Genetic Risk Score of Nine Type 2 Diabetes Risk Variants that Interact with Erythrocyte Phospholipid Alpha-Linolenic Acid for Type 2 Diabetes in Chinese Hans: A Case-Control Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Design
2.2. Genotyping and GRS Creation
2.3. Measurement of Erythrocyte Phospholipid Fatty Acid and Other Covariates
2.4. Statistical Analyses
3. Results
3.1. Population Characteristics
3.2. Association of GRS with T2D, and Erythrocyte Phospholipid n-3 Fatty Acids with T2D
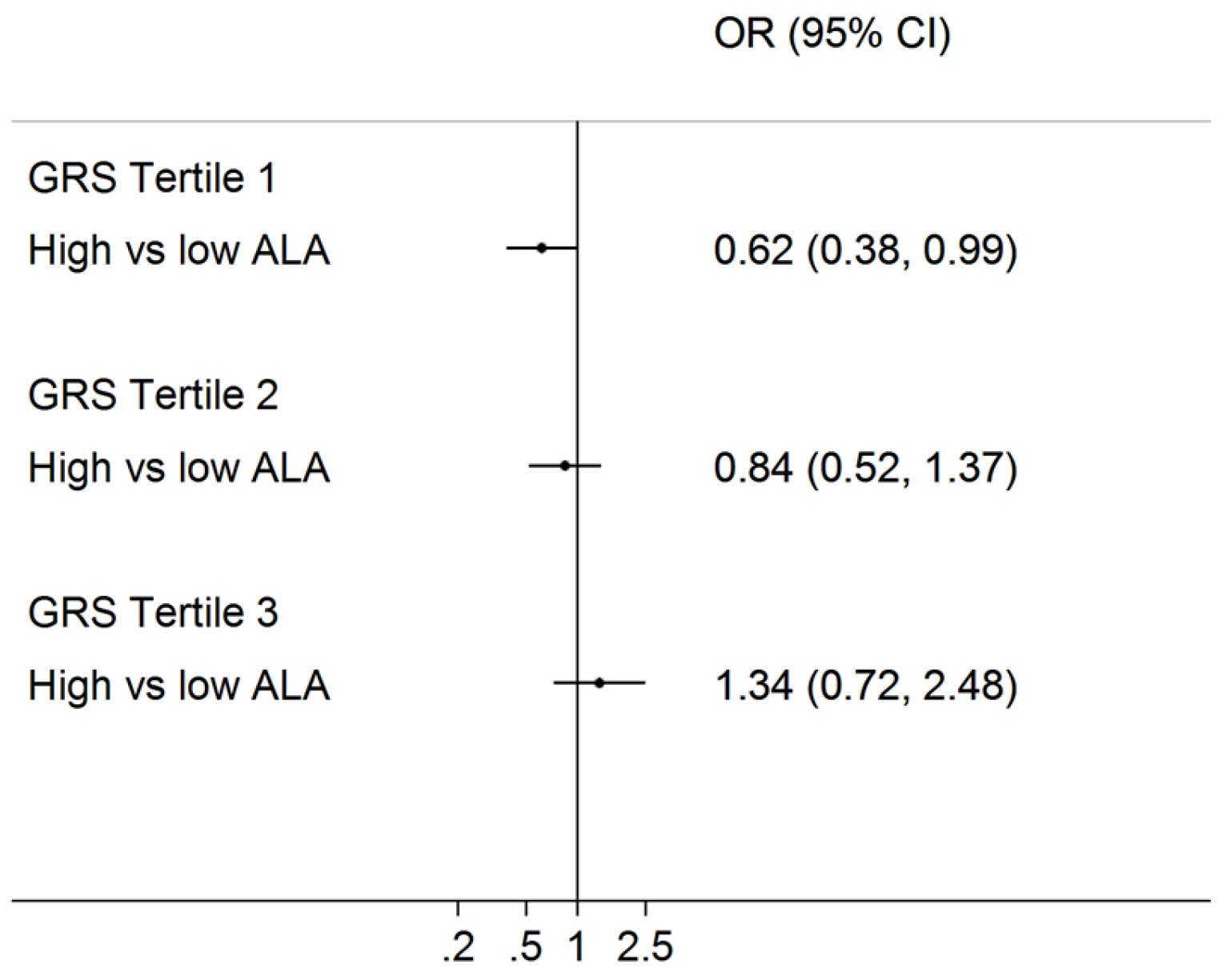
3.3. Interaction of Erythrocyte Phospholipid Fatty Acids with GRS on T2D
4. Discussion
5. Conclutions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Frojdo, S.; Vidal, H.; Pirola, L. Alterations of insulin signaling in type 2 diabetes: A review of the current evidence from humans. Biochim. Biophys. Acta 2009, 1792, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Schellenberg, E.S.; Dryden, D.M.; Vandermeer, B.; Ha, C.; Korownyk, C. Lifestyle interventions for patients with and at risk for type 2 diabetes: A systematic review and meta-analysis. Ann. Intern. Med. 2013, 159, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Snehalatha, C.; Mary, S.; Mukesh, B.; Bhaskar, A.D.; Vijay, V. The indian diabetes prevention programme shows that lifestyle modification and metformin prevent type 2 diabetes in asian indian subjects with impaired glucose tolerance (IDPP-1). Diabetologia 2006, 49, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Lotta, L.A.; Scott, R.A.; Sharp, S.J.; Burgess, S.; Luan, J.; Tillin, T.; Schmidt, A.F.; Imamura, F.; Stewart, I.D.; Perry, J.R.; et al. Genetic predisposition to an impaired metabolism of the branched-chain amino acids and risk of type 2 diabetes: A mendelian randomisation analysis. PLoS Med. 2016, 13, e1002179. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, A.; Go, M.J.; Zhang, W.; Below, J.E.; Gaulton, K.J.; Ferreira, T.; Horikoshi, M.; Johnson, A.D.; Ng, M.C.Y.; Prokopenko, I.; et al. Genome-wide trans-ancestry meta-analysis provides insight into the genetic architecture of type 2 diabetes susceptibility. Nat. Genet. 2014, 46, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, M.C.; Hu, F.B. Gene-environment interactions in the development of type 2 diabetes: Recent progress and continuing challenges. Annu. Rev. Nutr. 2012, 32, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Corella, D.; Ordovas, J.M. Interactions between dietary n-3 fatty acids and genetic variants and risk of disease. Br. J. Nutr. 2012, 107, S271–S283. