Osthole Enhances Osteogenesis in Osteoblasts by Elevating Transcription Factor Osterix via cAMP/CREB Signaling In Vitro and In Vivo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Cell Culture
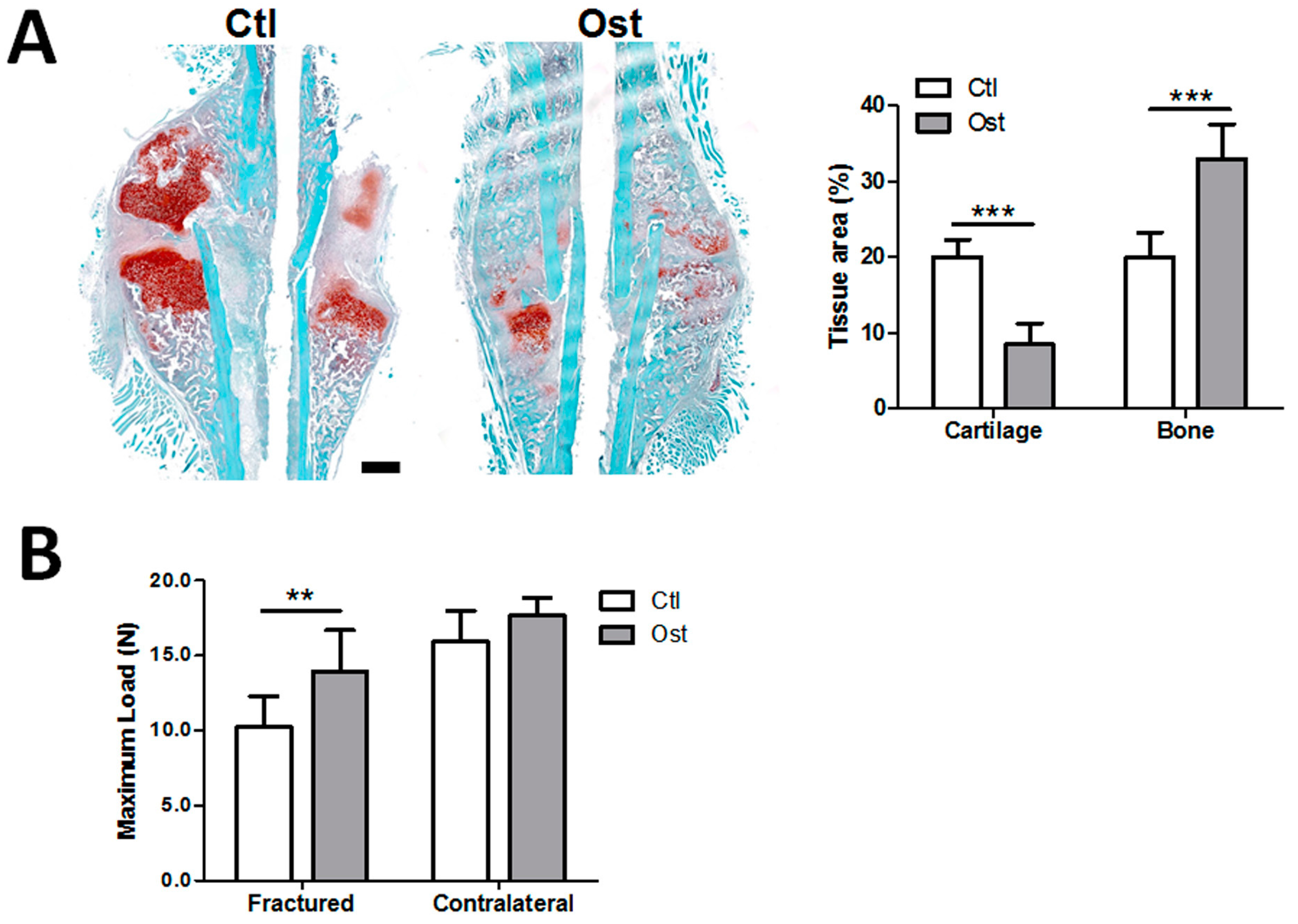
2.2. Mouse Femoral Osteotomy
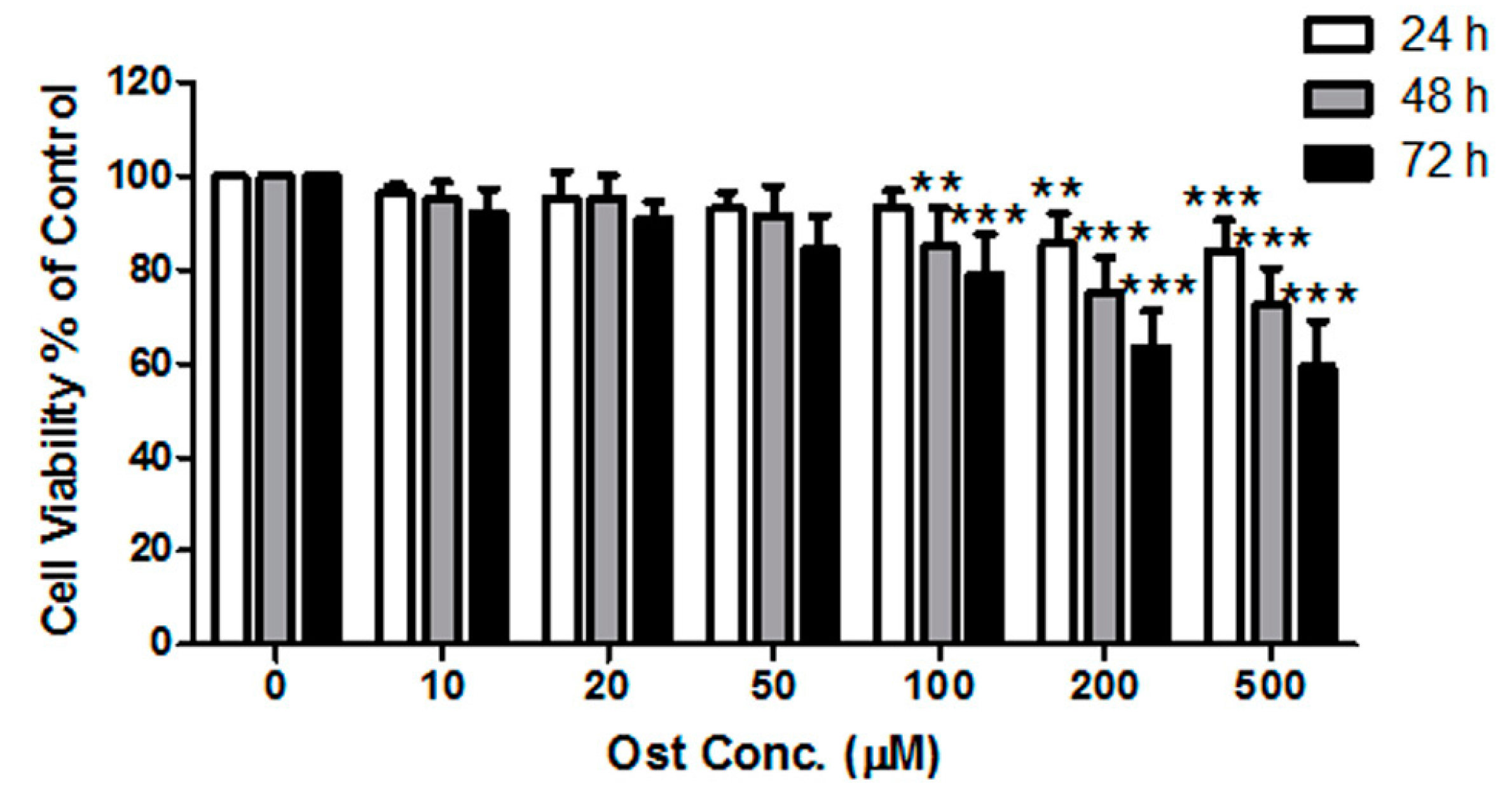
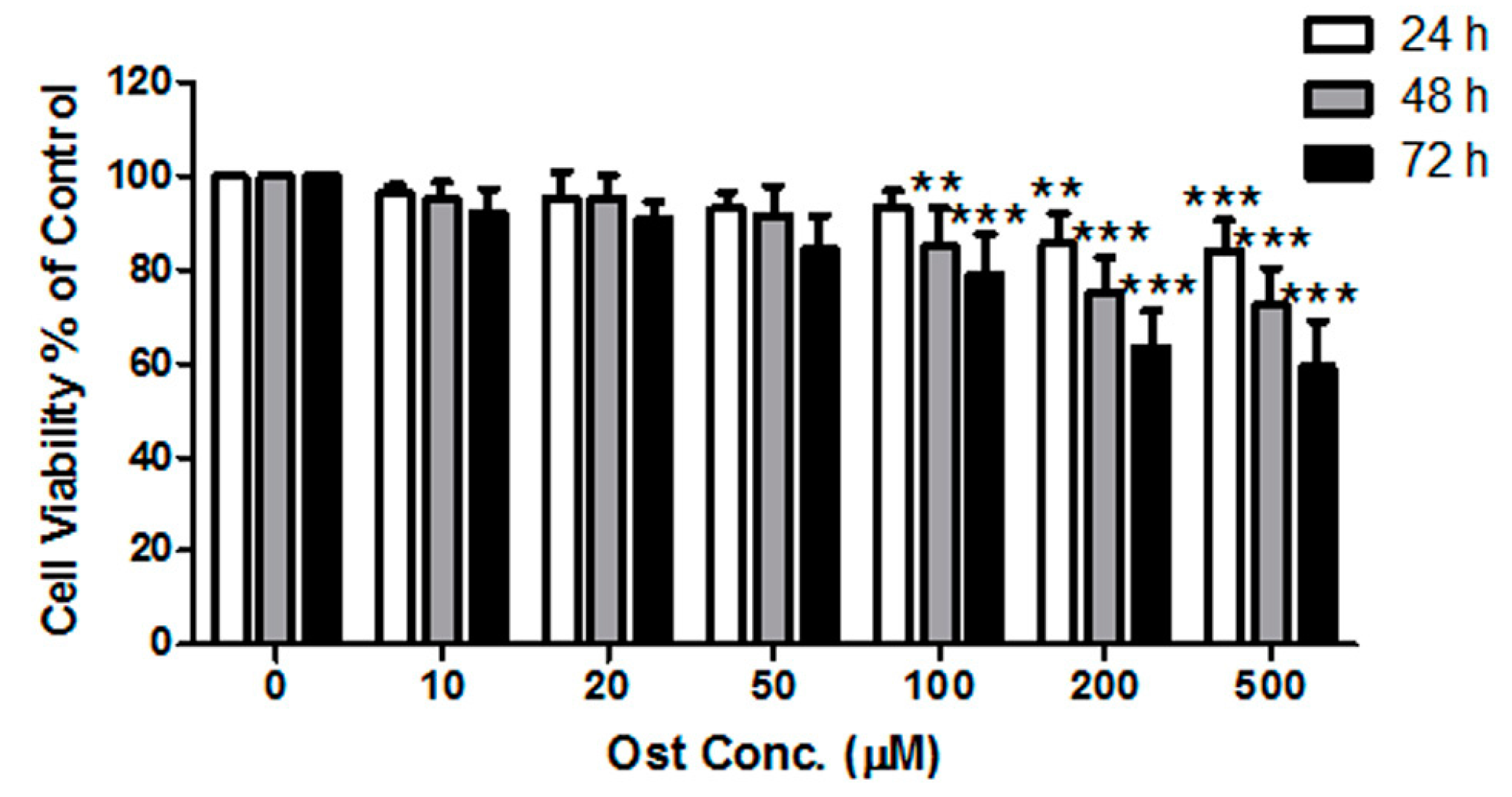
2.3. Cell Viability/Proliferation Assay
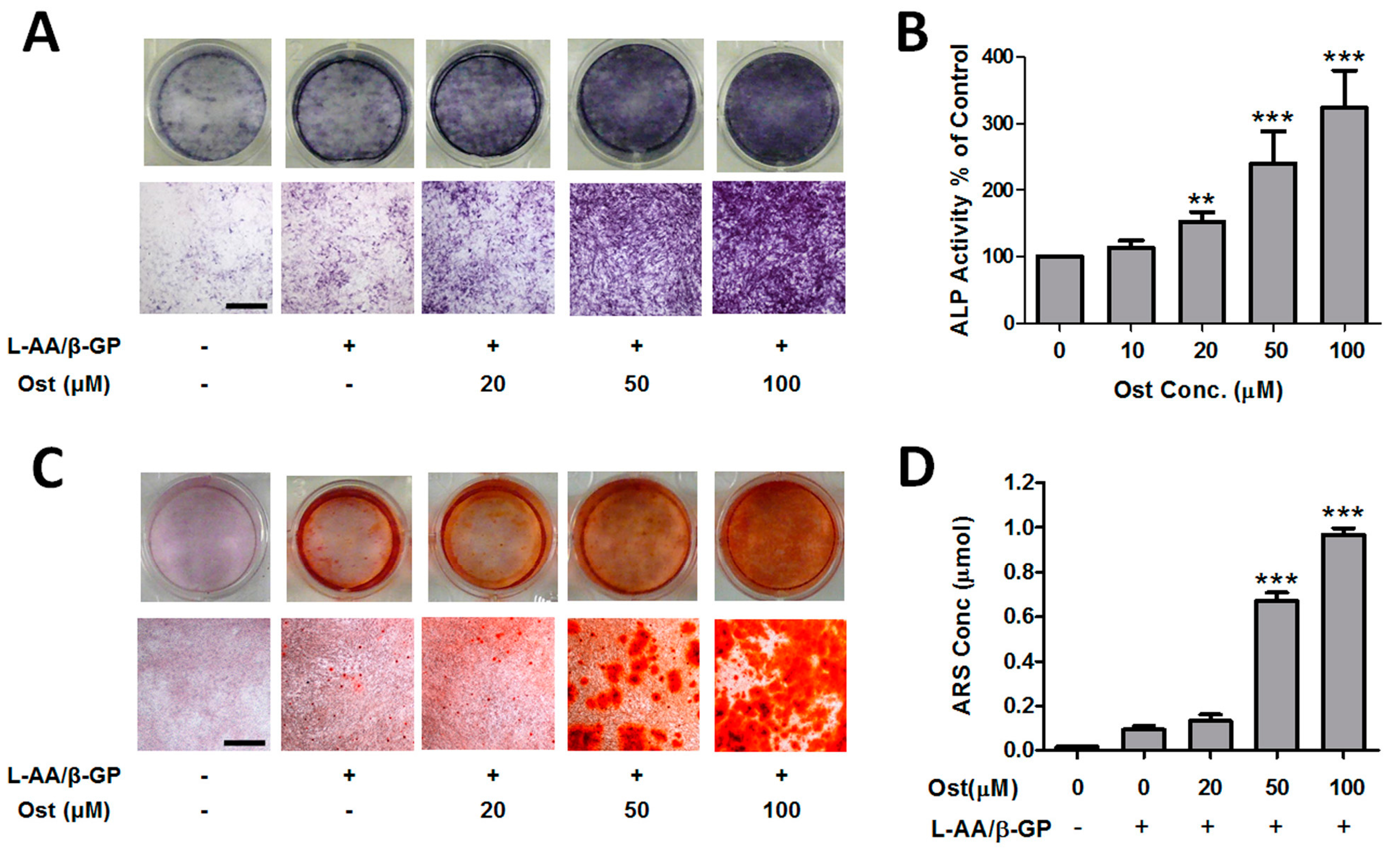
2.4. Alkaline Phosphatase (ALP) Enzyme-Cytochemistry and ALP Activity Assay
2.5. Calcium Nodule Staining and Quantification
2.6. RNA Extraction and Real-Time RT-PCR
2.7. Western Blot
2.8. cAMP Assay
2.9. Histomorphometry and Immunohistochemistry
2.10. Bone Biomechanical Test
2.11. Statistical Analysis
3. Results
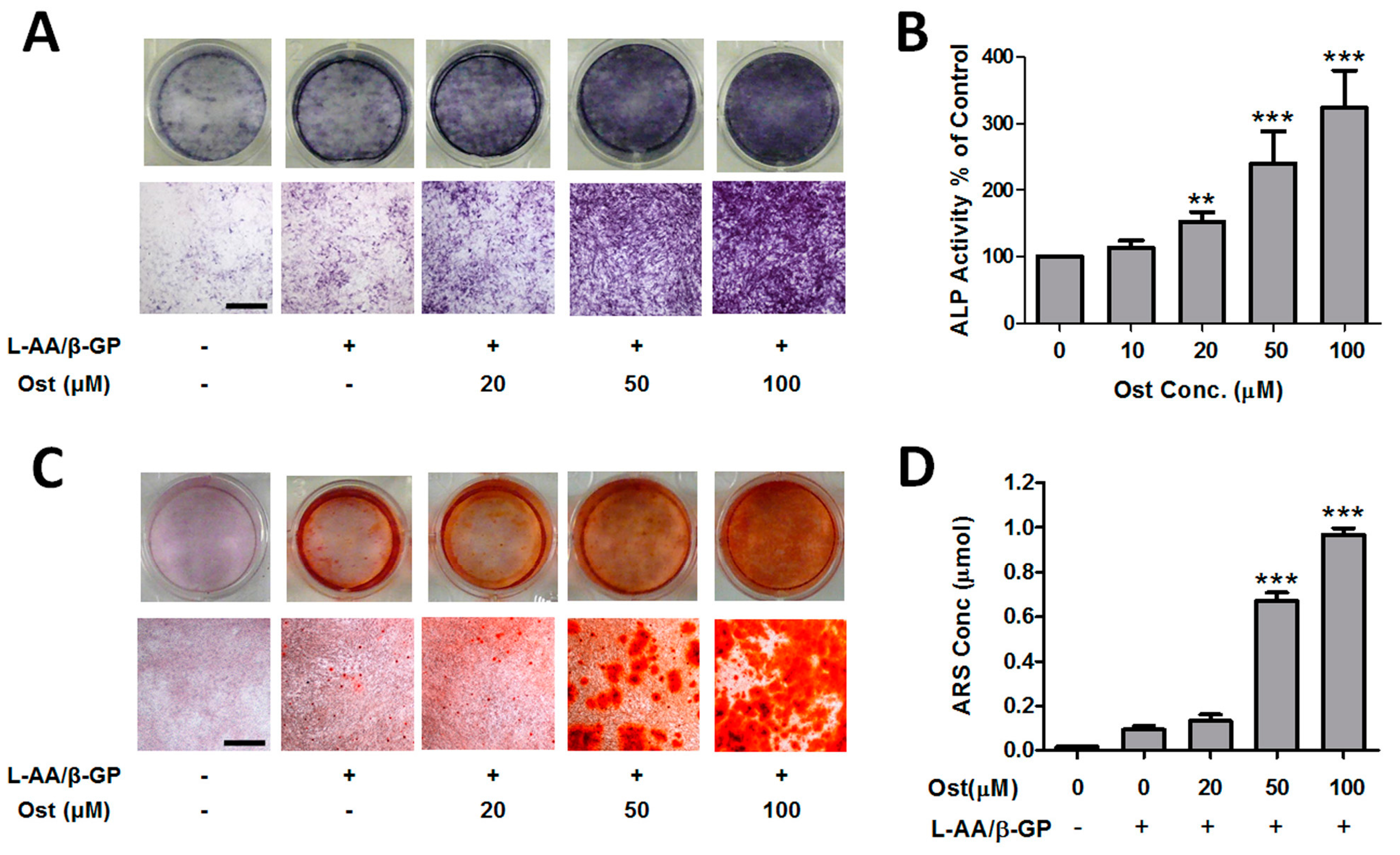
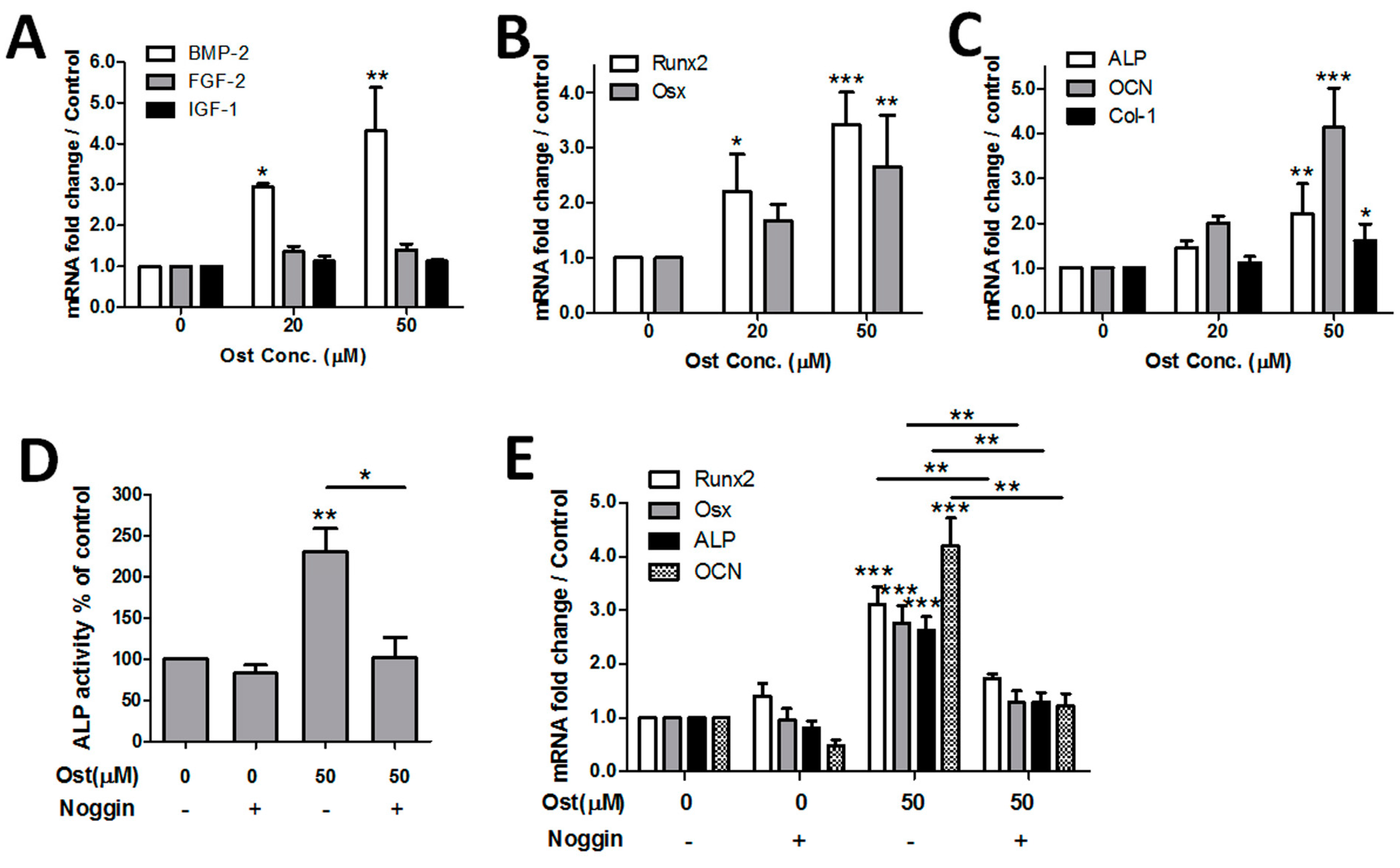
3.1. Osthole Promoted Osteogenesis in Osteoblasts
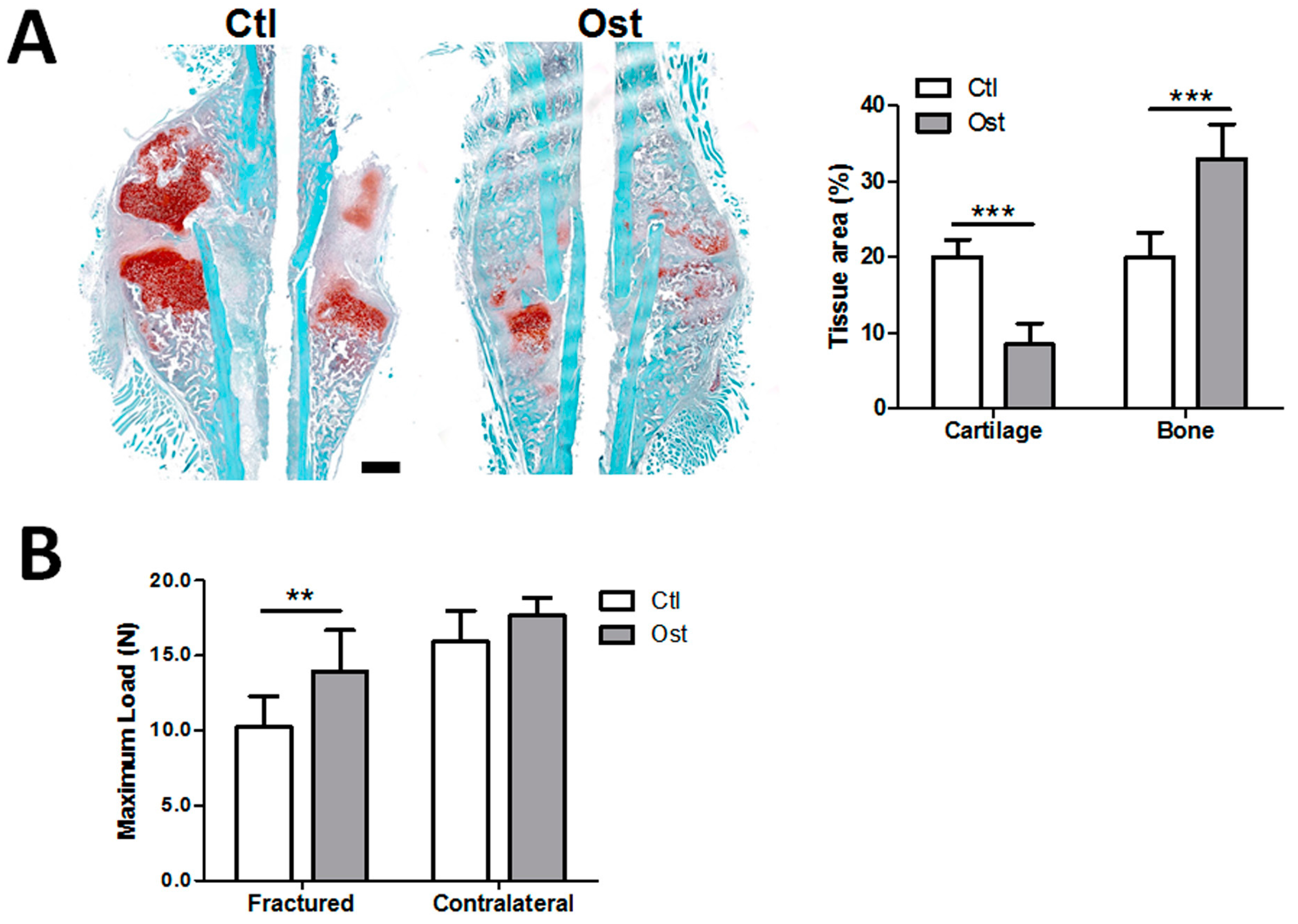
3.2. Osthole Promoted Bone Regeneration and Bone Strength
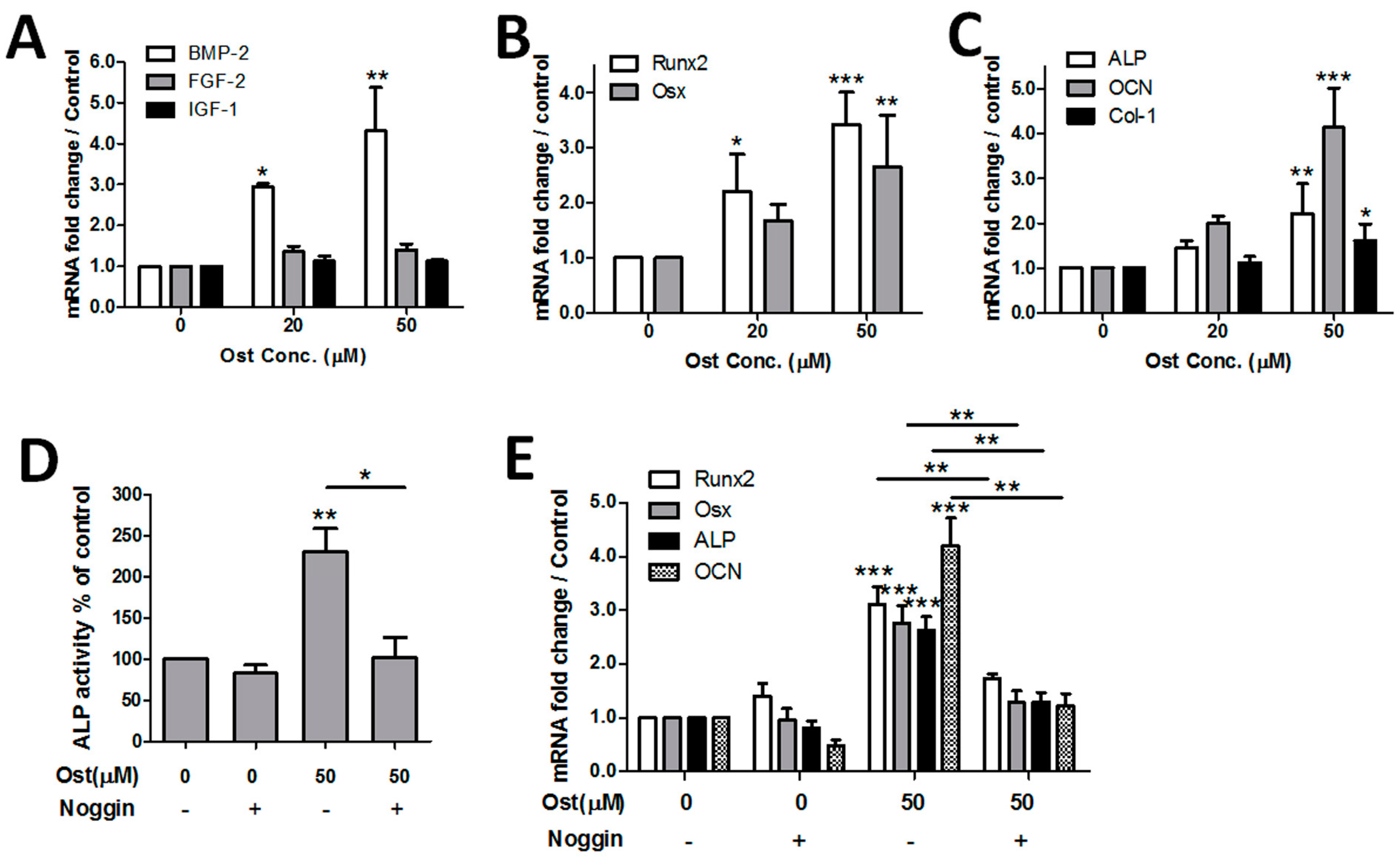
3.3. Osthole Induced Osteogenesis via the BMP-Dependent Signaling Pathway
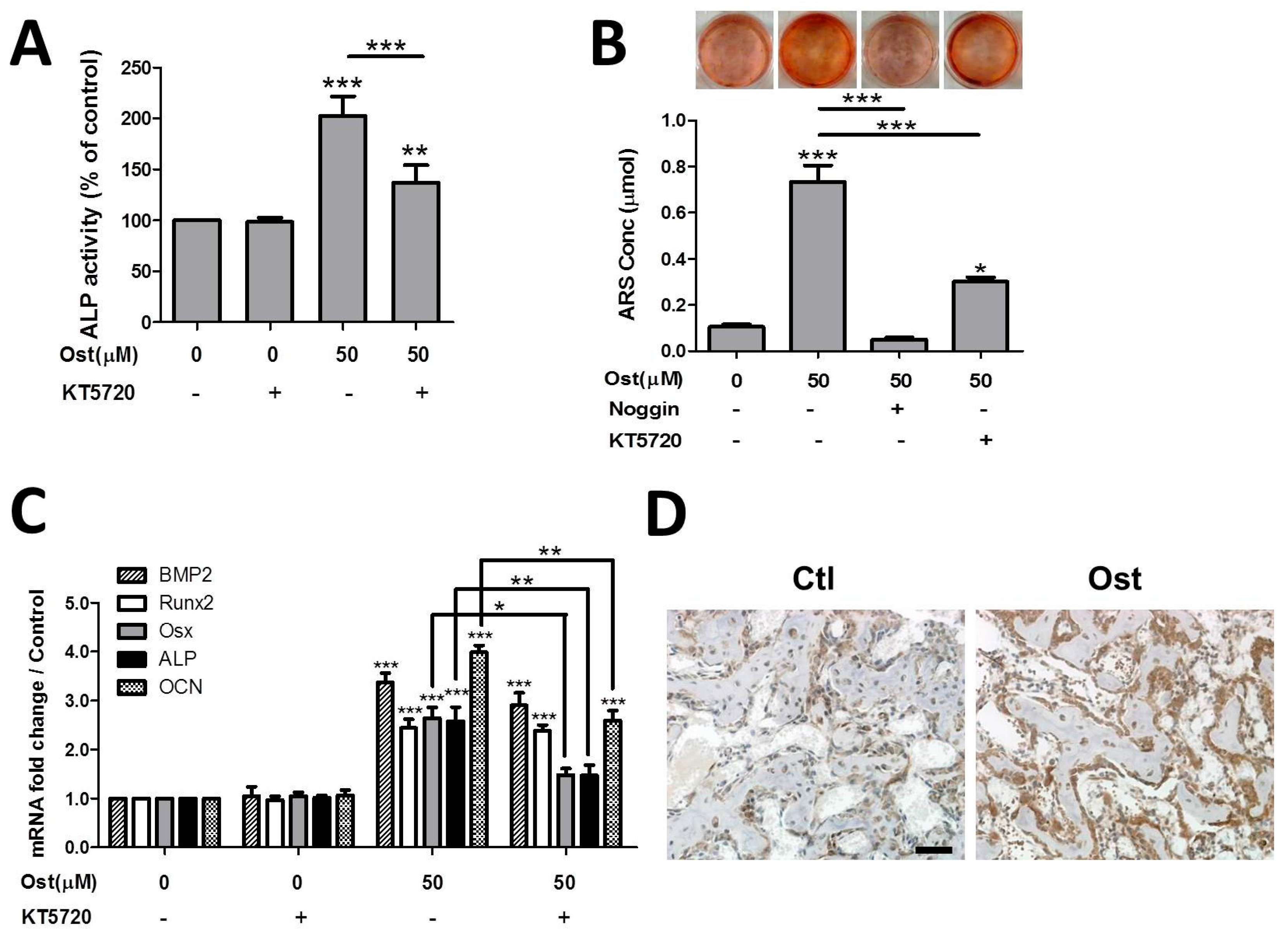
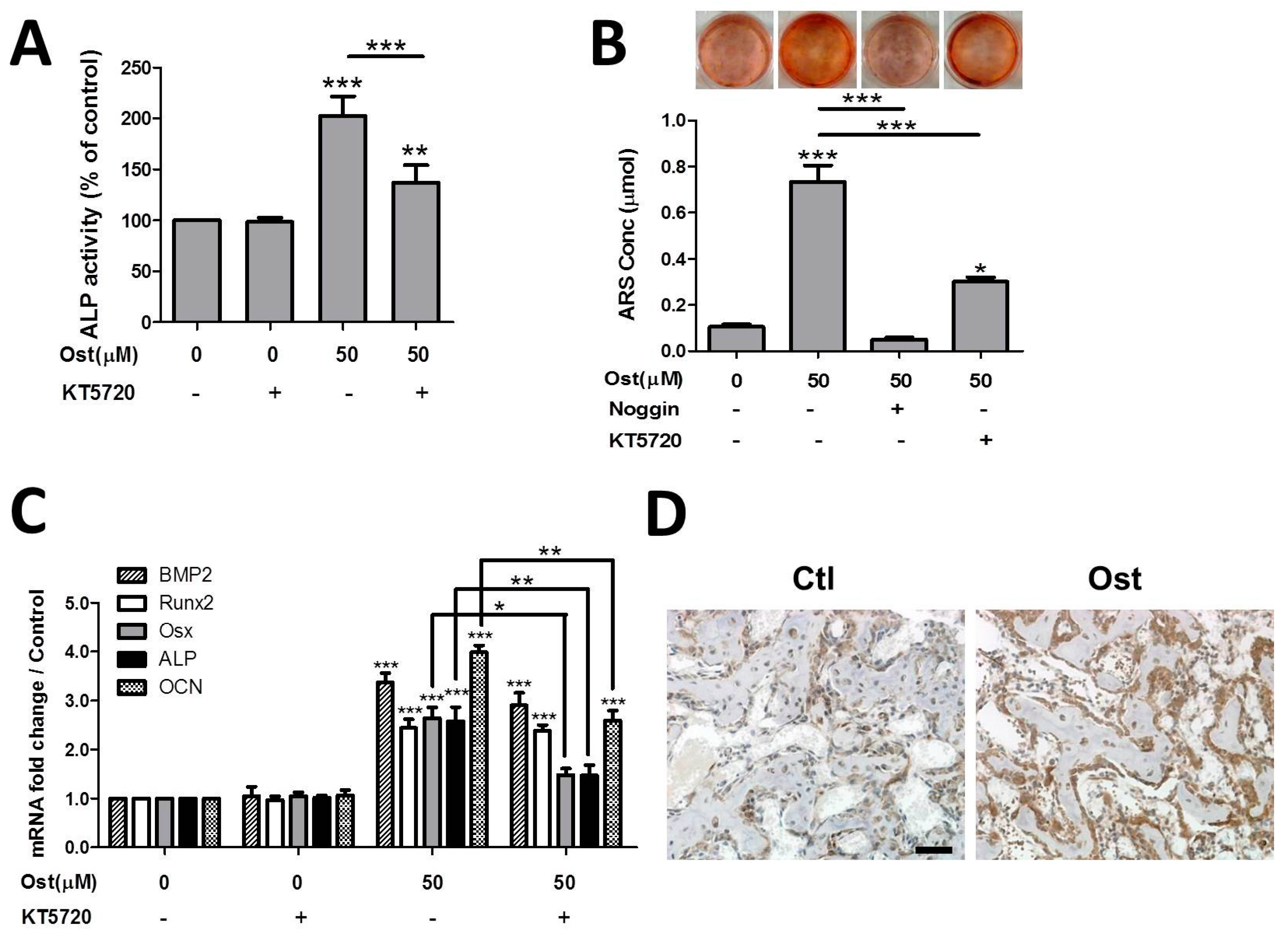
3.4. Osthole Activated cAMP/CREB Signaling Pathway
3.5. Osthole Enhanced Osteogenesis through Osterix Activiated by cAMP/CREB Signaling
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Raisz, L.G. Pathogenesis of osteoporosis: Concepts, conflicts, and prospects. J. Clin. Investig. 2005, 115, 3318–3325. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, P.; Cooper, C. Osteoporosis. Lancet 2006, 367, 2010–2018. [Google Scholar] [CrossRef]
