homeostasis of tolerance and immunity in the intestine. Int. J. Med. Microbiol. 2010, 300, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, A.H.; Yakymenko, O.; Olivier, I.; Håkansson, F.; Postma, E.; Keita, Å.V.; Söderholm, J.D. Faecalibacterium prausnitzii supernatant improves intestinal barrier function in mice DSS colitis. Scand. J. Gastroenterol. 2013, 48, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Ulluwishewa, D.; Anderson, R.C.; Young, W.; McNabb, W.C.; van Baarlen, P.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Live Faecalibacterium prausnitzii in an apical anaerobic model of the intestinal epithelial barrier. Cell. Microbiol. 2015, 17, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Haller, D.; Bode, C.; Hammes, W.P. Cytokine secretion by stimulated monocytes depends on the growth phase and heat treatment of bacteria: A comparative study between lactic acid bacteria and invasive pathogens. Microbiol. Immunol. 1999, 43, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Yuan, L.; Deng, J.; Yang, Q. Lactobacillus protects the integrity of intestinal epithelial barrier damaged by pathogenic bacteria. Front. Cell. Infect. Microbiol. 2015, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Young, W.; Clerens, S.; Cookson, A.L.; McCann, M.J.; Armstrong, K.M.; Roy, N.C. Human oral isolate Lactobacillus fermentum AGR1487 reduces intestinal barrier integrity by increasing the turnover of microtubules in Caco-2 cells. PLoS ONE 2013, 8, e78774. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, T.D.; Chalgren, S. Medium for use in antibiotic susceptibility testing of anaerobic bacteria. Antimicrob. Agents Chemother. 1976, 10, 926–928. [Google Scholar] [CrossRef] [PubMed]
- Wandersman, C.; Delepelaire, P. Bacterial iron sources: From siderophores to hemophores. Annu. Rev. Microbiol. 2004, 58, 611–647. [Google Scholar] [CrossRef] [PubMed]
- Bentley, R.; Meganathan, R. Biosynthesis of vitamin K (menaquinone) in bacteria. Microbiol. Rev. 1982, 46, 241–280. [Google Scholar] [PubMed]
- Kurosu, M.; Begari, E. Vitamin K2 in electron transport system: Are enzymes involved in vitamin K2 biosynthesis promising drug targets? Molecules 2010, 15, 1531. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Hirota, Y.; Sawada, N.; Yuge, N.; Watanabe, M.; Uchino, Y.; Okuda, N.; Shimomura, Y.; Suhara, Y.; Okano, T. Identification of UBIAD1 as a novel human menaquinone-4 biosynthetic enzyme. Nature 2010, 468, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Heinken, A.; Khan, M.T.; Paglia, G.; Rodionov, D.A.; Harmsen, H.J.M.; Thiele, I. Functional metabolic map of Faecalibacterium prausnitzii, a beneficial human gut microbe. J. Bacteriol. 2014, 196, 3289–3302. [Google Scholar] [CrossRef] [PubMed]