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.S.; Lai, C.Q.; Parnell, L.D.; Lee, Y.C.; Shen, J.; Smith, C.E.; Casas-Agustench, P.; Richardson, K.; Li, D.; Noel, S.E.; et al. Genome-wide interaction of genotype by erythrocyte n-3 fatty acids contributes to phenotypic variance of diabetes-related traits. BMC Genom. 2014, 15, 781. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.S.; Huang, T.; Li, K.; Chen, Y.; Xie, H.; Xu, D.; Sun, J.; Li, D. Modulation of the association between the pepd variant and the risk of type 2 diabetes by n-3 fatty acids in chinese hans. J. Nutigenet. Nutrigenom. 2015, 8, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L.; Skeaff, C.M.; Fielding, B.A. Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Prog. Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef] [PubMed]
- Lall, K.; Magi, R.; Morris, A.; Metspalu, A.; Fischer, K. Personalized risk prediction for type 2 diabetes: The potential of genetic risk scores. Genet. Med. 2016, 19, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Chu, A.Y.; Kang, J.H.; Huang, J.; Rose, L.M.; Jensen, M.K.; Liang, L.; Curhan, G.C.; Pasquale, L.R.; Wiggs, J.L.; et al. Fried food consumption, genetic risk, and body mass index: Gene-diet interaction analysis in three us cohort studies. BMJ 2014, 348, g1610. [Google Scholar] [CrossRef] [PubMed]
- Langenberg, C.; Sharp, S.J.; Franks, P.W.; Scott, R.A.; Deloukas, P.; Forouhi, N.G.; Froguel, P.; Groop, L.C.; Hansen, T.; Palla, L.; et al. Gene-lifestyle interaction and type 2 diabetes: The epic interact case-cohort study. PLoS Med. 2014, 11, e1001647. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Chu, A.Y.; Kang, J.H.; Jensen, M.K.; Curhan, G.C.; Pasquale, L.R.; Ridker, P.M.; Hunter, D.J.; Willett, W.C.; Rimm, E.B.; et al. Sugar-sweetened beverages and genetic risk of obesity. N. Engl. J. Med. 2012, 367, 1387–1396. [Google Scholar] [CrossRef] [PubMed]
- Welter, D.; MacArthur, J.; Morales, J.; Burdett, T.; Hall, P.; Junkins, H.; Klemm, A.; Flicek, P.; Manolio, T.; Hindorff, L.; et al. The nhgri gwas catalog, a curated resource of snp-trait associations. Nucleic Acids Res. 2014, 42, D1001–D1006. [Google Scholar] [CrossRef] [PubMed]
