An Exercise‐Only Intervention in Obese Fathers Restores Glucose and Insulin Regulation in Conjunction with the Rescue of Pancreatic Islet Cell Morphology and MicroRNA Expression in Male Offspring

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Founder Animals and Diet
2.2. Exercise Intervention (Swimming)
2.3. Natural Mating to Produce Male Offspring
2.4. Body Composition
2.5. Glucose and Insulin Tolerance
2.6. Male Offspring Insulin Secretion
2.7. Metabolites and Hormone Analysis
2.8. Male Offspring Pancreatic Histology and Islet Cell Morphology
2.9. Male Offspring Pancreas MicroRNA Analysis
2.10. Statistics
3. Results
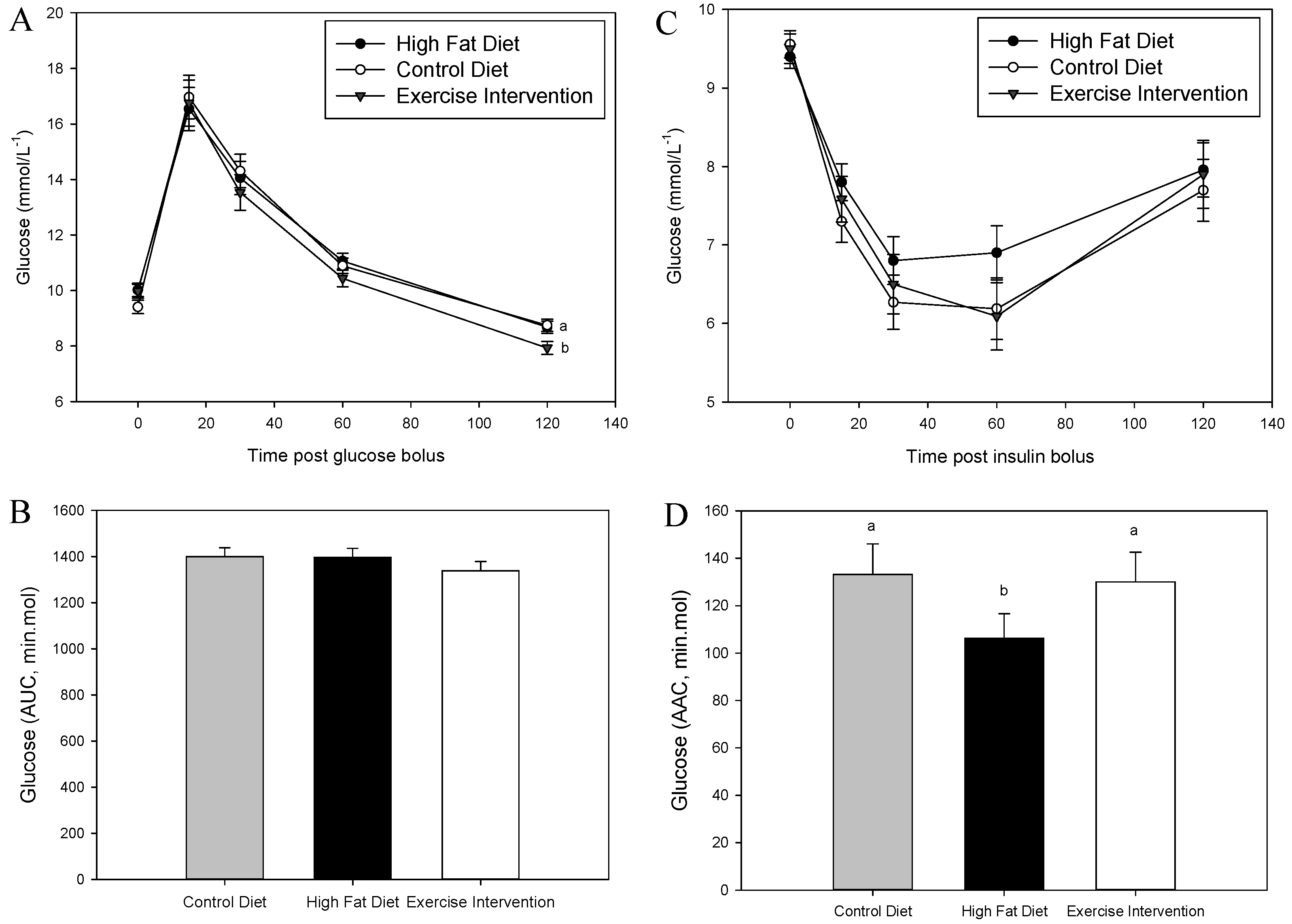
3.1. Founder Male Phenotype
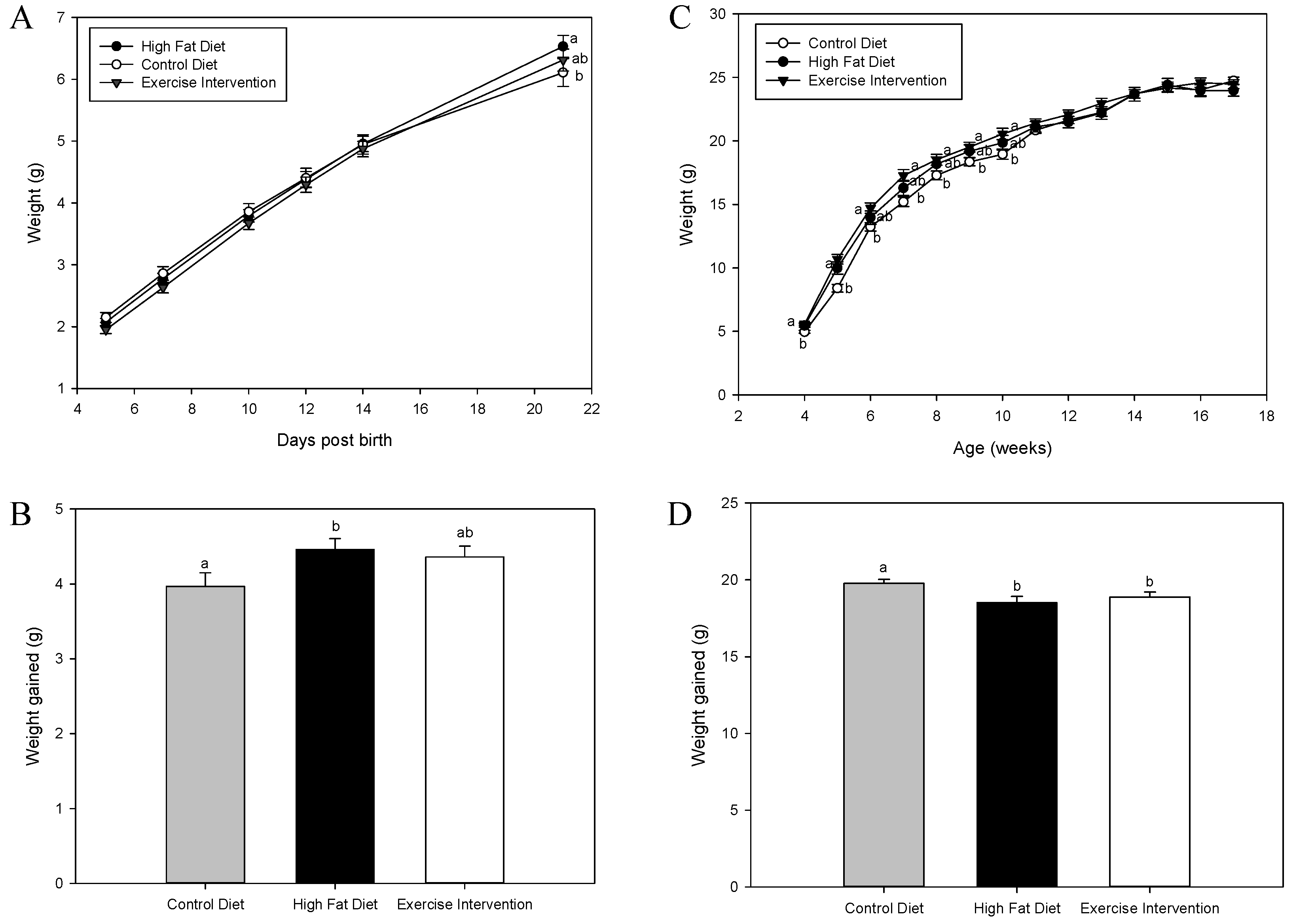
3.2. Exercise Interventions in Fathers Partially Restore Pre-Weaning Weights in Male Offspring
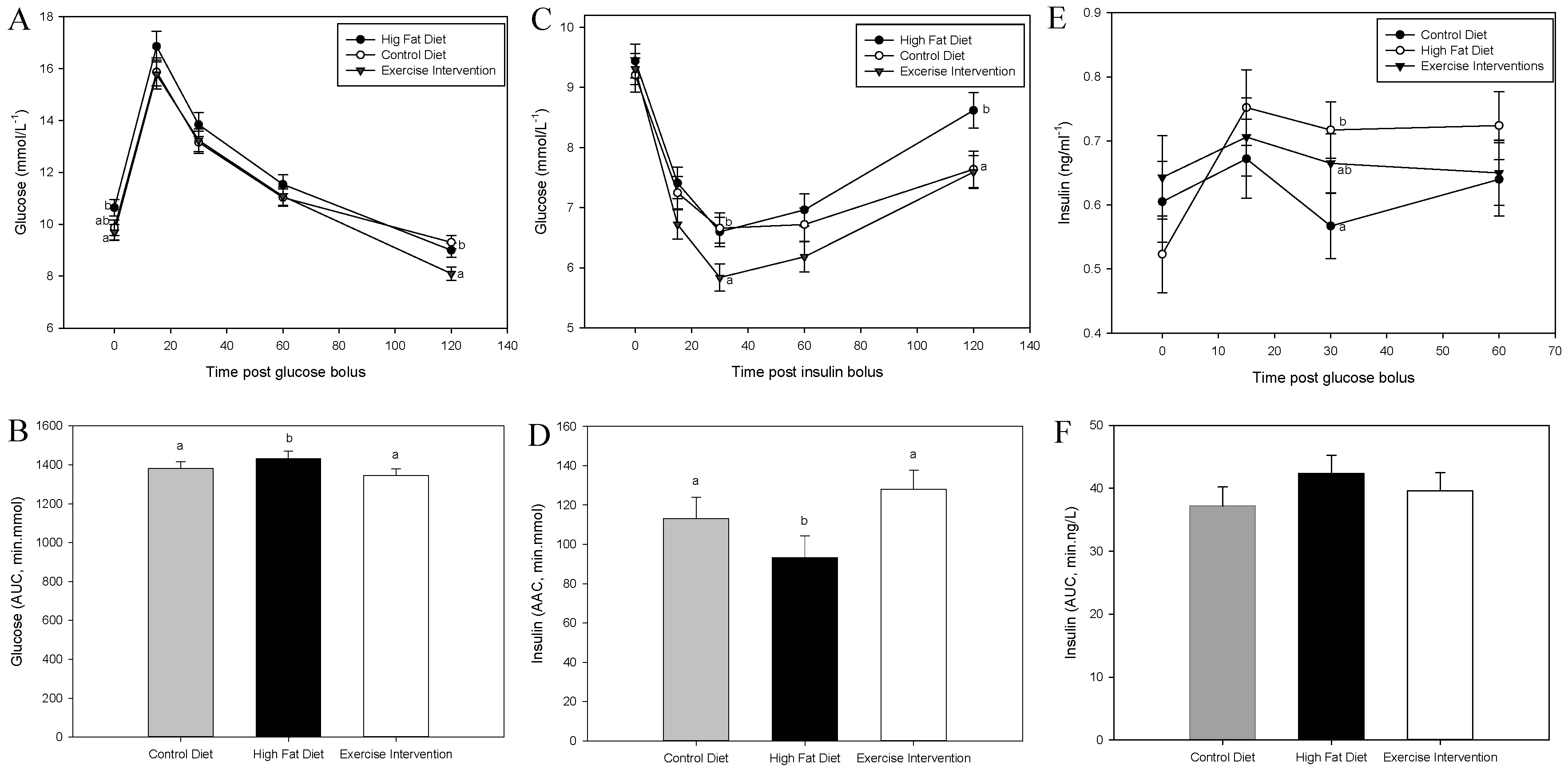
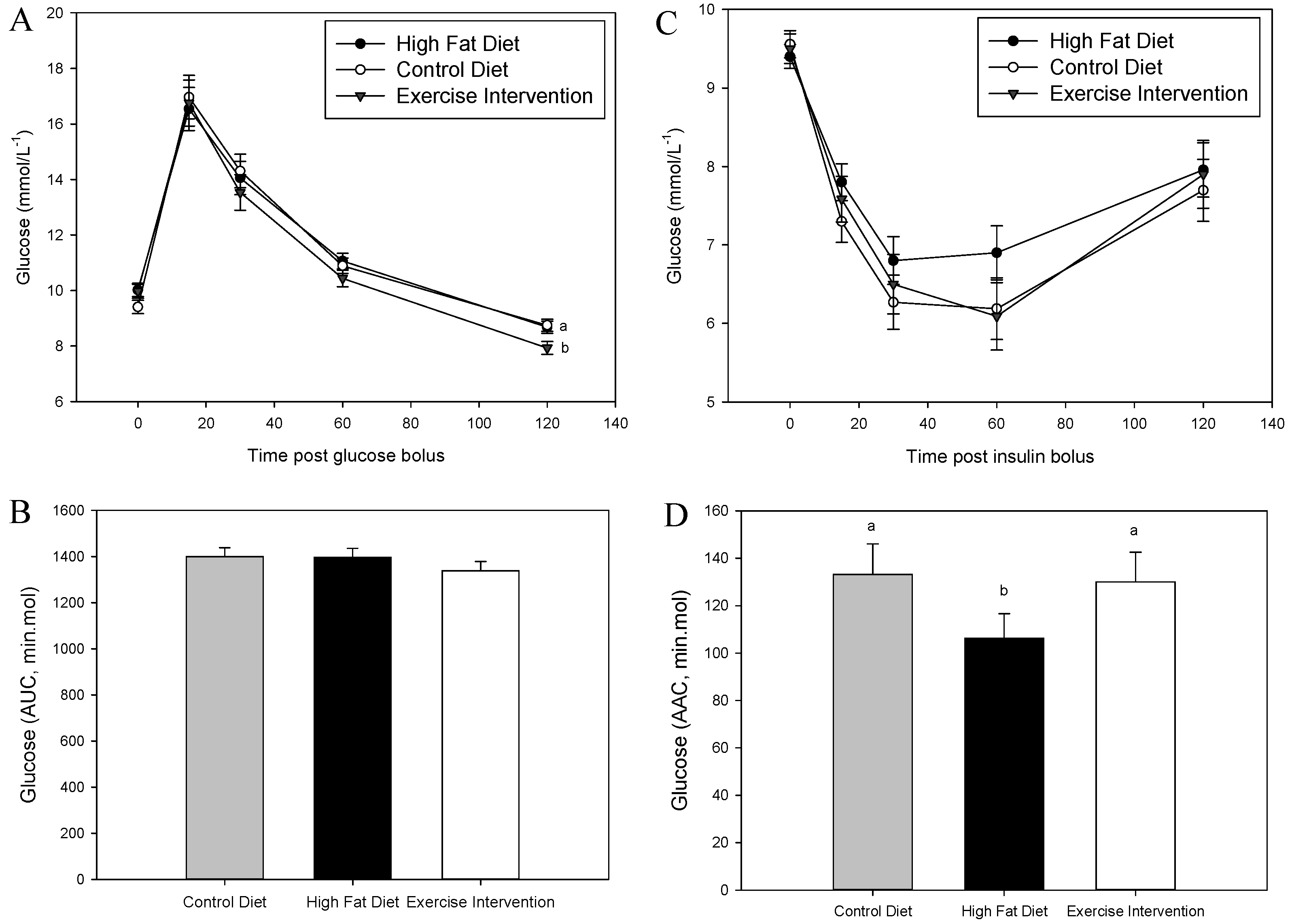
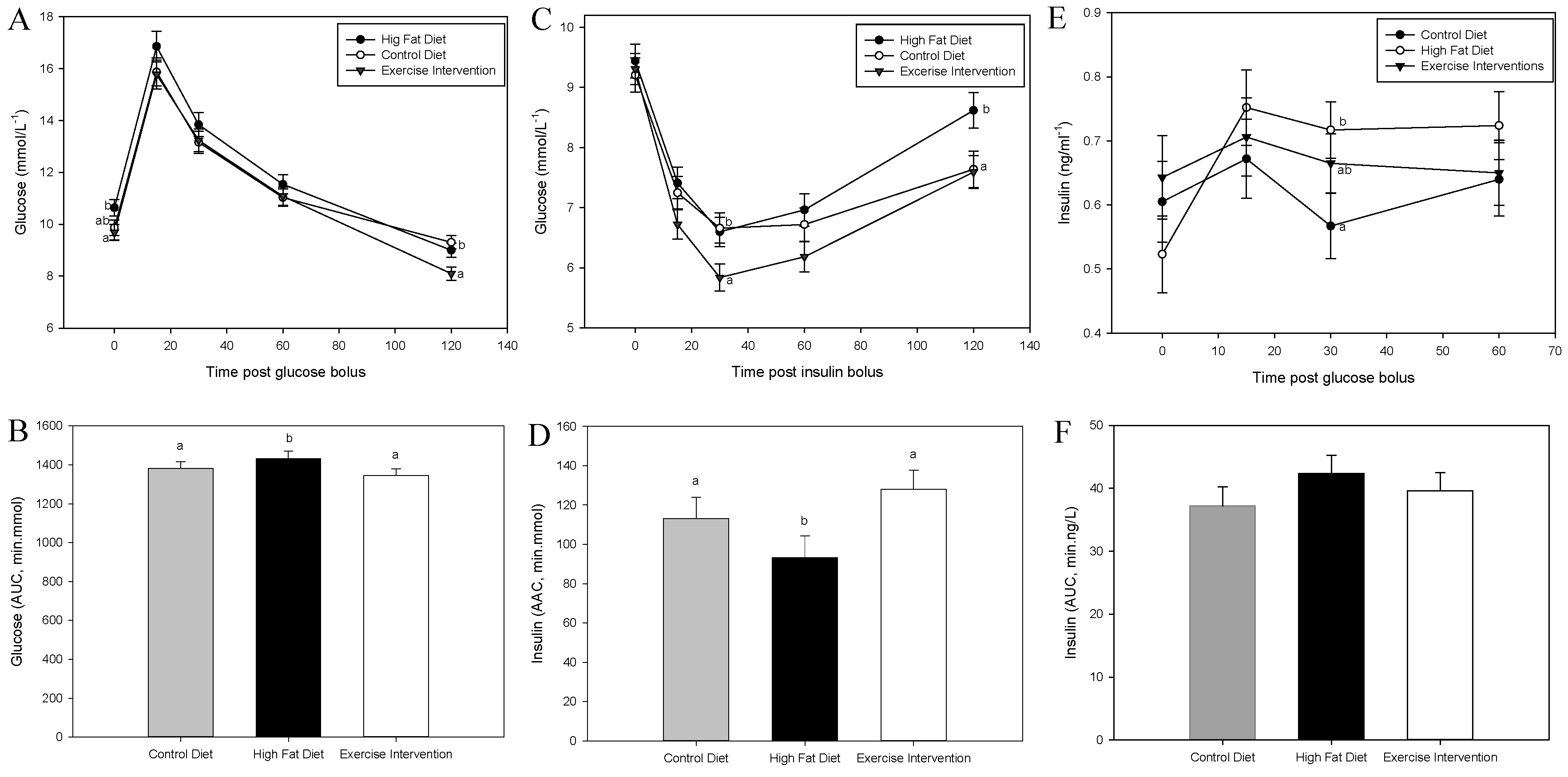
3.3. Exercise Intervention in Fathers Restores Glucose and Insulin Sensitivity in Male Offspring
3.4. Exercise Intervention in Fathers Reduced Total Adiposity and Plasma Lipids in Male Offspring
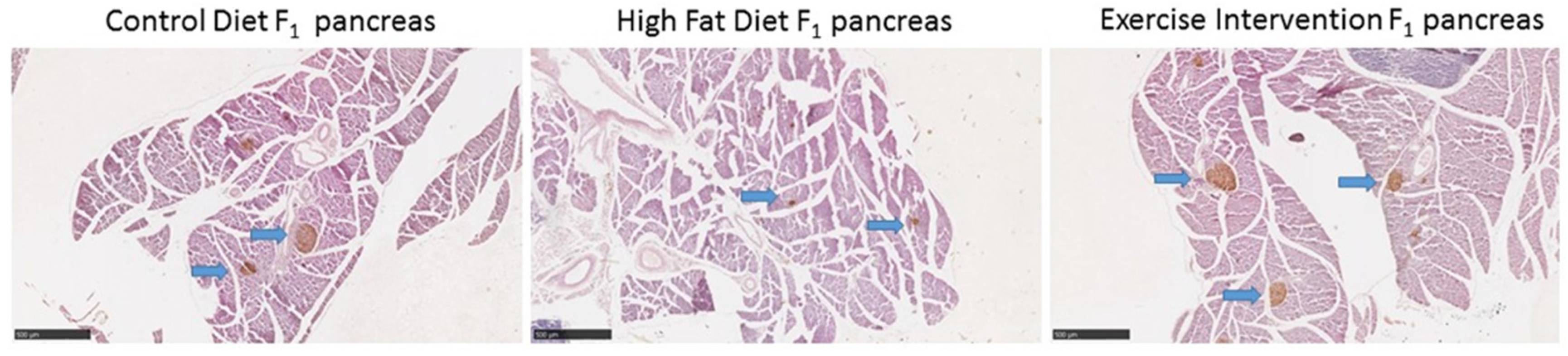
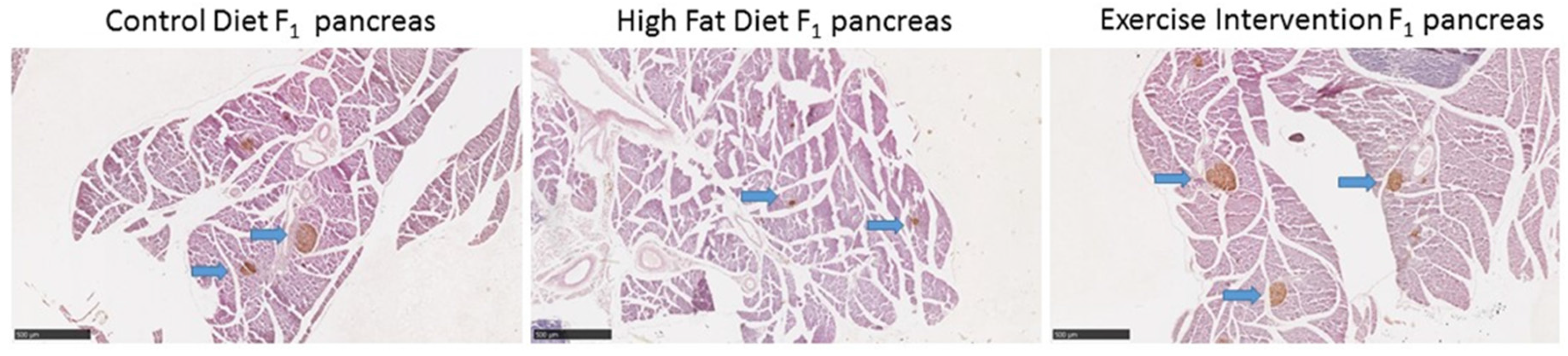
3.5. Paternal Obesity Reduced Islet Cell Number and Size in Male Offspring, Which Was Partially Restored by Exercise Intervention in Fathers
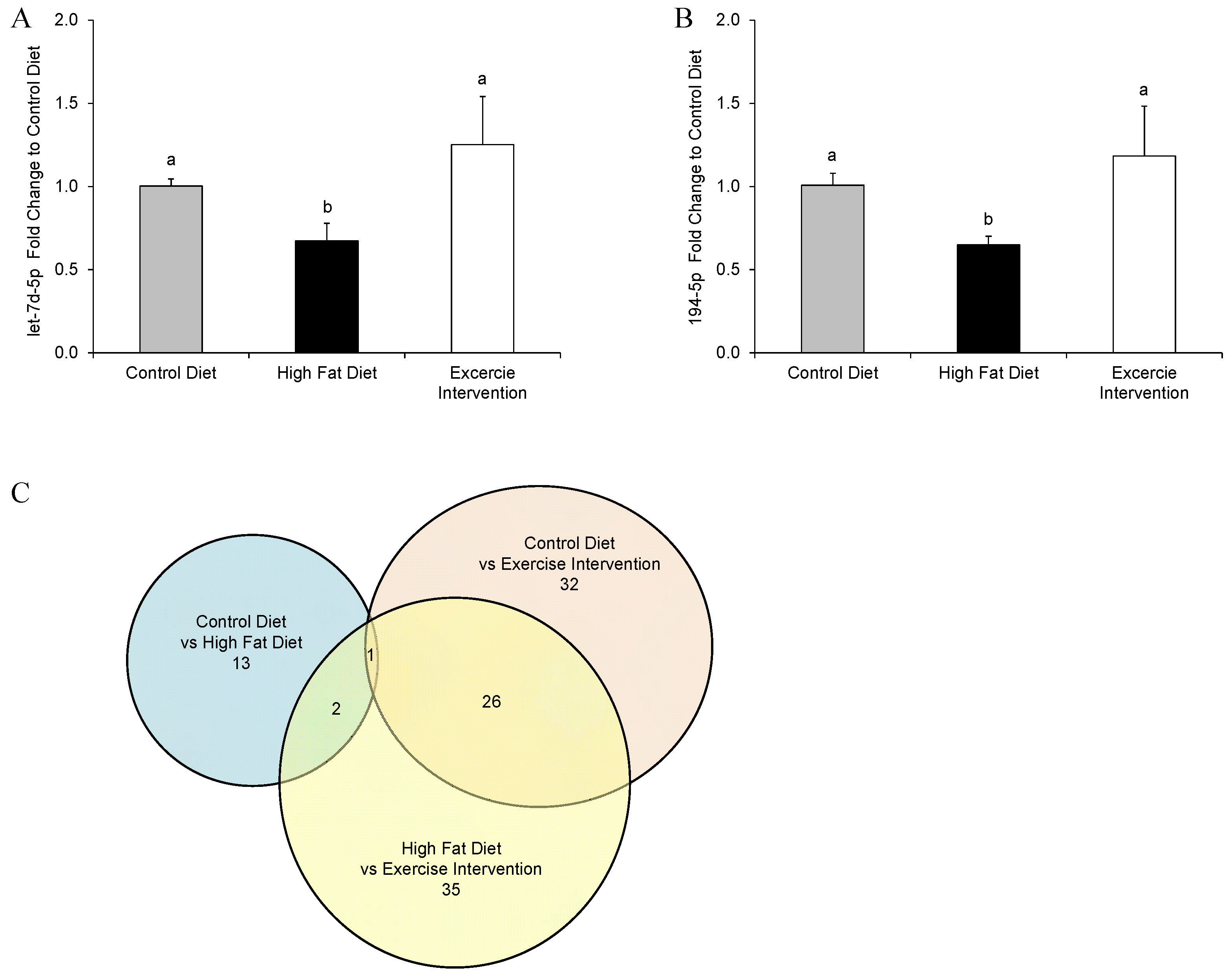
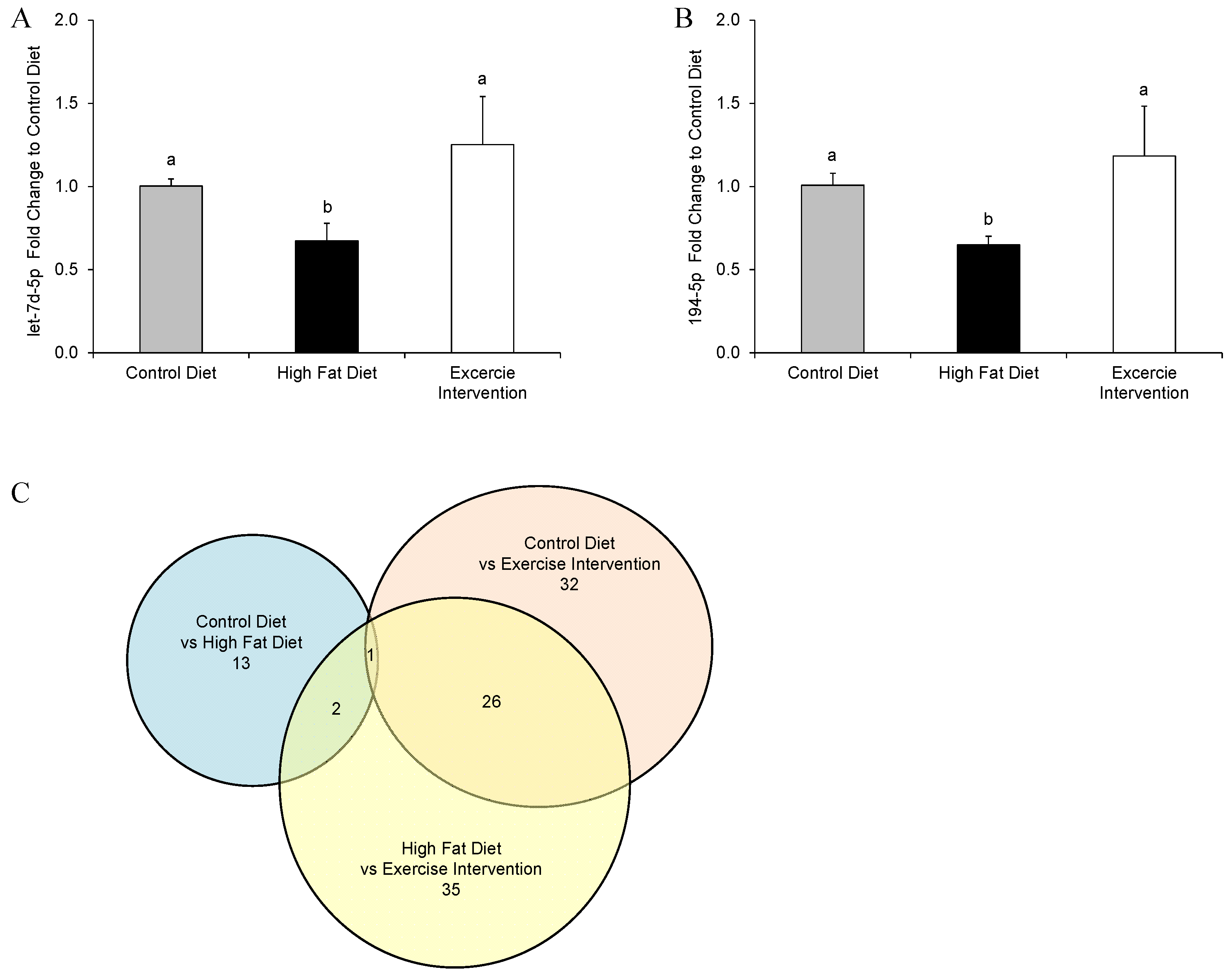
3.6. Paternal Obesity Alters Pancreatic MicroRNA Expression inMale Offspring, Which Was Partially Restored by Exercise Intervention in Fathers
4. Discussion
4.1. Prolonged Paternal HFD Feeding Induces Glucose Intolerance, Insulin Resistance, Pancreatic Islet Cell Dysfunction, and Hyperlipidaemia in Male Offspring
4.2. A Short-Term Founder Exercise Intervention Normalised the Metabolic Profile of Male Offspring
4.3. Exercise Intervention in Founders Further Changed Adipose Accumulation, Circulating FFAs, and Pancreatic MicroRNA Abundance in Male Offspring
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Australian Bureau of Statistics (ABS). Gender Indicators; Australian Bureau of Statistics: Canberra, Australia, 2013. [Google Scholar]
