The Effects of Tocotrienol and Lovastatin Co-Supplementation on Bone Dynamic Histomorphometry and Bone Morphogenetic Protein-2 Expression in Rats with Estrogen Deficiency



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Annatto Tocotrienol and Lovastatin
2.2. Animal Treatment
2.3. Preparation of Bone Sample
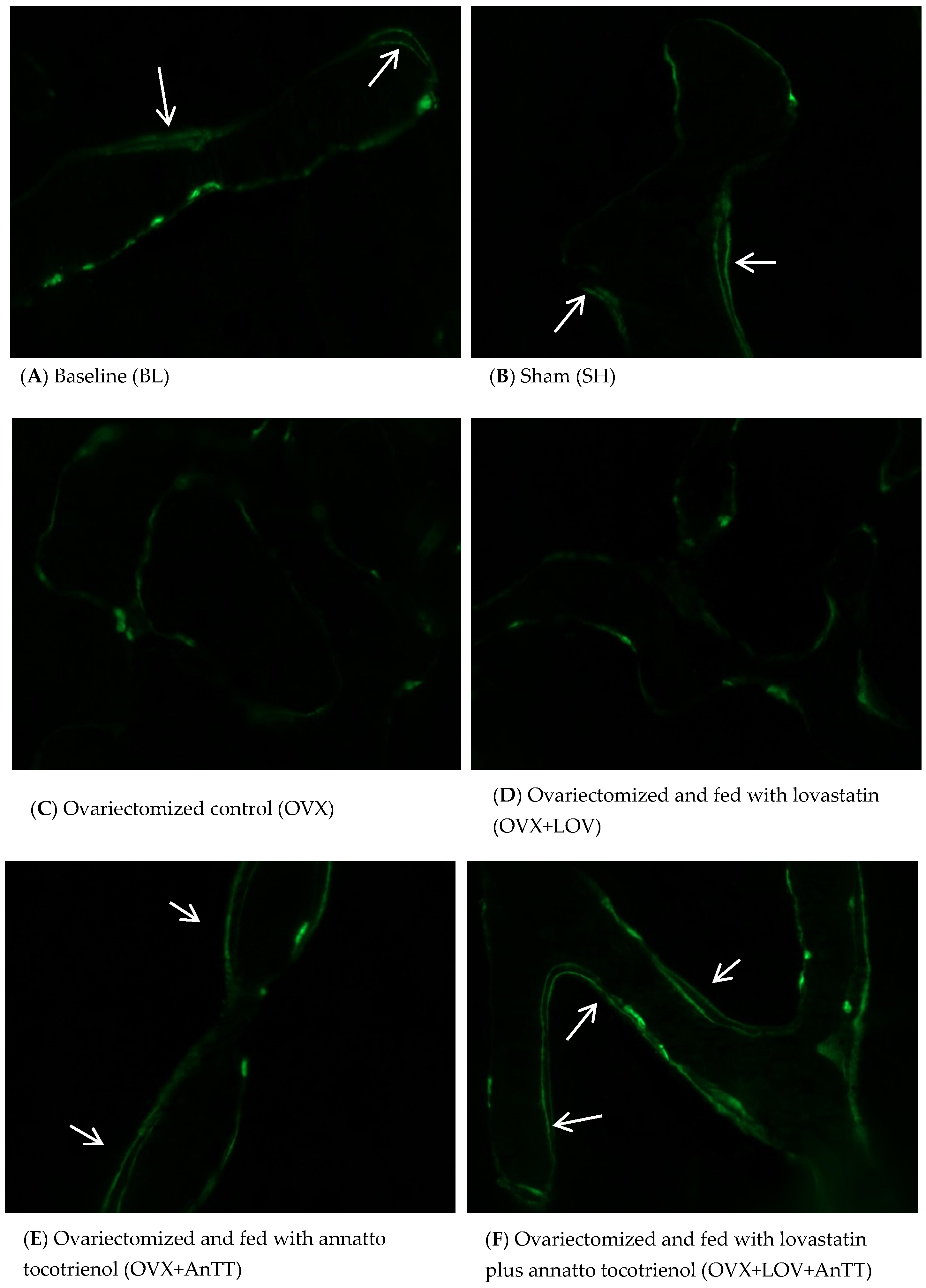
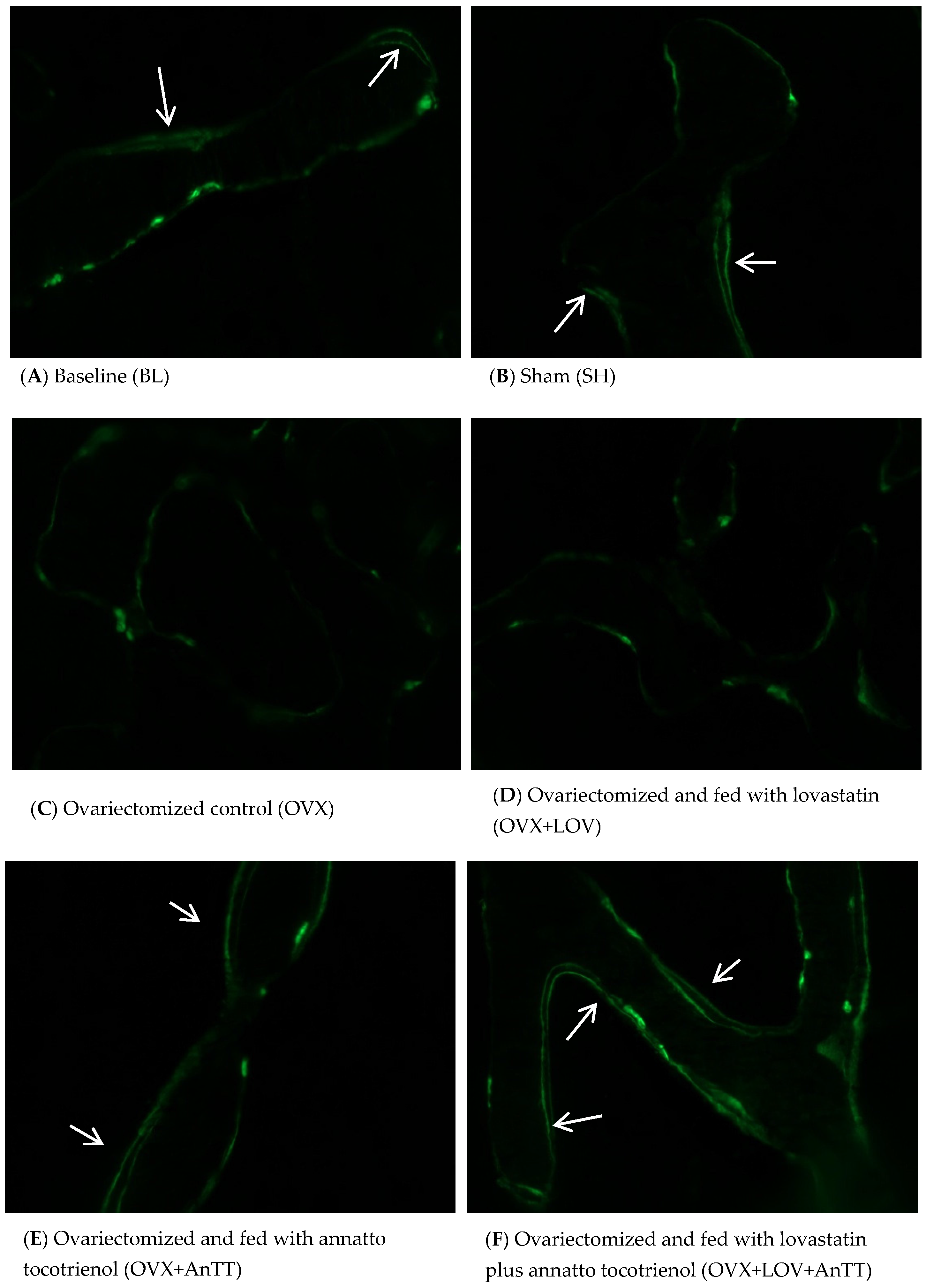
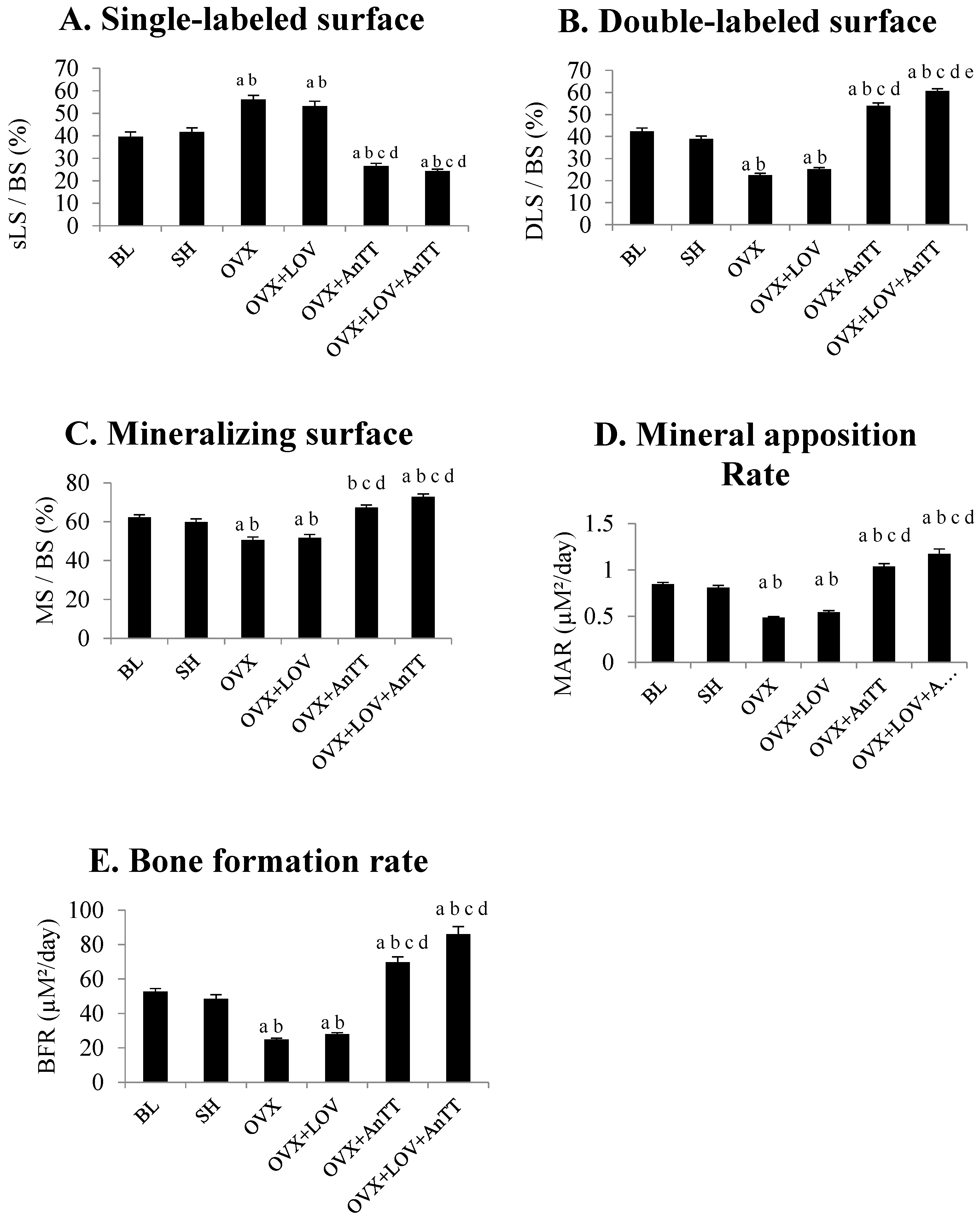
2.4. Assessment of Dynamic Histomorphometric Indices
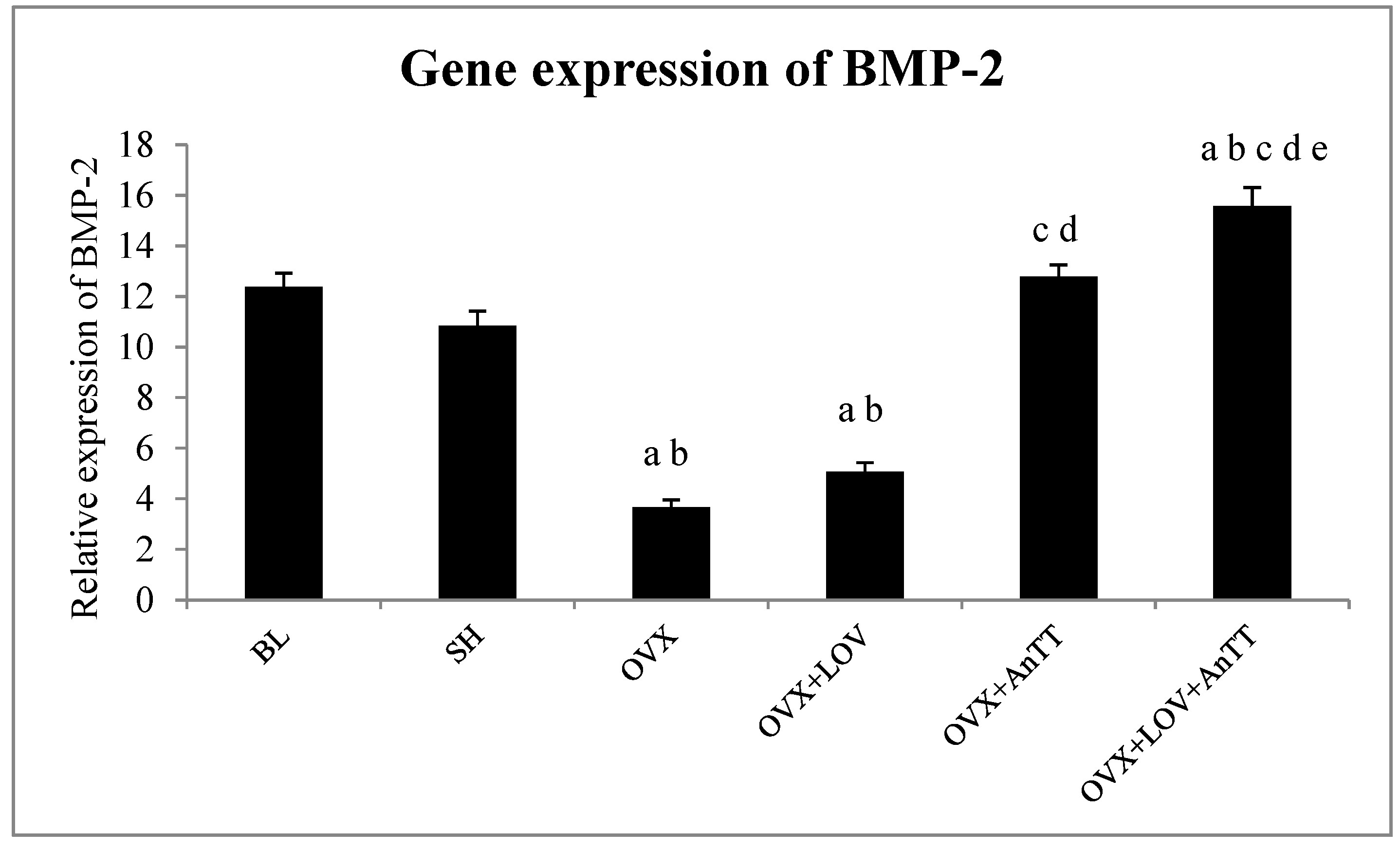
2.5. Determination of Bone Morphogenetics Protein-2 (BMP-2) Expression in Bone
2.6. Statistical Analysis
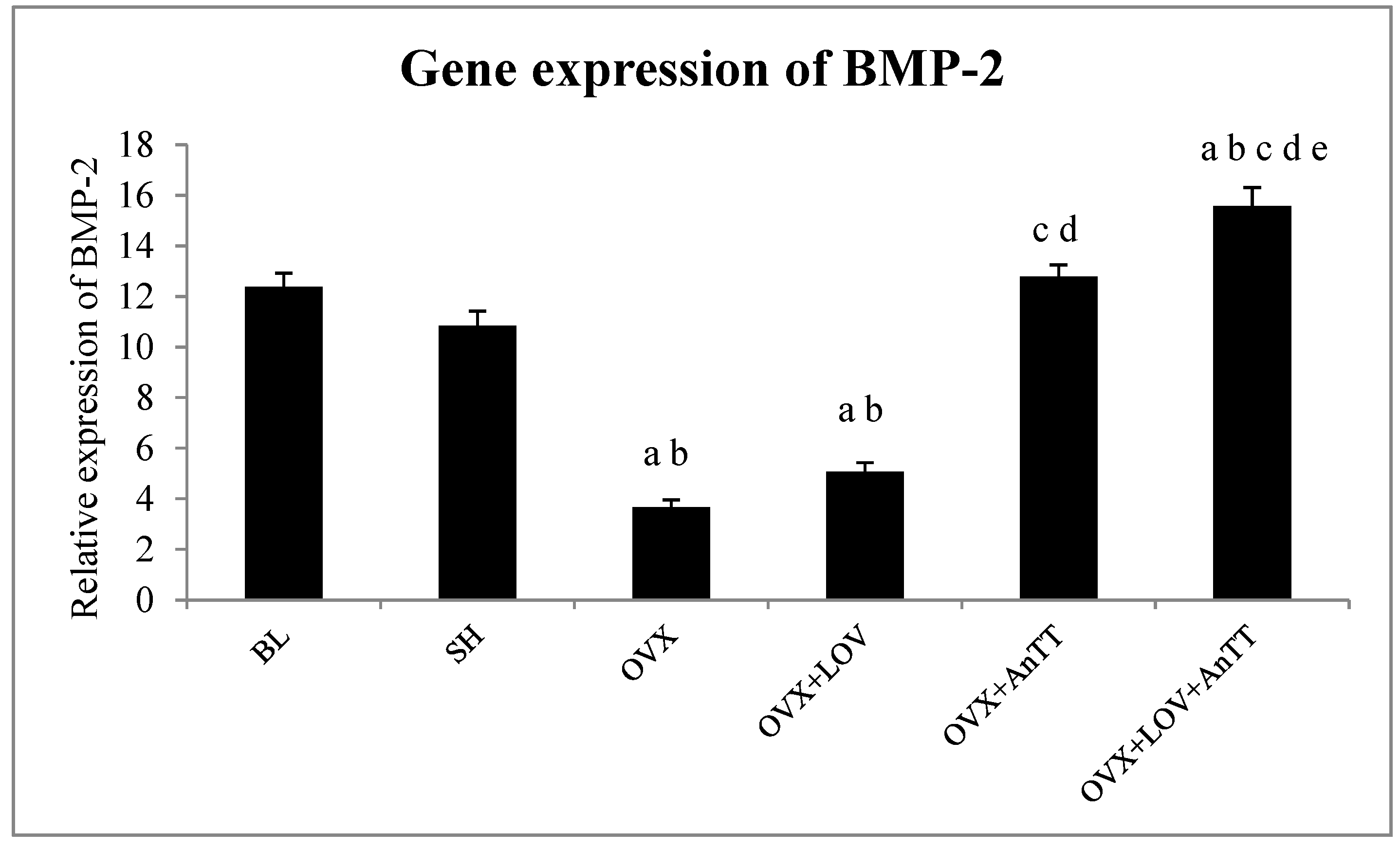
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Basulaiman, M.; El Bcheraoui, C.; Tuffaha, M.; Robinson, M.; Daoud, F.; Jaber, S.; Mikhitarian, S.; Wilson, S.; Memish, Z.A.; Al Saeedi, M.; et al. Hypercholesterolemia and its associated risk factors-Kingdom of Saudi Arabia, 2013. Ann. Epidemiol. 2014, 24, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Guallar-Castillon, P.; Gil-Montero, M.; Leon-Munoz, L.M.; Graciani, A.; Bayan-Bravo, A.; Taboada, J.M.; Banegas, J.R.; Rodriguez-Artalejo, F. Magnitude and management of hypercholesterolemia in the adult population of Spain, 2008–2010: The Enrica Study. Rev. Esp. Cardiol. 2012, 65, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, Y.; Cho, Y.; Jun, B.; Oh, K.W. Trends in the prevalence of major cardiovascular disease risk factors among Korean adults: Results from the Korea National Health and Nutrition Examination Survey, 1998–2012. Int. J. Cardiol. 2014, 174, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Stone, N.J.; Robinson, J.G.; Lichtenstein, A.H.; Bairey Merz, C.N.; Blum, C.B.; Eckel, R.H.; Goldberg, A.C.; Gordon, D.; Levy, D.; Lloyd-Jones, D.M.; et al. 2013 acc/aha guideline on the treatment of blood cholesterol to reduce atherosclerotic cardiovascular risk in adults: A report of the American college of cardiology/american heart association task force on practice guidelines. J. Am. Coll. Cardiol. 