- Nikolaou, V.S.; Efstathopoulos, N.; Kontakis, G.; Kanakaris, N.K.; Giannoudis, P.V. The influence of osteoporosis in femoral fracture healing time. Injury 2009, 40, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Kyllonen, L.; D’Este, M.; Alini, M.; Eglin, D. Local drug delivery for enhancing fracture healing in osteoporotic bone. Acta Biomater. 2015, 11, 412–434. [Google Scholar] [CrossRef] [PubMed]
- Rothberg, D.L.; Lee, M.A. Internal fixation of osteoporotic fractures. Curr. Osteoporos. Rep. 2015, 13, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.W.; Qin, L.; Lee, K.M.; Zhang, M.; Cheng, J.C.; Leung, K.S. Low intensity pulsed ultrasound accelerated bone remodeling during consolidation stage of distraction osteogenesis. J. Orthop. Res. 2006, 24, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.W.; Qin, L.; Lee, K.M.; Cheung, W.H.; Cheng, J.C.; Leung, K.S. Dose-dependent effect of low-intensity pulsed ultrasound on callus formation during rapid distraction osteogenesis. J. Orthop. Res. 2006, 24, 2072–2079. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Lu, X.; Wang, K.; Yuan, H.; Fang, L.; Zheng, X.; Chan, C.; Ren, F.; Zhao, C. Pulse electrochemical driven rapid layer-by-layer assembly of polydopamine and hydroxyapatite nanofilms via alternative redox in situ synthesis for bone regeneration. ACS Biomater. Sci. Eng. 2016, 2, 920–928. [Google Scholar] [CrossRef]
- Hardcastle, A.C.; Aucott, L.; Fraser, W.D.; Reid, D.M.; Macdonald, H.M. Dietary patterns, bone resorption and bone mineral density in early post-menopausal scottish women. Eur. J. Clin. Nutr. 2011, 65, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.S.; Kazmier, P.; Burd, T.A.; Anglen, J.; Stoker, A.M.; Kuroki, K.; Carson, W.L.; Cook, J.L. Enhanced fracture and soft-tissue healing by means of anabolic dietary supplementation. J. Bone Joint Surg. Am. 2006, 88, 2386–2394. [Google Scholar] [CrossRef] [PubMed]