- Flegal, K.M.; Carroll, M.D.; Kit, B.K.; Ogden, C.L. Prevalence of obesity and trends in the distribution of body mass index among us adults, 1999–2010. JAMA 2012, 307, 491–497. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, J.R.; Reynolds, R.M. The risk of maternal obesity to the long-term health of the offspring. Clin. Endocrinol. (Oxf.) 2013, 78, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Isganaitis, E.; Woo, M.; Ma, H.; Chen, M.; Kong, W.; Lytras, A.; Sales, V.; Decoste-Lopez, J.; Lee, K.J.; Leatherwood, C.; et al. Developmental programming by maternal insulin resistance: Hyperinsulinemia, glucose intolerance, and dysregulated lipid metabolism in male offspring of insulin-resistant mice. Diabetes 2014, 63, 688–700. [Google Scholar] [CrossRef] [PubMed]
- Zalbahar, N.; Najman, J.; McIntrye, H.D.; Mamun, A. Parental pre-pregnancy BMI influences on offspring BMI and waist circumference at 21 years. Aust. N. Z. J. Public Health 2016, 40, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Fullston, T.; Palmer, N.O.; Owens, J.A.; Mitchell, M.; Bakos, H.W.; Lane, M. Diet-induced paternal obesity in the absence of diabetes diminishes the reproductive health of two subsequent generations of mice. Hum. Reprod. 2012, 27, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Fullston, T.; Teague, E.M.; Palmer, N.O.; de Blasio, M.J.; Mitchell, M.; Print, C.G.; Owens, J.A.; Lane, M. Paternal obesity initiates metabolic disturbances in two generations of mice and alters the transcription profile of tesis and sperm microrna content. FASEB J. 2013, 27, 4226–4243. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.F.; Lin, R.C.; Laybutt, D.R.; Barres, R.; Owens, J.A.; Morris, M.J. Chronic high-fat diet in fathers programs beta-cell dysfunction in female rat offspring. Nature 2010, 467, 963–966. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.F.; Lin, R.C.; Maloney, C.A.; Youngson, N.A.; Owens, J.A.; Morris, M.J. Paternal high-fat diet consumption induces common changes in the transcriptomes of retroperitoneal adipose and pancreatic islet tissues in female rat offspring. FASEB J. 2014, 28, 1830–1841. [Google Scholar] [CrossRef] [PubMed]
- Cropley, J.E.; Eaton, S.A.; Aiken, A.; Young, P.E.; Giannoulatou, E.; Ho, J.W.; Buckland, M.E.; Keam, S.P.; Hutvagner, G.; Humphreys, D.T.; et al. Male-lineage transmission of an acquired metabolic phenotype induced by grand-paternal obesity. Mol. Metab. 2016, 5, 699–708. [Google Scholar] [CrossRef] [PubMed]
- De Castro Barbosa, T.; Ingerslev, L.R.; Alm, P.S.; Versteyhe, S.; Massart, J.; Rasmussen, M.; Donkin, I.; Sjogren, R.; Mudry, J.M.; Vetterli, L.; et al. High-fat diet reprograms the epigenome of rat spermatozoa and transgenerationally affects metabolism of the offspring. Mol. Metab. 2016, 5, 184–197. [Google Scholar] [CrossRef] [PubMed]