2014, 63, 2889–2934. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.H.; Dennison, E.M.; Aihie Sayer, A.; Fielding, R.; Cooper, C. Osteoporosis and sarcopenia in older age. Bone 2015, 80, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Cauley, J.A. Estrogen and bone health in men and women. Steroids 2015, 99, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, L.P.; Yang, X.L.; Huang, H.L.; Ye, D.Q. Hmg-coa reductase inhibitors (statins) and bone mineral density: A meta-analysis. Bone 2013, 54, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Uzzan, B.; Cohen, R.; Nicolas, P.; Cucherat, M.; Perret, G.Y. Effects of statins on bone mineral density: A meta-analysis of clinical studies. Bone 2007, 40, 1581–1587. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, Y.; Zhou, F.; Piao, Z.; Hao, J. Effects of statins on bone mineral density and fracture risk: A prisma-compliant systematic review and meta-analysis. Medicine (Baltimore) 2016, 95, e3042. [Google Scholar] [CrossRef] [PubMed]
- Mo, H.; Yeganehjoo, H.; Shah, A.; Mo, W.K.; Soelaiman, I.N.; Shen, C.L. Mevalonate-suppressive dietary isoprenoids for bone health. J. Nutr. Biochem. 2012, 23, 1543–1551. [Google Scholar] [CrossRef] [PubMed]
- Maritz, F.J.; Conradie, M.M.; Hulley, P.A.; Gopal, R.; Hough, S. Effect of statins on bone mineral density and bone histomorphometry in rodents. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1636–1641. [Google Scholar] [CrossRef] [PubMed]