- Yaman, F.; Acikan, I.; Dundar, S.; Simsek, S.; Gul, M.; Ozercan, I.H.; Komorowski, J.; Sahin, K. Dietary arginine silicate inositol complex increased bone healing: Histologic and histomorphometric study. Drug Des. Dev. Ther. 2016, 10, 2081–2086. [Google Scholar] [CrossRef] [PubMed]
- Welch, A.A.; Hardcastle, A.C. The effects of flavonoids on bone. Curr. Osteoporos. Rep. 2014, 12, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Qin, L.; Sheng, H.; Yeung, K.W.; Yeung, H.Y.; Cheung, W.H.; Griffith, J.; Chan, C.W.; Lee, K.M.; Leung, K.S. Epimedium-derived phytoestrogen exert beneficial effect on preventing steroid-associated osteonecrosis in rabbits with inhibition of both thrombosis and lipid-deposition. Bone 2007, 40, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Liu, M.; Sun, L.; Rui, F. Icariin recovers the osteogenic differentiation and bone formation of bone marrow stromal cells from a rat model of estrogen deficiency-induced osteoporosis. Mol. Med. Rep. 2015, 12, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Gu, D.R.; So, H.S.; Kim, K.J.; Lee, S.H. Dual role of cyanidin-3-glucoside on the differentiation of bone cells. J. Dent. Res. 2015, 94, 1676–1683. [Google Scholar] [CrossRef] [PubMed]
- Dou, C.; Cao, Z.; Ding, N.; Hou, T.; Luo, F.; Kang, F.; Yang, X.; Jiang, H.; Xie, Z.; Hu, M.; et al. Cordycepin prevents bone loss through inhibiting osteoclastogenesis by scavenging ros generation. Nutrients 2016, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.R.; Leung, W.N.; Cheung, H.Y.; Chan, C.W. Osthole: A review on its bioactivities, pharmacological properties, and potential as alternative medicine. Evid. Based Complement. Alternat. Med. 2015, 2015, 919616. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Yang, H.; Zhang, Q.; Liu, C.; Zhao, J.; Zhang, L.; Chen, B. Natural products for treatment of osteoporosis: The effects and mechanisms on promoting osteoblast-mediated bone formation. Life Sci. 2016, 147, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Hara, I.; Matsumiya, T. Effects of osthole on postmenopausal osteoporosis using ovariectomized rats; comparison to the effects of estradiol. Biol. Pharm. Bull. 2002, 25, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.Z.; Hou, W.; Zhou, Q.; Zhang, M.; Holz, J.; Sheu, T.J.; Li, T.F.; Cheng, S.D.; Shi, Q.; Harris, S.E.; et al. Osthole stimulates osteoblast differentiation and bone formation by activation of beta-catenin-BMP signaling. J. Bone Miner. Res. 2010, 25, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Leung, W.N.; Li, G.; Lai, Y.M.; Chan, C.W. Osthole promotes endochondral ossification and accelerates fracture healing in mice. Calcif. Tissue Int. 2016, 99, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Rodan, G.A.; Martin, T.J. Therapeutic approaches to bone diseases. Science 2000, 289, 1508–1514. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, S.K.; Hampson, G. The pathogenesis, diagnosis, investigation and management of osteoporosis. J. Clin. Pathol. 2011, 64, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Lane, N.E.; Kelman, A. A review of anabolic therapies for osteoporosis. Arthritis Res. Ther. 2003, 5, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.L.; Hsu, Y.L.; Chang, C.H.; Chang, J.K. Osthole-mediated cell differentiation through bone morphogenetic protein-2/p38 and extracellular signal-regulated kinase 1/2 pathway in human osteoblast cells. J. Pharmacol. Exp. Ther. 2005, 314, 1290–1299. [Google Scholar] [CrossRef] [PubMed]