- McPherson, N.O.; Owens, J.A.; Fullston, T.; Lane, M. Preconception diet or exercise interventions in obese fathers normalizes sperm microRNA profile and metabolic syndrome in female offspring. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E805–E821. [Google Scholar] [CrossRef] [PubMed]
- Fullston, T.; Ohlsson-Teague, E.M.; Print, C.G.; Sandeman, L.Y.; Lane, M. Sperm microRNA content is altered in a mouse model of male obesity, but the same suite of microRNAs are not altered in offspring’s sperm. PLoS ONE 2016, 11, e0166076. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, V.; Fourre, S.; De Abreu, D.A.; Derieppe, M.A.; Remy, J.J.; Rassoulzadegan, M. RNA-mediated paternal heredity of diet-induced obesity and metabolic disorders. Sci. Rep. 2015, 5, 18193. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yan, M.; Cao, Z.; Li, X.; Zhang, Y.; Shi, J.; Feng, G.H.; Peng, H.; Zhang, X.; Zhang, Y.; et al. Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder. Science 2016, 351, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Palmer, N.O.; Fullston, T.; Mitchell, M.; Setchell, B.P.; Lane, M. SIRT6 in mouse spermatogenesis is modulated by diet-induced obesity. Reprod. Fertil. Dev. 2011, 23, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Palmer, N.O.; Bakos, H.W.; Owens, J.A.; Setchell, B.P.; Lane, M. Diet and exercise in an obese mouse fed a high-fat diet improve metabolic health and reverse perturbed sperm function. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E768–E780. [Google Scholar] [CrossRef] [PubMed]
- McPherson, N.O.; Fullston, T.; Bakos, H.W.; Setchell, B.P.; Lane, M. Obese father’s metabolic state, adiposity, and reproductive capacity indicate son’s reproductive health. Fertil. Steril. 2014, 101, 865–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPherson, N.O.; Bakos, H.W.; Setchell, B.P.; Owens, J.A.; Lane, M. Improving metabolic health in obese male mice via diet and exercise restores embryo development and fetal growth. PLoS ONE 2013, 8, e71459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brake, D.K.; Smith, E.O.; Mersmann, H.; Smith, C.W.; Robker, R.L. ICAM-1 expression in adipose tissue: Effects of diet-induced obesity in mice. Am. J. Physiol. Cell Physiol. 2006, 291, C1232–C1239. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.; Bakos, H.W.; Lane, M. Paternal diet-induced obesity impairs embryo development and implantation in the mouse. Fertil. Steril. 2011, 95, 1349–1353. [Google Scholar] [CrossRef] [PubMed]