- Miyakawa, M.F.; Creydt, V.P.; Uzal, F.; McClane, B.; Ibarra, C. Clostridium perfringens enterotoxin damages the human intestine in vitro. Infect. Immun. 2005, 73, 8407–8410. [Google Scholar] [CrossRef] [PubMed]
- Soler, A.P.; Miller, R.D.; Laughlin, K.V.; Carp, N.Z.; Klurfeld, D.M.; Mullin, J.M. Increased tight junctional permeability is associated with the development of colon cancer. Carcinogenesis 1999, 20, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Sadaghian Sadabad, M.; von Martels, J.Z.H.; Khan, M.T.; Blokzijl, T.; Paglia, G.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N. A simple coculture system shows mutualism between anaerobic faecalibacteria and epithelial Caco-2 cells. Sci. Rep. 2015, 5, 17906. [Google Scholar] [CrossRef] [PubMed]
- Shekels, L.L.; Lyftogt, C.T.; Ho, S.B. Bile acid-induced alterations of mucin production in differentiated human colon cancer cell lines. Int. J. Biochem. Cell Biol. 1996, 28, 193–201. [Google Scholar] [CrossRef]
- Klinken, B.J.-W.; Oussoren, E.; Weenink, J.-J.; Strous, G.J.; Büller, H.A.; Dekker, J.; Einerhand, A.W.C. The human intestinal cell lines Caco-2 and LS174T as models to study cell-type specific mucin expression. Glycoconj. J. 1996, 13, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.; Duncan, S.H.; Stams, A.J.M.; van Dijl, J.M.; Flint, H.J.; Harmsen, H.J.M. The gut anaerobe Faecalibacterium prausnitzii uses an extracellular electron shuttle to grow at oxic-anoxic interphases. ISME J. 2012, 6, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.; Browne, W.R.; Van Dijl, J.M.; Harmsen, H.J.M. How can Faecalibacterium prausnitzii employ riboflavin for extracellular electron transfer? Antioxid. Redox Signal. 2012, 17, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Ewaschuk, J.B.; Diaz, H.; Meddings, L.; Diederichs, B.; Dmytrash, A.; Backer, J.; Looijer-van Langen, M.; Madsen, K.L. Secreted bioactive factors from Bifidobacterium infantis enhance epithelial cell barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1025–G1034. [Google Scholar] [CrossRef] [PubMed]
- Segawa, S.; Fujiya, M.; Konishi, H.; Ueno, N.; Kobayashi, N.; Shigyo, T.; Kohgo, Y. Probiotic-derived polyphosphate enhances the epithelial barrier function and maintains intestinal homeostasis through integrin–p38 MAPK pathway. PLoS ONE 2011, 6, e23278. [Google Scholar] [CrossRef] [PubMed]