- Soares, E.A.; Novaes, R.D.; Nakagaki, W.R.; Fernandes, G.J.; Garcia, J.A.; Camilli, J.A. Metabolic and structural bone disturbances induced by hyperlipidic diet in mice treated with simvastatin. Int. J. Exp. Pathol. 2015, 96, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Simic, I.; Reiner, Z. Adverse effects of statins—Myths and reality. Curr. Pharm. Des. 2015, 21, 1220–1226. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.Y.; Ima-Nirwana, S. The biological effects of tocotrienol on bone: A review on evidence from rodent models. Drug Des. Devel. Ther. 2015, 9, 2049–2061. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.-Y.; Ima-Nirwana, S. Vitamin E as an antiosteoporotic agent via receptor activator of nuclear factor kappa-B ligand signaling disruption: Current evidence and other potential research areas. Evid. Based Complement. Altern. Med. 2012, 2012, 747020. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.-Y.; Mo, H.; Soelaiman, I.-N. A review of the possible mechanisms of action of tocotrienol–A potential antiosteoporotic agent. Curr. Drug Targets 2013, 14, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.Y.; Pang, K.L.; Soelaiman, I.N. Tocotrienol and its role in chronic diseases. Adv. Exp. Med. Biol. 2016, 928, 97–130. [Google Scholar] [PubMed]
- Aggarwal, B.; Sundaram, C.; Prasad, S.; Kannappan, R. Tocotrienols, the vitamin E of the 21st century: It’s potential against cancer and other chronic diseases. Biochem. Pharmacol. 2010, 80, 1613–1631. [Google Scholar] [CrossRef] [PubMed]
- Frega, N.; Mozzon, M.; Bocci, F. Identification and estimation of tocotrienols in the annatto lipid fraction by gas chromatography-mass spectrometry. J. Am. Oil Chem. Soc. 1998, 75, 1723–1727. [Google Scholar] [CrossRef]
- Ng, M.; Choo, Y.; Ma, A.; Chuah, C.; Hashim, M. Separation of vitamin E (tocopherol, tocotrienol, and tocomonoenol) in palm oil. Lipids 2004, 39, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.H.; Bergman, C.J. A rapid procedure for analysing rice bran tocopherol, tocotrienol and γ-oryzanol contents. J. Food Compost. Anal. 2005, 18, 139–151. [Google Scholar] [CrossRef]