- Ming, L.G.; Zhou, J.; Cheng, G.Z.; Ma, H.P.; Chen, K.M. Osthol, a coumarin isolated from common cnidium fruit, enhances the differentiation and maturation of osteoblasts in vitro. Pharmacology 2011, 88, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Partridge, N.C. Stimulation of amphiregulin expression in osteoblastic cells by parathyroid hormone requires the protein kinase a and camp response element-binding protein signaling pathway. J. Cell. Biochem. 2005, 96, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Singh, R.; Divieti, P.; Guo, J.; Bouxsein, M.L.; Bringhurst, F.R. Contributions of parathyroid hormone (PTH)/PTH-related peptide receptor signaling pathways to the anabolic effect of pth on bone. Bone 2007, 40, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Edwards, J.R.; Ko, S.Y.; Dong, S.; Liu, H.; Oyajobi, B.O.; Papasian, C.; Deng, H.W.; Zhao, M. Transcriptional regulation of BMP2 expression by the PTH-CREB signaling pathway in osteoblasts. PLoS ONE 2011, 6, e20780. [Google Scholar] [CrossRef] [PubMed]
- Tyson, D.R.; Swarthout, J.T.; Partridge, N.C. Increased osteoblastic c-fos expression by parathyroid hormone requires protein kinase a phosphorylation of the cyclic adenosine 3′,5′-monophosphate response element-binding protein at serine 133. Endocrinology 1999, 140, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Xie, Z.; Konaka, H.; Sodek, J.; Zhau, H.E.; Chung, L.W. Human osteocalcin and bone sialoprotein mediating osteomimicry of prostate cancer cells: Role of cAMP-dependent protein kinase a signaling pathway. Cancer Res. 2005, 65, 2303–2313. [Google Scholar] [CrossRef] [PubMed]