- Bakos, H.W.; Mitchell, M.; Setchell, B.P.; Lane, M. The effect of paternal diet-induced obesity on sperm function and fertilization in a mouse model. Int. J. Androl. 2011, 34, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Lazic, S.E.; Essioux, L. Improving basic and translational science by accounting for litter-to-litter variation in animal models. BMC Neurosci. 2013, 14, 37. [Google Scholar] [CrossRef] [PubMed]
- Nagy, T.R.; Clair, A.L. Precision and accuracy of dual-energy X-ray absorptiometry for determining in vivo body composition of mice. Obes. Res. 2000, 8, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Gatford, K.L.; Mohammad, S.N.; Harland, M.L.; de Blasio, M.J.; Fowden, A.L.; Robinson, J.S.; Owens, J.A. Impaired beta-cell function and inadequate compensatory increases in beta-cell mass after intrauterine growth restriction in sheep. Endocrinology 2008, 149, 5118–5127. [Google Scholar] [CrossRef] [PubMed]
- McPherson, N.O.; Fullston, T.; Owens, J.A.; Lane, M. Paternal lifestyle interventions in obese males restores early embryo development and fetal weights, improving the metabolic health and adiposity status in subsequent female offspring. PLoS ONE 2013, 8, e71459. [Google Scholar]
- Wilcox, G. Insulin and insulin resistance. Clin. Biochem. Rev. 2005, 26, 19–39. [Google Scholar] [PubMed]
- Mirra, P.; Raciti, G.A.; Nigro, C.; Fiory, F.; D’Esposito, V.; Formisano, P.; Beguinot, F.; Miele, C. Circulating mirnas as intercellular messengers, potential biomarkers and therapeutic targets for type 2 diabetes. Epigenomics 2015, 7, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Deiuliis, J.A. Micrornas as regulators of metabolic disease: Pathophysiologic significance and emerging role as biomarkers and therapeutics. Int. J. Obes. (Lond.) 2016, 40, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Costes, S.; Langen, R.; Gurlo, T.; Matveyenko, A.V.; Butler, P.C. Beta-cell failure in type 2 diabetes: A case of asking too much of too few? Diabetes 2013, 62, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.E.; Janson, J.; Bonner-Weir, S.; Ritzel, R.; Rizza, R.A.; Butler, P.C. Beta-cell deficit and increased beta-cell apoptosis in humans with type 2 diabetes. Diabetes 2003, 52, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Binder, N.K.; Beard, S.A.; Kaitu’u-Lino, T.J.; Tong, S.; Hannan, N.J.; Gardner, D.K. Paternal obesity in a rodent model affects placental gene expression in a sex-specific manner. Reproduction 2015, 149, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, A.B.; Morgan, C.P.; Bronson, S.L.; Revello, S.; Bale, T.L. Paternal stress exposure alters sperm microrna content and reprograms offspring hpa stress axis regulation. J. Neurosci. 