- Muhammad, N.; Luke, D.A.; Shuid, A.N.; Mohamed, N.; Soelaiman, I.N. Two different isomers of vitamin E prevent bone loss in postmenopausal osteoporosis rat model. Evid. Based Complement. Altern. Med. 2012, 2012, 161527. [Google Scholar] [CrossRef] [PubMed]
- Soelaiman, I.N.; Ming, W.; Abu Bakar, R.; Hashnan, N.A.; Mohd Ali, H.; Mohamed, N.; Muhammad, N.; Shuid, A.N. Palm tocotrienol supplementation enhanced bone formation in oestrogen-deficient rats. Int. J. Endocrinol. 2012, 2012, 532862. [Google Scholar] [CrossRef] [PubMed]
- Nazrun, A.; Khairunnur, A.; Norliza, M.; Norazlina, M.; Ima Nirwana, S. Effects of palm tocotrienol on oxidative stress and bone strength in ovariectomised rats. Med. Health 2008, 3, 83–90. [Google Scholar]
- Muhammad, N.; Razali, S.; Shuid, A.N.; Mohamed, N.; Soelaiman, I.N. Membandingkan kesan antara fraksi-kaya tokotrienol, kalsium dan estrogen terhadap metabolisme tulang tikus terovariektomi. Sains Malays. 2013, 42, 1591–1597. [Google Scholar]
- Abdul-Majeed, S.; Mohamed, N.; Soelaiman, I.-N. Effects of tocotrienol and lovastatin combination on osteoblast and osteoclast activity in estrogen-deficient osteoporosis. Evid. Based Complement. Altern. Med. 2012, 2012, 960742. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Majeed, S.; Mohamed, N.; Soelaiman, I.N. The use of delta-tocotrienol and lovastatin for anti-osteoporotic therapy. Life Sci. 2015, 125, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.Y.; Ima Nirwana, S. The effects of annatto-derived tocotrienol supplementation in osteoporosis induced by testosterone deficiency in rats. Clin. Interv. Aging 2014, 9, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Ding, Y.; Peng, Y.; Wu, Y.; Fan, J.; Li, W.; Yang, R.; Yang, M.; Fu, Q. γ-Tocotrienol protects against ovariectomy-induced bone loss via mevalonate pathway as HMG-CoA reductase inhibitor. Bone 2014, 67, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Vedi, S.; Compston, J. Bone histomorphometry. Methods Mol. Med. 2003, 80, 283–298. [Google Scholar] [PubMed]
- Kanazawa, I.; Yamaguchi, T.; Yano, S.; Hayashi, K.; Yamauchi, M.; Sugimoto, T. Inhibition of the mevalonate pathway rescues the dexamethasone-induced suppression of the mineralization in osteoblasts via enhancing bone morphogenetic protein-2 signal. Horm. Metab. Res. 2009, 41, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Khor, H.T.; Ng, T.T. Effects of administration of alpha-tocopherol and tocotrienols on serum lipids and liver HMG CoA reductase activity. Int. J. Food Sci. Nutr. 2000, 51, S3–S11. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.G.; Thomas, A.M.; Gapor, A.; Tan, B.; Qureshi, N.; Qureshi, A.A. Dose-response impact of various tocotrienols on serum lipid parameters in 5-week-old female chickens. Lipids 2006, 41, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B. The use of simvastatin in bone regeneration. Med. Oral Patol. Oral Cir. Bucal. 2009, 14, e485–e488. [Google Scholar] [PubMed]
- Jadhav, S.B.; Narayana Murthy, P.S.; Singh, M.M.; Jain, G.K. Distribution of lovastatin to bone and its effect on bone turnover in rats. J. Pharm. Pharmacol. 2006, 58, 1451–1458. [Google Scholar] [CrossRef] [PubMed]
- US Preventive Services Task Force. Statin use for the primary prevention of cardiovascular disease in adults: US preventive services task force recommendation statement. JAMA 2016, 316, 1997–2007. [Google Scholar]