- James, A.T. Qualitative and quantitative determination of the fatty acids by gas-liquid chromatography. Methods Biochem. Anal. 1960, 8, 1–59. [Google Scholar] [PubMed]
- Fisher, E.; Boeing, H.; Fritsche, A.; Doering, F.; Joost, H.G.; Schulze, M.B. Whole-grain consumption and transcription factor-7-like 2 (TCF7L2) rs7903146: Gene-diet interaction in modulating type 2 diabetes risk. Br. J. Nutr. 2009, 101, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, M.C.; Qi, L.; Kraft, P.; Hu, F.B. Tcf7l2, dietary carbohydrate, and risk of type 2 diabetes in us women. Am. J. Clin. Nutr. 2009, 89, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.S.; Arnett, D.K.; Lee, Y.C.; Shen, J.; Parnell, L.D.; Smith, C.E.; Richardson, K.; Li, D.; Borecki, I.B.; Ordovas, J.M.; et al. Genome-wide contribution of genotype by environment interaction to variation of diabetes-related traits. PLoS ONE 2013, 8, e77442. [Google Scholar] [CrossRef] [PubMed]
- Forouhi, N.G.; Imamura, F.; Sharp, S.J.; Koulman, A.; Schulze, M.B.; Zheng, J.; Ye, Z.; Sluijs, I.; Guevara, M.; Huerta, J.M.; et al. Association of plasma phospholipid n-3 and n-6 polyunsaturated fatty acids with type 2 diabetes: The epic-interact case-cohort study. PLoS Med. 2016, 13, e1002094. [Google Scholar] [CrossRef] [PubMed]
- Bloedon, L.T.; Balikai, S.; Chittams, J.; Cunnane, S.C.; Berlin, J.A.; Rader, D.J.; Szapary, P.O. Flaxseed and cardiovascular risk factors: Results from a double blind, randomized, controlled clinical trial. J. Am. Coll. Nutr. 2008, 27, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Rhee, Y.; Brunt, A. Flaxseed supplementation improved insulin resistance in obese glucose intolerant people: A randomized crossover design. Nutr. J. 2011, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, A.M.; Brown, B.D.; Cunnane, S.C.; Domitrovich, S.G.; Adams, E.R.; Bobowiec, C.E. Daily flaxseed consumption improves glycemic control in obese men and women with pre-diabetes: A randomized study. Nutr. Res. 2013, 33, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Gomes, P.M.; Hollanda-Miranda, W.R.; Beraldo, R.A.; Castro, A.V.; Geloneze, B.; Foss, M.C.; Foss-Freitas, M.C. Supplementation of alpha-linolenic acid improves serum adiponectin levels and insulin sensitivity in patients with type 2 diabetes. Nutrition 2015, 31, 853–857. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G. Alpha-linolenic acid metabolism in men and women: Nutritional and biological implications. Curr. Opin. Clin. Nutr. Metab. Care. 2004, 7, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Bhaswant, M.; Poudyal, H.; Brown, L. Mechanisms of enhanced insulin secretion and sensitivity with n-3 unsaturated fatty acids. J. Nutr. Biochem. 2015, 26, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.S.; Lin, M.; Fang, L.; Yu, Y.; Yuan, L.; Jin, Y.; Feng, J.; Wang, L.; Yang, H.; Chen, W.; et al. Effects of n-3 fatty acid supplements on glycemic traits in chinese type 2 diabetic patients: A double-blind randomized controlled trial. Mol. Nutr. Food Res. 2016, 60, 2176–2184. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A.; Ware, E.B.; Middha, P.; Beacher, L.; Kardia, S.L. Current applications of genetic risk scores to cardiovascular outcomes and subclinical phenotypes. Curr. Epidemiol. Rep. 2015, 2, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, R.M.; Galli, C.; Ferroluzzi, A.; Iacono, J.M. Lipid and phospholipid fatty-acid composition of plasma, red-blood-cells, and platelets and how they are affected by dietary lipids—A study of normal subjects from italy, finland, and the USA. Am. J. Clin. Nutr. 1987, 45, 443–455. [Google Scholar] [PubMed]
- Sun, Q.; Ma, J.; Campos, H.; Hankinson, S.E.; Hu, F.B. Comparison between plasma and erythrocyte fatty acid content as biomarkers of fatty acid intake in us women. Am. J. Clin. Nutr. 2007, 86, 74–81. [Google Scholar] [PubMed]


{kind=link}
| SNP Name | Chromosome | Region | Nearby Gene | p-HWE | MAF | Alleles (Minor Allele) | Risk Allele | Weight | Source |
|---|---|---|---|---|---|---|---|---|---|
| rs831571 | 3 | 3p14.1 | PSMD6 | 0.057 | 0.347 | C/T(T) | C | 0.086 | PMID: 22158537 |
| rs4712524 | 6 | 6p22.3 | CDKAL1 | 0.951 | 0.438 | A/G(G) | G | 0.199 | PMID: 18711366 |
| rs2383208 | 6 | 6p22.3 | CDKN2B | 0.903 | 0.401 | A/G(G) | A | 0.199 | PMID: 18711366 |
| rs13266634 | 8 | 8q24.11 | SLC30A8 | 0.828 | 0.391 | C/T(T) | C | 0.199 | PMID: 19401414 |
| rs11257655 | 10 | 10p13 | CDC123 | 0.249 | 0.419 | C/T(C) | T | 0.14 | PMID: 22961080 |
| rs1359790 | 13 | 13q31.1 | SPRY2 | 0.563 | 0.290 | C/T(T) | C | 0.14 | PMID: 20862305 |
| rs1436953 | 15 | 15q22.2 | C2CD4A, C2CD4B | 0.587 | 0.343 | A/G(A) | G | 0.131 | PMID: 21799836 |
| rs16955379 | 16 | 16q23.2 | CMIP | 0.774 | 0.255 | C/T(T) | C | 0.077 | PMID: 22158537 |
| rs3786897 | 19 | 19q13.11 | PEPD | 0.729 | 0.440 | A/G(G) | A | 0.095 | PMID: 22158537 |