- Otte, J.-M.; Podolsky, D.K. Functional modulation of enterocytes by gram-positive and gram-negative microorganisms. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G613–G626. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.-J.M.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [PubMed]
- Cario, E.; Gerken, G.; Podolsky, D.K. Toll-like receptor 2 controls mucosal inflammation by regulating epithelial barrier function. Gastroenterology 2007, 132, 1359–1374. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Apical anaerobic co-culture model. (a) Schematic diagram of a single well of the apical anaerobic co-culture model used for the co-culture of Caco-2 cell monolayers with anaerobic F. prausnitzii. The top and bottom electrodes enable the determination of the effect of F. prausnitzii on TEER across the Caco-2 cell monolayers; (b,c) Photographs of the co-culture chamber including details of the components. 1: Top electrodes; 2: Transwell insert containing Caco-2 cell monolayer; 3: Septum for basal media sampling; 4: One-way pressure relief valve.
Figure 1.
Apical anaerobic co-culture model. (a) Schematic diagram of a single well of the apical anaerobic co-culture model used for the co-culture of Caco-2 cell monolayers with anaerobic F. prausnitzii. The top and bottom electrodes enable the determination of the effect of F. prausnitzii on TEER across the Caco-2 cell monolayers; (b,c) Photographs of the co-culture chamber including details of the components. 1: Top electrodes; 2: Transwell insert containing Caco-2 cell monolayer; 3: Septum for basal media sampling; 4: One-way pressure relief valve.
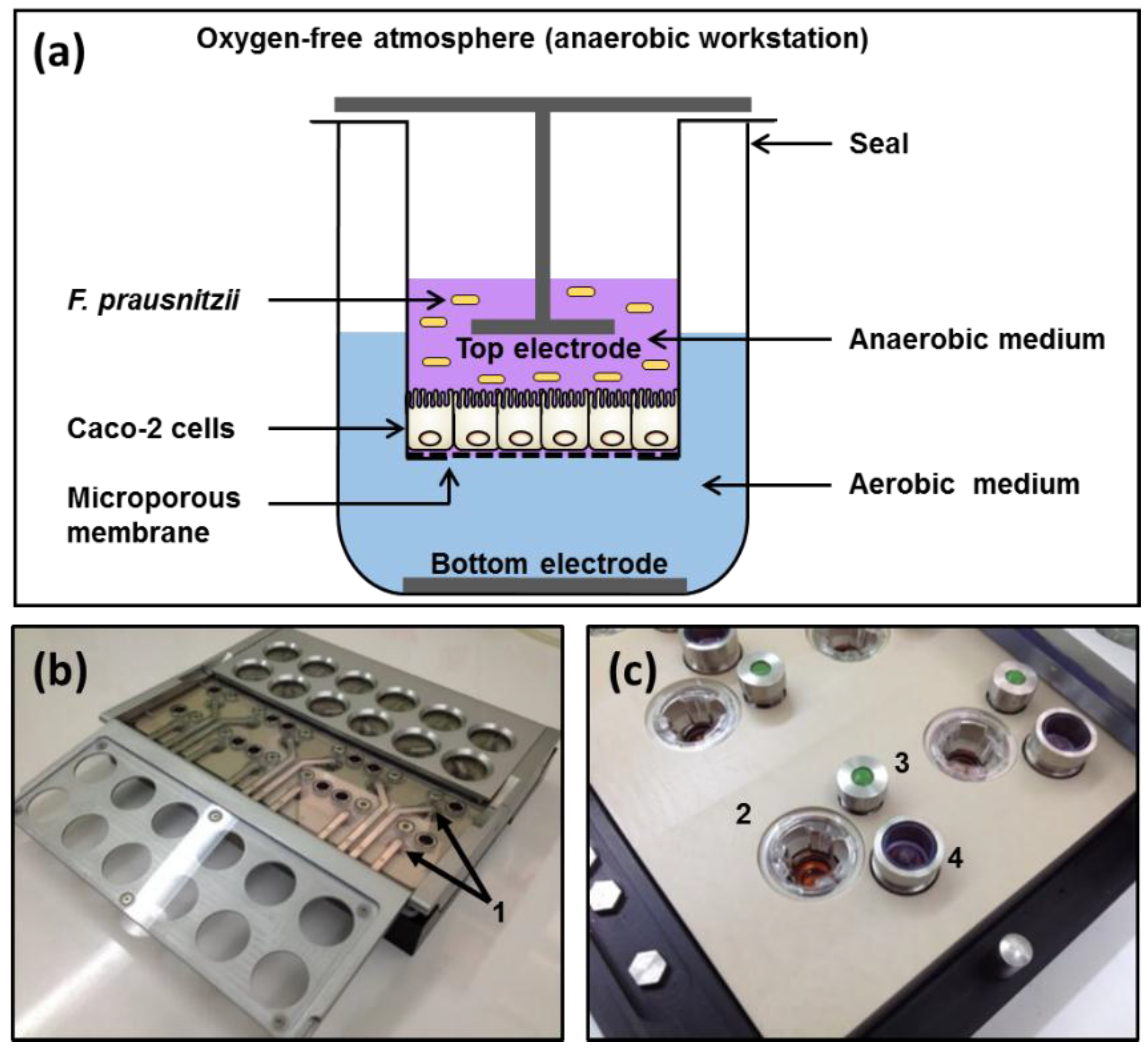
Figure 2.
Normalized change in OD600nm (%) of the three F. prausnitzii strains in an anaerobic cell culture medium. The three F. prausnitzii strains (A2-165, ATCC 27768, and HTF-F) in stationary phase were resuspended in anaerobic M199 TEER, incubated at 37 °C, and the OD600nm was measured over 24 h. The graph shows the mean values (±SEM; n = 3) after normalizing by the OD600nm at time 0 h for each of the three F. prausnitzii strains over 24 h. * Change in OD600nm different compared to previous time point for the same strain (p < 0.05).
Figure 2.
Normalized change in OD600nm (%) of the three F. prausnitzii strains in an anaerobic cell culture medium. The three F. prausnitzii strains (A2-165, ATCC 27768, and HTF-F) in stationary phase were resuspended in anaerobic M199 TEER, incubated at 37 °C, and the OD600nm was measured over 24 h. The graph shows the mean values (±SEM; n = 3) after normalizing by the OD600nm at time 0 h for each of the three F. prausnitzii strains over 24 h. * Change in OD600nm different compared to previous time point for the same strain (p < 0.05).