- Takai, H.; Nakayama, Y.; Kim, D.S.; Arai, M.; Araki, S.; Mezawa, M.; Nakajima, Y.; Kato, N.; Masunaga, H.; Ogata, Y. Androgen receptor stimulates bone sialoprotein (BSP) gene transcription via camp response element and activator protein 1/glucocorticoid response elements. J. Cell. Biochem. 2007, 102, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Chiou, W.F.; Huang, Y.L.; Chen, C.F.; Chen, C.C. Vasorelaxing effect of coumarins from cnidium monnieri on rabbit corpus cavernosum. Planta Med. 2001, 67, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.M.; Lin, C.H.; Ko, F.N.; Wu, T.S.; Huang, T.F. The relaxant action of osthole isolated from angelica pubescens in guinea-pig trachea. Naunyn Schmiedebergs Arch. Pharmacol. 1994, 349, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Fang, Z.; Lu, W.; Liu, X.; Zhang, Y. Osthole, a coumadin analog from Cnidium monnieri (L.) cusson, stimulates corticosterone secretion by increasing steroidogenic enzyme expression in mouse y1 adrenocortical tumor cells. J. Ethnopharmacol. 2015, 175, 456–462. [Google Scholar] [CrossRef] [PubMed]
- He, Y.X.; Zhang, G.; Pan, X.H.; Liu, Z.; Zheng, L.Z.; Chan, C.W.; Lee, K.M.; Cao, Y.P.; Li, G.; Wei, L.; et al. Impaired bone healing pattern in mice with ovariectomy-induced osteoporosis: A drill-hole defect model. Bone 2011, 48, 1388–1400. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Xiong, Z.; Sun, Y.; Li, F. Coumarins from Cnidium monnieri (L.) and their proliferation stimulating activity on osteoblast-like umr106 cells. Pharmazie 2004, 59, 643–645. [Google Scholar] [PubMed]