- Monteiro, L.O.; Macedo, A.P.; Shimano, R.C.; Shimano, A.C.; Yanagihara, G.R.; Ramos, J.; Paulini, M.R.; Tocchini de Figueiredo, F.A.; Gonzaga, M.G.; Issa, J.P. Effect of treatment with simvastatin on bone microarchitecture of the femoral head in an osteoporosis animal model. Microsc. Res. Tech. 2016, 79, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Starnes, J.W.; Neidre, D.B.; Nyman, J.S.; Roy, A.; Nelson, M.J.; Gutierrez, G.; Wang, X. Synergistic effect of exercise and statins on femoral strength in rats. Exp. Gerontol. 2013, 48, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Farmer, R.; Cooper, R.; Chmielewski, P.A.; Tian, X.Y.; Setterberg, R.B.; Jee, W.S.; Lundy, M.W. Simvastatin did not prevent nor restore ovariectomy-induced bone loss in adult rats. J. Musculoskelet. Neuron. Interact. 2006, 6, 277–283. [Google Scholar]
- Aktifanus, A.T.; Shuid, A.N.; Rashid, N.H.A.; Tam, H.L.; Chua, Y.L.; Saat, N.M.; Muhammad, N.; Mohamed, N.; Ima Nirwana, S. Comparison of the effects of tocotrienol and estrogen on the bone markers and dynamic changes in postmenopausal osteoporosis rat model. Asian J. Anim. Vet. Adv. 2012, 7, 225–234. [Google Scholar]
- Hermizi, H.; Faizah, O.; Ima-Nirwana, S.; Ahmad Nazrun, S.; Norazlina, M. Beneficial effects of tocotrienol and tocopherol on bone histomorphometric parameters in sprague–dawley male rats after nicotine cessation. Calcif. Tissue Int. 2009, 84, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.Y.; Abdul-Majeed, S.; Fozi, N.F.; Ima-Nirwana, S. Annatto tocotrienol improves indices of bone static histomorphometry in osteoporosis due to testosterone deficiency in rats. Nutrients 2014, 6, 4974–4983. [Google Scholar] [CrossRef] [PubMed]
- Ima-Nirwana, S.; Nurshazwani, Y.; Nazrun, A.S.; Norliza, M.; Norazlina, M. Subacute and subchronic toxicity studies of palm vitamin E in mice. J. Pharmacol. Toxicol. 2011, 6, 166–173. [Google Scholar] [CrossRef]