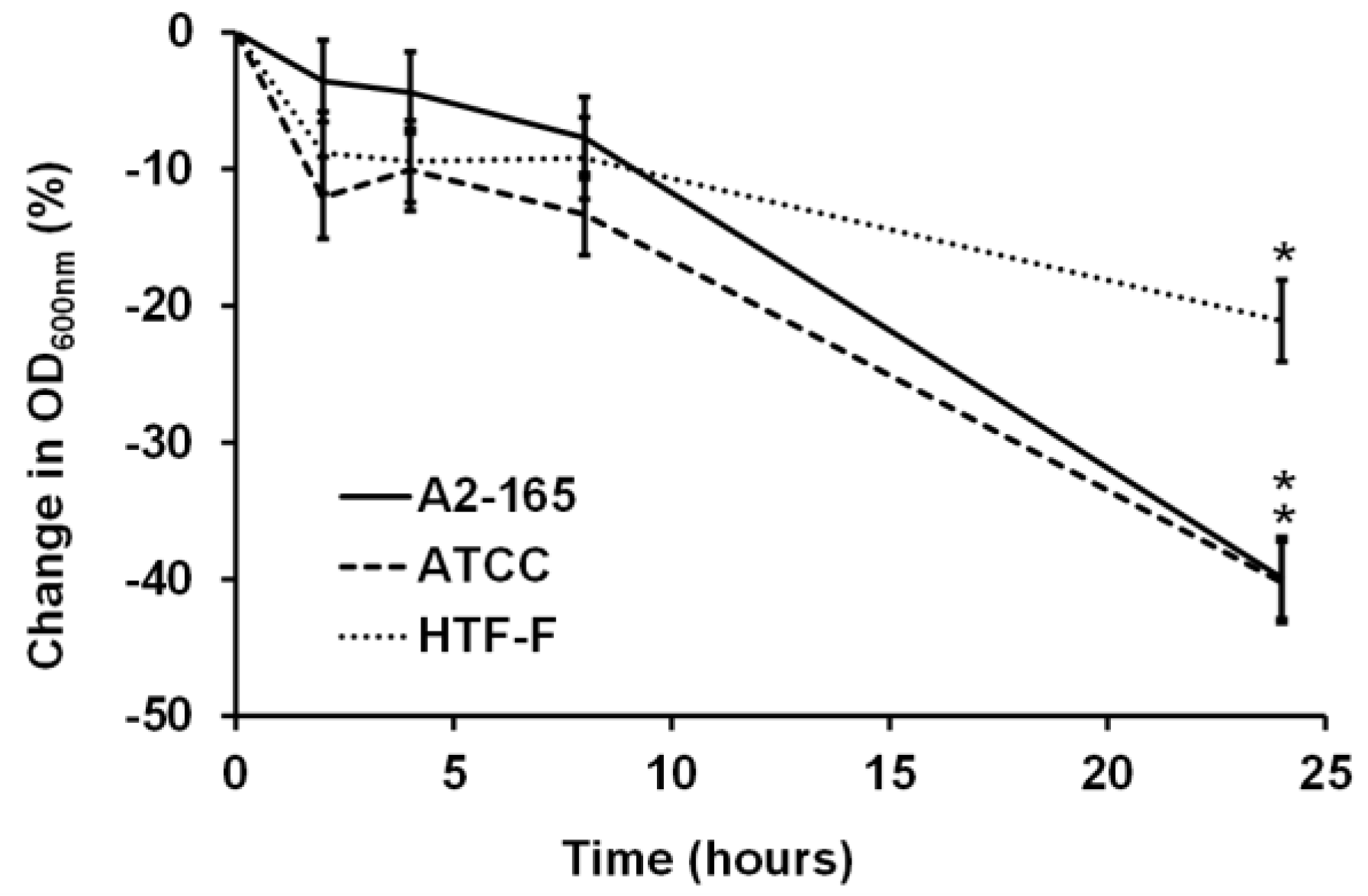
Figure 3.
Effect of combining cell and bacterial culture medium on the viability and TEER of Caco-2 cells in the apical anaerobic co-culture model. The apical compartment of Caco-2 cell monolayers were exposed to non-supplemented anaerobic M199 TEER medium (0% BHI), or M199 TEER medium supplemented with increasing concentrations of anaerobic BHI medium (25%, 50%, 75%, or 100% BHI). (a) Viability (mean ± SEM; n = 20) of Caco-2 cells after 12 h incubation. Neutral red absorbance was normalized by adjusting the 0% BHI exposed cells to 1. Viability of the Caco-2 cells was unchanged as a consequence of differing culture medium composition (p = 0.07). (b) Mean (±SEM; n = 20) change in TEER as a percentage of initial TEER across Caco-2 cell monolayers over 12 h for each medium. There was a significant interaction between the time and the BHI concentration on the change in TEER (p < 0.01). Treatments that do not share the same letters are significantly different (p < 0.05).