- Zhang, Q.; Qin, L.; He, W.; Van Puyvelde, L.; Maes, D.; Adams, A.; Zheng, H.; De Kimpe, N. Coumarins from cnidium monnieri and their antiosteoporotic activity. Planta Med. 2007, 73, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Damien, C.J.; Parsons, J.R. Bone graft and bone graft substitutes: A review of current technology and applications. J. Appl. Biomater. 1991, 2, 187–208. [Google Scholar] [CrossRef] [PubMed]
- Li, V.C.; Kirschner, M.W. Molecular ties between the cell cycle and differentiation in embryonic stem cells. Proc. Natl. Acad. Sci. USA 2014, 111, 9503–9508. [Google Scholar] [CrossRef] [PubMed]
- Dupree, M.A.; Pollack, S.R.; Levine, E.M.; Laurencin, C.T. Fibroblast growth factor 2 induced proliferation in osteoblasts and bone marrow stromal cells: A whole cell model. Biophys. J. 2006, 91, 3097–3112. [Google Scholar] [CrossRef] [PubMed]
- Radcliff, K.; Tang, T.B.; Lim, J.; Zhang, Z.; Abedin, M.; Demer, L.L.; Tintut, Y. Insulin-like growth factor-i regulates proliferation and osteoblastic differentiation of calcifying vascular cells via extracellular signal-regulated protein kinase and phosphatidylinositol 3-kinase pathways. Circ. Res. 2005, 96, 398–400. [Google Scholar] [CrossRef] [PubMed]
- Siddappa, R.; Martens, A.; Doorn, J.; Leusink, A.; Olivo, C.; Licht, R.; van Rijn, L.; Gaspar, C.; Fodde, R.; Janssen, F.; et al. cAMP/PKA pathway activation in human mesenchymal stem cells in vitro results in robust bone formation in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 7281–7286. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wen, Q.; Liang, W.; Kang, T.; Ren, L.; Zhang, N.; Zhao, D.; Sun, D.; Yang, J. Osthole reverses beta-amyloid peptide cytotoxicity on neural cells by enhancing cyclic amp response element-binding protein phosphorylation. Biol. Pharm. Bull. 2013, 36, 1950–1958. [Google Scholar] [CrossRef] [PubMed]