- Abdul-Majeed, S.; Mohamed, N.; Soelaiman, I. A review on the use of statins and tocotrienols, individually or in combination for the treatment of osteoporosis. Curr. Drug Targets 2013, 14, 1579–1590. [Google Scholar] [CrossRef] [PubMed]
- Song, B.L.; DeBose-Boyd, R.A. Insig-dependent ubiquitination and degradation of 3-hydroxy-3-methylglutaryl coenzyme a reductase stimulated by delta- and gamma-tocotrienols. J. Biol. Chem. 2006, 281, 25054–25061. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.A.; Pearce, B.C.; Clark, R.W.; Gordon, D.A.; Wright, J.J. Tocotrienols regulate cholesterol production in mammalian cells by post-transcriptional suppression of 3-hydroxy-3-methylglutaryl-coenzyme a reductase. J. Biol. Chem. 1993, 268, 11230–11238. [Google Scholar] [PubMed]
- Tiwari, V.; Khokhar, M. Mechanism of action of anti-hypercholesterolemia drugs and their resistance. Eur. J. Pharmacol. 2014, 741, 156–170. [Google Scholar] [CrossRef] [PubMed]
- McAnally, J.A.; Gupta, J.; Sodhani, S.; Bravo, L.; Mo, H. Tocotrienols potentiate lovastatin-mediated growth suppression in vitro and in vivo. Exp. Biol. Med. 2007, 232, 523–531. [Google Scholar]
- Wali, V.B.; Sylvester, P.W. Synergistic antiproliferative effects of gamma-tocotrienol and statin treatment on mammary tumor cells. Lipids 2007, 42, 1113–1123. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Deng, C.; Li, Y.P. TGF-beta and bmp signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.Y.; Sun, J.S.; Tsuang, Y.H.; Chen, M.H.; Weng, P.W.; Lin, F.H. Simvastatin promotes osteoblast viability and differentiation via ras/smad/erk/bmp-2 signaling pathway. Nutr. Res. 2010, 30, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Abukhadir, S.S.A.; Mohamed, N.; Makpol, S.; Muhammad, N. Effects of palm vitamin E on bone-formation-related gene expression in nicotine-treated rats. Evid. Based Complement. Altern. Med. 2012, 2012, 656025. [Google Scholar] [CrossRef] [PubMed]