2013, 33, 9003–9012. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A. Acute effects of exercise on glucose tolerance in non-insulin-dependent diabetes. Med. Sci. Sports Exerc. 1989, 21, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.M.; Riffle, L.; Wilson, R.; Travlos, G.S.; Lubomirski, M.S.; Alvord, W.G. Preconceptional fasting of fathers alters serum glucose in offspring of mice. Nutrition 2006, 22, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.J.; Olson, E.N. Control of glucose homeostasis and insulin sensitivity by the let-7 family of micrornas. Proc. Natl. Acad. Sci. USA 2011, 108, 21075–21080. [Google Scholar] [CrossRef] [PubMed]
- Barakat, R.; Pelaez, M.; Cordero, Y.; Perales, M.; Lopez, C.; Coteron, J.; Mottola, M.F. Exercise during pregnancy protects against hypertension and macrosomia: Randomized clinical trial. Am. J. Obstet. Gynecol. 2016, 214, 649. [Google Scholar] [CrossRef] [PubMed]
- Wajchenberg, B.L. Subcutaneous and visceral adipose tissue: Their relation to the metabolic syndrome. Endocr. Rev. 2000, 21, 697–738. [Google Scholar] [CrossRef] [PubMed]
- Warensjo, E.; Riserus, U.; Vessby, B. Fatty acid composition of serum lipids predicts the development of the metabolic syndrome in men. Diabetologia 2005, 48, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Banzet, S.; Chennaoui, M.; Girard, O.; Racinais, S.; Drogou, C.; Chalabi, H.; Koulmann, N. Changes in circulating micrornas levels with exercise modality. J. Appl. Physiol. 2013, 115, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Sawada, S.; Kon, M.; Wada, S.; Ushida, T.; Suzuki, K.; Akimoto, T. Profiling of circulating micrornas after a bout of acute resistance exercise in humans. PLoS ONE 2013, 8, e70823. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, A.B.; Morgan, C.P.; Leu, N.A.; Bale, T.L. Transgenerational epigenetic programming via sperm microrna recapitulates effects of paternal stress. Proc. Natl. Acad. Sci. USA 2015, 112, 13699–13704. [Google Scholar] [CrossRef] [PubMed]