| Healthy Controls (n = 293) | Type 2 Diabetes Cases (n = 622) | |||||
|---|---|---|---|---|---|---|
| GRS T1 (n = 119) | GRS T2 (n = 111) | GRS T3 (n = 63) | GRS T1 (n = 222) | GRS T2 (n = 228) | GRS T3 (n = 172) | |
| Median GRS = 9 | Median GRS = 11 | Median GRS = 13 | Median GRS = 9 | Median GRS = 11.9 | Median GRS = 13.3 | |
| Age, years | 51.8 (13.4) | 50.5 (12.7) | 50.8 (13.8) | 58.7 (12.7) | 59 (11) | 59.2 (11.3) |
| Male, % | 51 (43.6) | 61 (55.5) | 37 (58.7) | 117 (52.7) | 92 (40.4) | 70 (40.9) |
| Height, cm | 169.3 (8.3) | 169.2 (8.1) | 168.6 (8.7) | 164.1 (8.5) | 162.7 (7.6) | 162.5 (8.7) |
| Weight, kg | 69.4 (12.8) | 70.9 (10.1) | 71.3 (10.8) | 68.5 (14.8) | 66.2 (11.8) | 65.3 (12.2) |
| BMI, kg/m2 | 24.1 (3.2) | 24.6 (2.2) | 25.0 (2.7) | 25.3 (4.1) | 24.9 (3.5) | 24.7 (3.47) |
| DBP, mmHg | 73.1 (13.9) | 79.5 (12.6) | 77.7 (10.2) | 80 (10.1) | 80.5 (11) | 79.5 (11.7) |
| SBP, mmHg | 122.9 (14.8) | 128.1 (17.3) | 128.5 (15) | 131.2 (19.5) | 130.4 (18) | 128.9 (18.4) |
| Erythrocyte marine n-3 fatty acids (EPA + DHA + DPA), mol% | 6.06 (4.03–7.76) | 6.40 (3.93–8.22) | 6.01 (4.01–8.68) | 5.62 (3.46–6.4) | 5.37 (3.2–6.5) | 5.02 (3.1–6.03) |
| Erythrocyte EPA, mol% | 1.75 (0.69–2.39) | 1.77 (0.5–2.24) | 1.90 (0.59–2.31) | 1.76 (0.68–1.85) | 1.76 (0.8–1.89) | 1.58 (0.67–1.61) |
| Erythrocyte DHA, mol% | 3.07 (1.52–5.06) | 3.39 (1.54–5.2) | 2.99 (1.72–3.84) | 2.85 (1.65–3.62) | 2.64 (1.31–3.39) | 2.54 (1.55–3.21) |
| Erythrocyte DPA, mol% | 1.25 (0.88–1.59) | 1.24 (0.91–1.49) | 1.13 (0.79–1.46) | 1.01 (0.59–1.31) | 0.97 (0.53–1.27) | 0.90 (0.47–1.17) |
| Erythrocyte ALA, mol% | 0.61 (0.25–0.70) | 0.57 (0.25–0.50) | 0.51 (0.19–0.53) | 0.64 (0.19–0.57) | 0.60 (0.19–0.57) | 0.71 (0.21–0.67) |
| GRS Categories | ALA Categories | Based on Unweighted GRS | Based on Weighted GRS | ||||
|---|---|---|---|---|---|---|---|
| OR | 95% CI | p | OR | 95% CI | p | ||
| GRS Tertile 1 | ALA Quartile 1 | 1 | Reference | 1 | Reference | ||
| ALA Quartile 2 | 0.52 | 0.26–1.06 | 0.073 | 0.65 | 0.32–1.31 | 0.227 | |
| ALA Quartile 3 | 0.42 | 0.21–0.85 | 0.016 | 0.52 | 0.26–1.04 | 0.066 | |
| ALA Quartile 4 | 0.47 | 0.24–0.92 | 0.028 | 0.67 | 0.34–1.33 | 0.258 | |
| Per Quartile | 0.79 | 0.64–0.97 | 0.027 | 0.87 | 0.71–1.08 | 0.212 | |
| GRS Tertile 2 | ALA Quartile 1 | 1 | Reference | 1 | Reference | ||