Figure 3.
Effect of combining cell and bacterial culture medium on the viability and TEER of Caco-2 cells in the apical anaerobic co-culture model. The apical compartment of Caco-2 cell monolayers were exposed to non-supplemented anaerobic M199 TEER medium (0% BHI), or M199 TEER medium supplemented with increasing concentrations of anaerobic BHI medium (25%, 50%, 75%, or 100% BHI). (a) Viability (mean ± SEM; n = 20) of Caco-2 cells after 12 h incubation. Neutral red absorbance was normalized by adjusting the 0% BHI exposed cells to 1. Viability of the Caco-2 cells was unchanged as a consequence of differing culture medium composition (p = 0.07). (b) Mean (±SEM; n = 20) change in TEER as a percentage of initial TEER across Caco-2 cell monolayers over 12 h for each medium. There was a significant interaction between the time and the BHI concentration on the change in TEER (p < 0.01). Treatments that do not share the same letters are significantly different (p < 0.05).
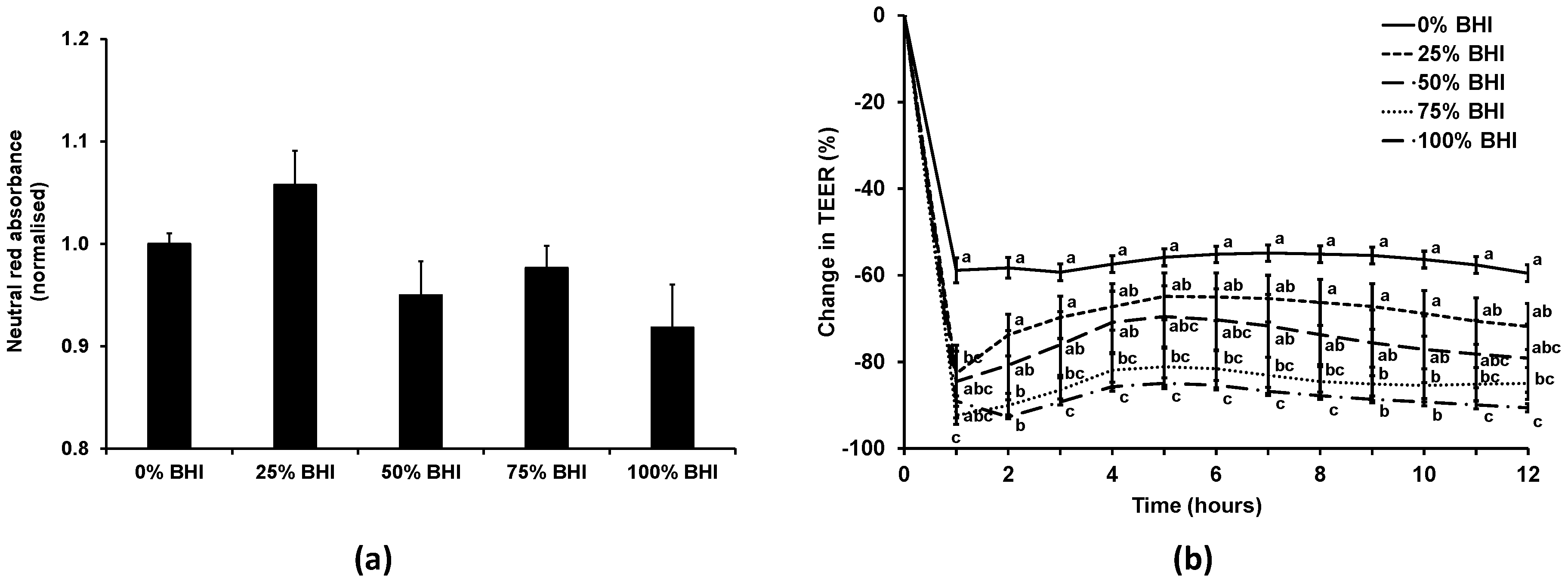
Figure 4.