- Gapp, K.; Jawaid, A.; Sarkies, P.; Bohacek, J.; Pelczar, P.; Prados, J.; Farinelli, L.; Miska, E.; Mansuy, I.M. Implication of sperm RNAs in transgenerational inheritance of the effects of early trauma in mice. Nat. Neurosci. 2014, 17, 667–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, U.; Conine, C.C.; Shea, J.M.; Boskovic, A.; Derr, A.G.; Bing, X.Y.; Belleannee, C.; Kucukural, A.; Serra, R.W.; Sun, F.; et al. Biogenesis and function of trna fragments during sperm maturation and fertilization in mammals. Science 2016, 351, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Waldron, D. Non-coding RNA: Inheritance of diet-induced metabolic changes via tsrnas. Nat. Rev. Genet. 2016, 17, 128. [Google Scholar] [CrossRef] [PubMed]
- Gapp, K.; Miska, E.A. TRNA fragments: Novel players in intergenerational inheritance. Cell Res. 2016, 26, 395–396. [Google Scholar] [CrossRef] [PubMed]
- Murashov, A.K.; Pak, E.S.; Koury, M.; Ajmera, A.; Jeyakumar, M.; Parker, M.; Williams, O.; Ding, J.; Walters, D.; Neufer, P.D. Paternal long-term exercise programs offspring for low energy expenditure and increased risk for obesity in mice. FASEB J. 2016, 30, 775–784. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Diet | High-Fat Diet | Exercise Intervention | |
|---|---|---|---|
| Pre intervention | |||
| Total body weight (g) | 26.6 ± 1.3 a | 32.6 ± 1.1 b | 32.1 ± 1.1 b |
| % of weight gained | 28.0 a | 43.0 b | 45.0 b |
| Adipose tissue (% of total body weight) | 15.1 a | 25.3 b | 24.7 b |
| Post intervention | |||
| Total body weight (g) | 29.3 ± 1.1 a | 35.8 ± 0.9 b | 32.3 ± 0.9 c,* |
| % of weight gained | 9.2 a | 11.2 a | 0.2 b |
| Adipose tissue (% of total body weight) | 17.5 a | 28.2 b | 23.7 c,& |
| Metabolites | |||
| Glucose AUC (min·mmol) | 1670 ± 131 a | 2106 ± 110 b | 1562 ± 111 a |
| Insulin AAC (min·mmol) | 148 ± 17 a | 102 ± 14 b | 93 ± 15 b |
| Cholesterol (mmol/L−1) | 3.14 ± 0.49 a | 4.39 ± 0.42 b,# | 3.90 ± 0.41 a,b |
| Control Diet | High-Fat Diet | Exercise Intervention | |
|---|---|---|---|
| Total body weight | 24.9 ± 0.3 | 25.4 ± 0.3 | 24.3 ± 0.3 |
| DEXA Body Composition | |||
| Grams (g) | |||
| Adipose tissue | 2.07 ± 0.06 a | 2.07 ± 0.06 a | 1.76 ± 0.06 b |
| Lean Mass | 22.0 ± 0.3 | 22.1 ± 0.3 | 21.6 ± 0.3 |
| Bone | 0.43 ± 0.01 | 0.42 ± 0.01 | 0.44 ± 0.01 |
| % of total body weight | |||
| Adipose tissue | 8.31 ± 0.20 a | 8.17 ± 0.21 a | 7.29 ± 0.21 b |
| Lean Mass | 88.1 ± 0.6 a,b | 86.9 ± 0.7 a | 89.0 ± 0.7 b |
| Bone | 3.77 ± 0.13 | 3.54 ± 0.14 | 3.84 ± 0.15 |
| Post Mortem Body Composition | |||