| ALA Quartile 2 | 0.46 | 0.23–0.92 | 0.028 | 0.36 | 0.17–0.78 | 0.009 | |
| ALA Quartile 3 | 0.61 | 0.3–1.23 | 0.165 | 0.52 | 0.24–1.16 | 0.112 | |
| ALA Quartile 4 | 0.6 | 0.29–1.27 | 0.183 | 0.36 | 0.17–0.8 | 0.012 | |
| Per Quartile | 0.89 | 0.71–1.11 | 0.304 | 0.78 | 0.62–0.99 | 0.043 | |
| GRS Tertile 3 | ALA Quartile 1 | 1 | Reference | 1 | Reference | ||
| ALA Quartile 2 | 0.83 | 0.36–1.92 | 0.665 | 0.69 | 0.32–1.46 | 0.327 | |
| ALA Quartile 3 | 0.9 | 0.38–2.14 | 0.812 | 0.66 | 0.31–1.42 | 0.291 | |
| ALA Quartile 4 | 1.5 | 0.62–3.65 | 0.371 | 1.28 | 0.56–2.92 | 0.558 | |
| Per Quartile | 1.13 | 0.86–1.48 | 0.373 | 1.06 | 0.83–1.35 | 0.658 | |
| ALA Categories | GRS Categories | Based on Unweighted GRS | Based on Weighted GRS | ||||
|---|---|---|---|---|---|---|---|
| OR | 95% CI | p | OR | 95% CI | p | ||
| Low ALA group (≤median) | GRS Tertile 1 | 1 | Ref. | 1 | Ref. | ||
| GRS Tertile 2 | 0.88 | 0.54–1.43 | 0.606 | 1.33 | 0.8–2.21 | 0.266 | |
| GRS Tertile 3 | 0.93 | 0.54–1.6 | 0.794 | 1.35 | 0.81–2.24 | 0.251 | |
| Per risk allele | 0.96 | 0.74–1.25 | 0.763 | 1.16 | 0.9–1.5 | 0.249 | |
| High ALA group (>median) | GRS Tertile 1 | 1 | Ref. | 1 | Ref. | ||
| GRS Tertile 2 | 1.29 | 0.81–2.06 | 0.289 | 1.4 | 0.85–2.3 | 0.185 | |
| GRS Tertile 3 | 2.11 | 1.21–3.66 | 0.008 | 2.03 | 1.21–3.4 | 0.007 | |
| Per risk allele | 1.43 | 1.09–1.87 | 0.009 | 1.42 | 1.1–1.84 | 0.007 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, J.-S.; Li, K.; Huang, T.; Chen, Y.; Xie, H.; Xu, D.; Sun, J.; Li, D. Genetic Risk Score of Nine Type 2 Diabetes Risk Variants that Interact with Erythrocyte Phospholipid Alpha-Linolenic Acid for Type 2 Diabetes in Chinese Hans: A Case-Control Study. Nutrients 2017, 9, 376. https://doi.org/10.3390/nu9040376
Zheng J-S, Li K, Huang T, Chen Y, Xie H, Xu D, Sun J, Li D. Genetic Risk Score of Nine Type 2 Diabetes Risk Variants that Interact with Erythrocyte Phospholipid Alpha-Linolenic Acid for Type 2 Diabetes in Chinese Hans: A Case-Control Study. Nutrients. 2017; 9(4):376. https://doi.org/10.3390/nu9040376
Chicago/Turabian StyleZheng, Ju-Sheng, Kelei Li, Tao Huang, Yanqiu Chen, Hua Xie, Danfeng Xu, Jianqin Sun, and Duo Li. 2017. "Genetic Risk Score of Nine Type 2 Diabetes Risk Variants that Interact with Erythrocyte Phospholipid Alpha-Linolenic Acid for Type 2 Diabetes in Chinese Hans: A Case-Control Study" Nutrients 9, no. 4: 376. https://doi.org/10.3390/nu9040376