Viability of the three F. prausnitzii strains in a cell culture medium supplemented with bacterial culture medium. (a) In Hungate culture tubes. The three F. prausnitzii strains in stationary phase were resuspended in anaerobic M199 TEER: BHI (1:1) (referred to as 50% BHI) and incubated anaerobically at 37 °C. The graph shows the mean (±SEM; n = 3) log10(CFU/mL) of the three F. prausnitzii strains before and after 12 h of incubation in 50% BHI. * Mean log10(CFU/mL) differ between 0 and 12 h at p < 0.05. (b) In the apical anaerobic co-culture model with Caco-2 cells. The three F. prausnitzii strains were co-cultured with Caco-2 cells using two different media on the apical side of the Caco-2 cell monolayer (25% and 50% BHI). The graph shows the mean (±SEM; n = 4 and 8 for time 0 and 12 h, respectively) log10(CFU/mL) of the bacteria at 0 and 12 h of incubation with Caco-2 cells in the co-culture model. Mean log10(CFU/mL) differ between 0 and 12 h at * p < 0.05 and ** p < 0.01.
Figure 4.
Viability of the three F. prausnitzii strains in a cell culture medium supplemented with bacterial culture medium. (a) In Hungate culture tubes. The three F. prausnitzii strains in stationary phase were resuspended in anaerobic M199 TEER: BHI (1:1) (referred to as 50% BHI) and incubated anaerobically at 37 °C. The graph shows the mean (±SEM; n = 3) log10(CFU/mL) of the three F. prausnitzii strains before and after 12 h of incubation in 50% BHI. * Mean log10(CFU/mL) differ between 0 and 12 h at p < 0.05. (b) In the apical anaerobic co-culture model with Caco-2 cells. The three F. prausnitzii strains were co-cultured with Caco-2 cells using two different media on the apical side of the Caco-2 cell monolayer (25% and 50% BHI). The graph shows the mean (±SEM; n = 4 and 8 for time 0 and 12 h, respectively) log10(CFU/mL) of the bacteria at 0 and 12 h of incubation with Caco-2 cells in the co-culture model. Mean log10(CFU/mL) differ between 0 and 12 h at * p < 0.05 and ** p < 0.01.
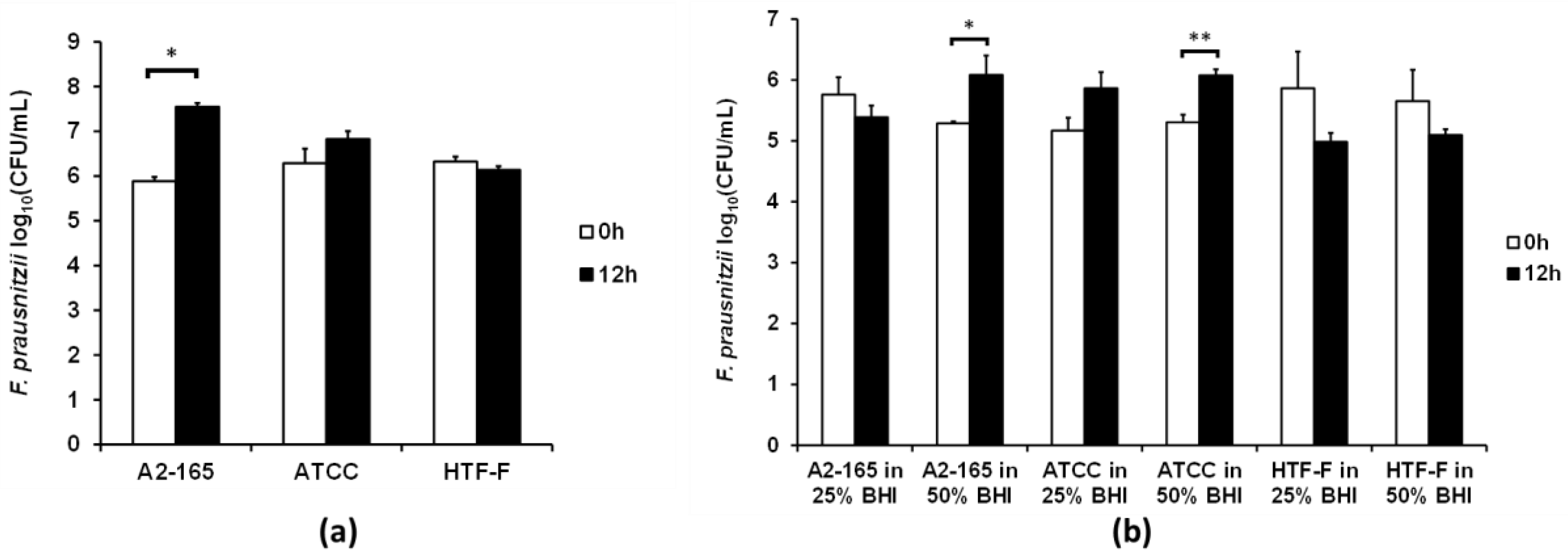
Figure 5.