| Grams (g) | |||
| Liver | 0.87 ± 0.02 a | 1.04 ± 0.02 b | 0.92 ± 0.02 a |
| Pancreas | 0.15 ± 0.01 | 0.17 ± 0.01 | 0.15 ± 0.01 |
| Kidneys | 0.27 ± 0.01 a | 0.29 ± 0.01 b | 0.25 ± 0.08 a |
| Soleus (mg) | 8.44 ± 0.48 a | 9.70 ± 0.47 a | 6.43 ± 0.47 b |
| Vastus Lateralis (mg) | 144.9 ± 5.7 | 159.7 ± 5.7 a,b | 145.0 ± 4.5 |
| % of total body weight | |||
| Gonadal Adiposity | 1.08 ± 0.09 | 1.04 ± 0.09 | 0.98 ± 0.09 |
| Omental Adiposity | 0.20 ± 0.03 | 0.23 ± 0.03 | 0.16 ± 0.03 |
| Perirenal Adiposity | 0.09 ± 0.02 | 0.09 ± 0.01 | 0.08 ± 0.01 |
| Retro Adiposity | 0.14 ± 0.02 | 0.12 ± 0.02 | 0.10 ± 0.01 |
| Dorsal Adiposity | 0.39 ± 0.02 | 0.39 ± 0.02 | 0.39 ± 0.02 |
| Total sum Adiposity | 1.89 ± 0.14 | 1.86 ± 0.14 | 1.71 ± 0.12 |
| Liver | 3.67 ± 0.10 a | 3.98 ± 0.10 b | 3.84 ± 0.10 a,b |
| Pancreas | 0.61 ± 0.02 a | 0.67 ± 0.02 b | 0.63 ± 0.02 a |
| Kidneys | 1.13 ± 0.02 a | 1.11 ± 0.02 a,b | 1.06 ± 0.02 b |
| Metabolites | |||
| Glucose (mmol/L−1) | 9.15 ± 0.60 | 7.97 ± 0.69 | 8.54 ± 0.66 |
| Cholesterol (mmol/L−1) | 2.31 ± 0.01 a | 2.41 ± 0.02 b | 2.32 ± 0.01 a |
| FFA (mmol/L−1) | 0.43 ± 0.01 a | 0.42 ± 0.01 a | 0.39 ± 0.01 b |
| Triglycerides (mmol/L−1) | 0.51 ± 0.03 | 0.42 ± 0.03 | 0.40 ± 0.03 |
| Insulin (ng/L−1) | 0.61 ± 0.06 | 0.52 ± 0.06 | 0.64 ± 0.07 |
| Control Diet | High-Fat Diet | Exercise Intervention | |
|---|---|---|---|
| Pancreas (g) | 0.115 ± 0.010 | 0.123 ± 0.008 | 0.116 ± 0.009 |
| Pancreas (% of body weight) | 0.44 ± 0.03 | 0.52 ± 0.03 | 0.51 ± 0.03 |
| Islet cell density (0.1 mm2) | 0.42 ± 0.07 a | 0.29 ± 0.06 b | 0.35 ± 0.07 a,b |
| Small islets (%) (0–5000 µm2) | 62 | 65 | 68 |
| Medium islets (%) (5000–10,000 µm2) | 20 | 15 | 12 |
| Large islets (%) (>10000 µm2) | 18 | 20 | 20 |
| Small islet size (µm2) | 1864 ± 578 | 1900 ± 491 | 2180 ± 506 |
| Medium islet size (µm2) | 6771 ± 1124 | 6551 ± 827 | 7446 ± 1111 |
| Large islet size (µm2) | 20,983 ± 930 a | 15,930 ± 1200 b | 17,778 ± 945 a |
| β-cell area (%) | 0.43 ± 0.11 | 0.32 ± 0.09 | 0.37 ± 0.10 |
| # | Molecules in Network | IPA Score | Focus Molecules | Top Diseases and Functions |
|---|---|---|---|---|
| 1 | ARHGAP5, ATG10, BICD2, CALU, CARHSP1 *, CEP164, COL14A1, CSNK2A1, CTPS1, DNAJA2, ESPL1, FKBP6, FLRT2, HDLBP, IGDCC4, KMT2C, KPNA1, NRARP, PMEPA1, PPFIBP1, PPP2R2A, PTPRU, RAP2B, RBM38, REV3L, RTKN, SEMA6A, SH3BGRL2, SMARCAD1, SPCS2 *, TMEM43, TP53, UBR5, VAPA, ZMAT3 | 38 | 35 | Cancer, Cell Cycle, Hematological Disease |