- Salman Khan, M.; Akhtar, S.; Al-Sagair, O.A.; Arif, J.M. Protective effect of dietary tocotrienols against infection and inflammation-induced hyperlipidemia: An in vivo and in silico study. Phytother. Res. 2011, 25, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, A.A.; Reis, J.C.; Qureshi, N.; Papasian, C.J.; Morrison, D.C.; Schaefer, D.M. Delta-tocotrienol and quercetin reduce serum levels of nitric oxide and lipid parameters in female chickens. Lipids Health Dis. 2011, 10, 39. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Primer Sequence | Base Pairs |
|---|---|---|---|
| GAPDH | NM 017008 | F: 5′-GTGGACCTCATGGCCTACAT-3′ | 129 |
| R: 5′-TGTGAGGGAGATGCTCAGTG-3′ | |||
| BMP-2 | NM 017178 | F: 5′-TGAACACAGCTGGTCTCAGG-3′ | 120 |
| R: 5′-TTAAGACGCTTCCGCTGTTT-3′ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chin, K.-Y.; Abdul-Majeed, S.; Mohamed, N.; Ima-Nirwana, S. The Effects of Tocotrienol and Lovastatin Co-Supplementation on Bone Dynamic Histomorphometry and Bone Morphogenetic Protein-2 Expression in Rats with Estrogen Deficiency. Nutrients 2017, 9, 143. https://doi.org/10.3390/nu9020143
Chin K-Y, Abdul-Majeed S, Mohamed N, Ima-Nirwana S. The Effects of Tocotrienol and Lovastatin Co-Supplementation on Bone Dynamic Histomorphometry and Bone Morphogenetic Protein-2 Expression in Rats with Estrogen Deficiency. Nutrients. 2017; 9(2):143. https://doi.org/10.3390/nu9020143
Chicago/Turabian StyleChin, Kok-Yong, Saif Abdul-Majeed, Norazlina Mohamed, and Soelaiman Ima-Nirwana. 2017. "The Effects of Tocotrienol and Lovastatin Co-Supplementation on Bone Dynamic Histomorphometry and Bone Morphogenetic Protein-2 Expression in Rats with Estrogen Deficiency" Nutrients 9, no. 2: 143. https://doi.org/10.3390/nu9020143