Change in TEER across Caco-2 monolayers co-cultured with live F. prausnitzii using 25% and 50% BHI as apical medium. Differentiated Caco-2 monolayers were co-cultured with the three F. prausnitzii strains for 12 h in the apical anaerobic co-culture model using 25% or 50% BHI as apical medium. The interaction between the treatment and time was significant for the change in TEER (p < 0.001). The graphs show the mean (±SEM; n = 12) change in TEER across Caco-2 monolayers when using (a) 25% BHI and (b) 50% BHI as apical medium. No differences were determined between the two control media and the three F. prausnitzii strains in the respective medium at any time point (p > 0.05).
Figure 5.
Change in TEER across Caco-2 monolayers co-cultured with live F. prausnitzii using 25% and 50% BHI as apical medium. Differentiated Caco-2 monolayers were co-cultured with the three F. prausnitzii strains for 12 h in the apical anaerobic co-culture model using 25% or 50% BHI as apical medium. The interaction between the treatment and time was significant for the change in TEER (p < 0.001). The graphs show the mean (±SEM; n = 12) change in TEER across Caco-2 monolayers when using (a) 25% BHI and (b) 50% BHI as apical medium. No differences were determined between the two control media and the three F. prausnitzii strains in the respective medium at any time point (p > 0.05).

Figure 6.
Change in TEER across Caco-2 monolayers co-cultured with live or UV-killed F. prausnitzii using 50% BHI as apical medium. Caco-2 monolayers were co-cultured with live or UV-killed F. prausnitzii (strains A2-165, ATCC 27768, or HTF-F). The TEER across the Caco-2 monolayers was recorded hourly over 12 h. The interaction between the treatment and time was not significant and so was removed from the statistical model. There was a significant treatment effect on the change in TEER (p = 0.002). The graph shows the mean (±SEM; n = 9) change in TEER across Caco-2 monolayers co-cultured with live or UV-killed F. prausnitzii. * indicates significant difference in TEER (p < 0.05).
Figure 6.
Change in TEER across Caco-2 monolayers co-cultured with live or UV-killed F. prausnitzii using 50% BHI as apical medium. Caco-2 monolayers were co-cultured with live or UV-killed F. prausnitzii (strains A2-165, ATCC 27768, or HTF-F). The TEER across the Caco-2 monolayers was recorded hourly over 12 h. The interaction between the treatment and time was not significant and so was removed from the statistical model. There was a significant treatment effect on the change in TEER (p = 0.002). The graph shows the mean (±SEM; n = 9) change in TEER across Caco-2 monolayers co-cultured with live or UV-killed F. prausnitzii. * indicates significant difference in TEER (p < 0.05).
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Maier, E.; Anderson, R.C.; Roy, N.C. Live Faecalibacterium prausnitzii Does Not Enhance Epithelial Barrier Integrity in an Apical Anaerobic Co-Culture Model of the Large Intestine. Nutrients 2017, 9, 1349. https://doi.org/10.3390/nu9121349
AMA Style
Maier E, Anderson RC, Roy NC. Live Faecalibacterium prausnitzii Does Not Enhance Epithelial Barrier Integrity in an Apical Anaerobic Co-Culture Model of the Large Intestine. Nutrients. 2017; 9(12):1349. https://doi.org/10.3390/nu9121349
Chicago/Turabian StyleMaier, Eva, Rachel C. Anderson, and Nicole C. Roy. 2017. "Live Faecalibacterium prausnitzii Does Not Enhance Epithelial Barrier Integrity in an Apical Anaerobic Co-Culture Model of the Large Intestine" Nutrients 9, no. 12: 1349. https://doi.org/10.3390/nu9121349
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.