| 2 | BBX, CHD4, CHD7, DNMT3A, DOT1L, DUSP7, DZIP1, E2F5, E2F6, EZH2, FBXO32, FGF5, GCNT4, GLI3, HISTONE H3, HOXA9, IDH2, IKBKAP, LOR, MEIS2, MYCN, RAB38, RAG1, RBBP4, SALL1, SMARCC1, SOX17, SS18, ST3GAL1, STK40, TLR4 *, TOR1AIP2, UHR *, USP12, ZIC1 | 36 | 34 | Gene Expression, Cell Death and Survival, Embryonic Development |
| 3 | AGO3, AKT2, ANKRD49, ANO1, ATP6V1G1, C5ORF51, CAP1, CPD, GLIS3, HLF, HNB, ID4, MTMR2, MYRF, NR2F2, NUDT4, P2RX1, PHLDA1, PIGA, PLEKHH1, PPARA, PPP1R16B, PRO-INSULIN, RFFL, SECISBP2L, SLC22A23, SNX30, SOCS7, SYPL1 *, TCF7L2, TGFßR1 *, TMEM2 *, TPP1, UCHL5, UGT8 * | 36 | 34 | Digestive System Development and Function, Endocrine System Development and Function, Organ Morphology |
| 4 | ACSL6, ACVR1C, ARID3B, ATP6V1H, CSNK1D *, DICER1 *,EGR3, EIF3J *, ELK4, ERCC6, HAS2, HIC2, HS3ST2, IGDCC3, IGF2BP2 *, KLF8, LPIN2, MAP2, NOTCH, NRIP1, PAG1, PBX3, PLXND1, POLR2C *, PSME3, RARA, RICTOR, RNAPOL II, RORC, SCN11A, SECRETASE Γ, SLC16A10, SLC25A24 *, SYT4, TARBP2 | 31 | 32 | RNA Post-Transcriptional Modification, Cellular Development, Cellular Growth and Proliferation |
| 5 | ACTA1, ACTIN, ALP, ARHGAP20, BMP2K *, CAPG *, CAPN3, CSRNP3, DMD, FZD3, FZD5, GAN, GATM, GPCPD1, GSK3, IGR, KCNC1, LDB3, LHX1, LHX6, MAP4K3, OTP, SEC14L1, SLC25A13, SLC45A4, SLC46A3, SNAP91, TET2, TMEM108, TRIB2, TTC39C, TTC7B, TTL, XK, ZFHX3 | 31 | 32 | Cellular Development, Cellular Growth and Proliferation, Embryonic Development |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McPherson, N.O.; Lane, M.; Sandeman, L.; Owens, J.A.; Fullston, T. An Exercise‐Only Intervention in Obese Fathers Restores Glucose and Insulin Regulation in Conjunction with the Rescue of Pancreatic Islet Cell Morphology and MicroRNA Expression in Male Offspring. Nutrients 2017, 9, 122. https://doi.org/10.3390/nu9020122
McPherson NO, Lane M, Sandeman L, Owens JA, Fullston T. An Exercise‐Only Intervention in Obese Fathers Restores Glucose and Insulin Regulation in Conjunction with the Rescue of Pancreatic Islet Cell Morphology and MicroRNA Expression in Male Offspring. Nutrients. 2017; 9(2):122. https://doi.org/10.3390/nu9020122
Chicago/Turabian StyleMcPherson, Nicole O., Michelle Lane, Lauren Sandeman, Julie A. Owens, and Tod Fullston. 2017. "An Exercise‐Only Intervention in Obese Fathers Restores Glucose and Insulin Regulation in Conjunction with the Rescue of Pancreatic Islet Cell Morphology and MicroRNA Expression in Male Offspring" Nutrients 9, no. 2: 122. https://doi.org/10.3390/nu9020122