- Javed, A.; Afzal, F.; Bae, J.S.; Gutierrez, S.; Zaidi, K.; Pratap, J.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. Specific residues of Runx2 are obligatory for formation of BMP2-induced Runx2-smad complex to promote osteoblast differentiation. Cells Tissues Organs 2009, 189, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Kwon, T.G.; Park, H.S.; Wozney, J.M.; Ryoo, H.M. BMP-2-induced osterix expression is mediated by Dlx5 but is independent of Runx2. Biochem. Biophys. Res. Commun. 2003, 309, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; de Crombrugghe, B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef]
- Matsubara, T.; Kida, K.; Yamaguchi, A.; Hata, K.; Ichida, F.; Meguro, H.; Aburatani, H.; Nishimura, R.; Yoneda, T. BMP2 regulates osterix through Msx2 and Runx2 during osteoblast differentiation. J. Biol. Chem. 2008, 283, 29119–29125. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.-R.; Leung, W.N.; Li, G.; Kong, S.K.; Lu, X.; Wong, Y.M.; Chan, C.W. Osthole Enhances Osteogenesis in Osteoblasts by Elevating Transcription Factor Osterix via cAMP/CREB Signaling In Vitro and In Vivo. Nutrients 2017, 9, 588. https://doi.org/10.3390/nu9060588
Zhang Z-R, Leung WN, Li G, Kong SK, Lu X, Wong YM, Chan CW. Osthole Enhances Osteogenesis in Osteoblasts by Elevating Transcription Factor Osterix via cAMP/CREB Signaling In Vitro and In Vivo. Nutrients. 2017; 9(6):588. https://doi.org/10.3390/nu9060588
Chicago/Turabian StyleZhang, Zhong-Rong, Wing Nang Leung, Gang Li, Siu Kai Kong, Xiong Lu, Yin Mei Wong, and Chun Wai Chan. 2017. "Osthole Enhances Osteogenesis in Osteoblasts by Elevating Transcription Factor Osterix via cAMP/CREB Signaling In Vitro and In Vivo" Nutrients 9, no. 6: 588. https://doi.org/10.3390/nu9060588