Feeding a Mixture of Choline Forms during Lactation Improves Offspring Growth and Maternal Lymphocyte Response to Ex Vivo Immune Challenges

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Tissue Collection
2.3. Choline Metabolite Analyses of Dam’s Splenocytes and Offspring Stomach Content
2.4. Immune Cell Isolation
2.5. Immune Cell PhenotyPe Analysis
2.6. Ex Vivo Cytokine Production by Mitogen-Stimulated Cells
2.7. Statistical Analyses
3. Results
3.1. AnthroPometric Characteristics and Daily Food Intake
3.2. Choline Metabolites in Pup’s Stomach Content and Dams’s SPleen
3.3. Splenocyte Phenotypes
3.4. Mesenteric Lymphocyte Phenotypes
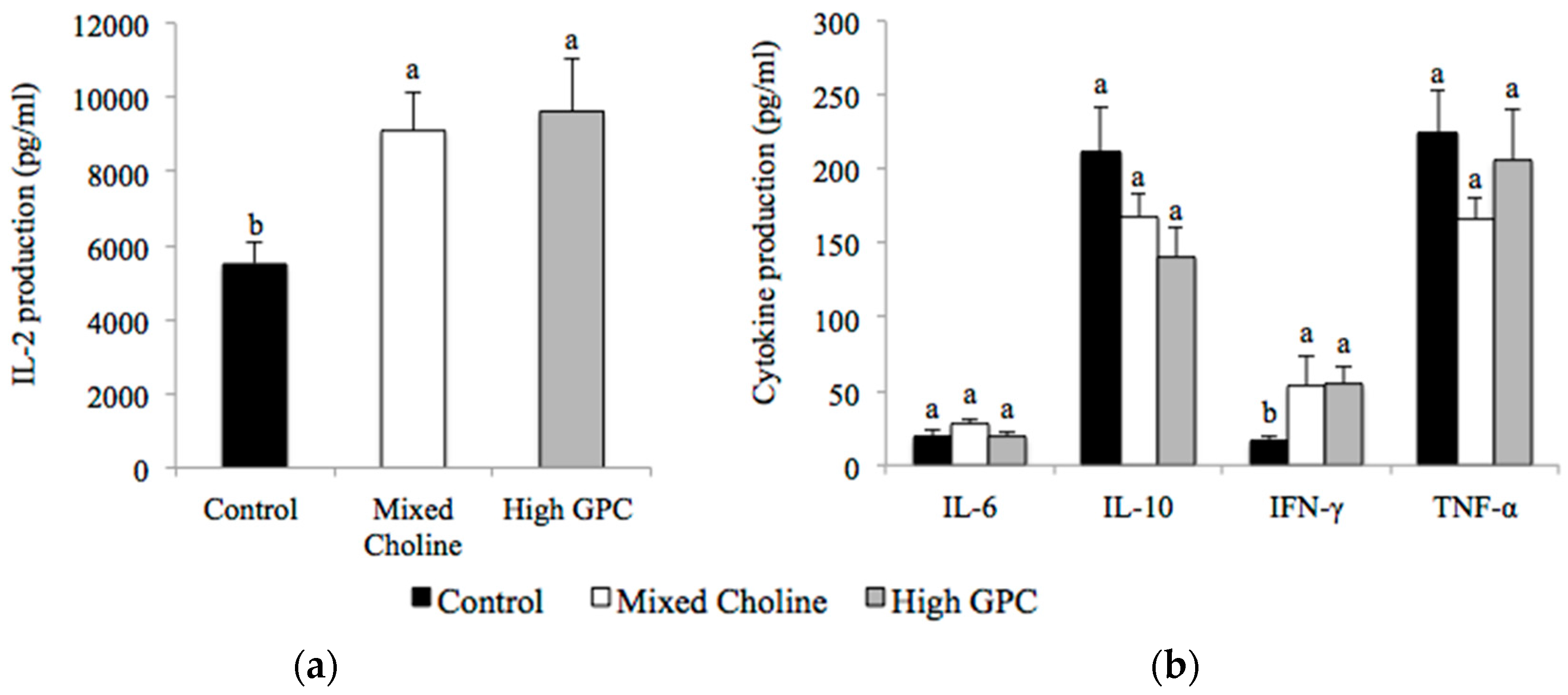
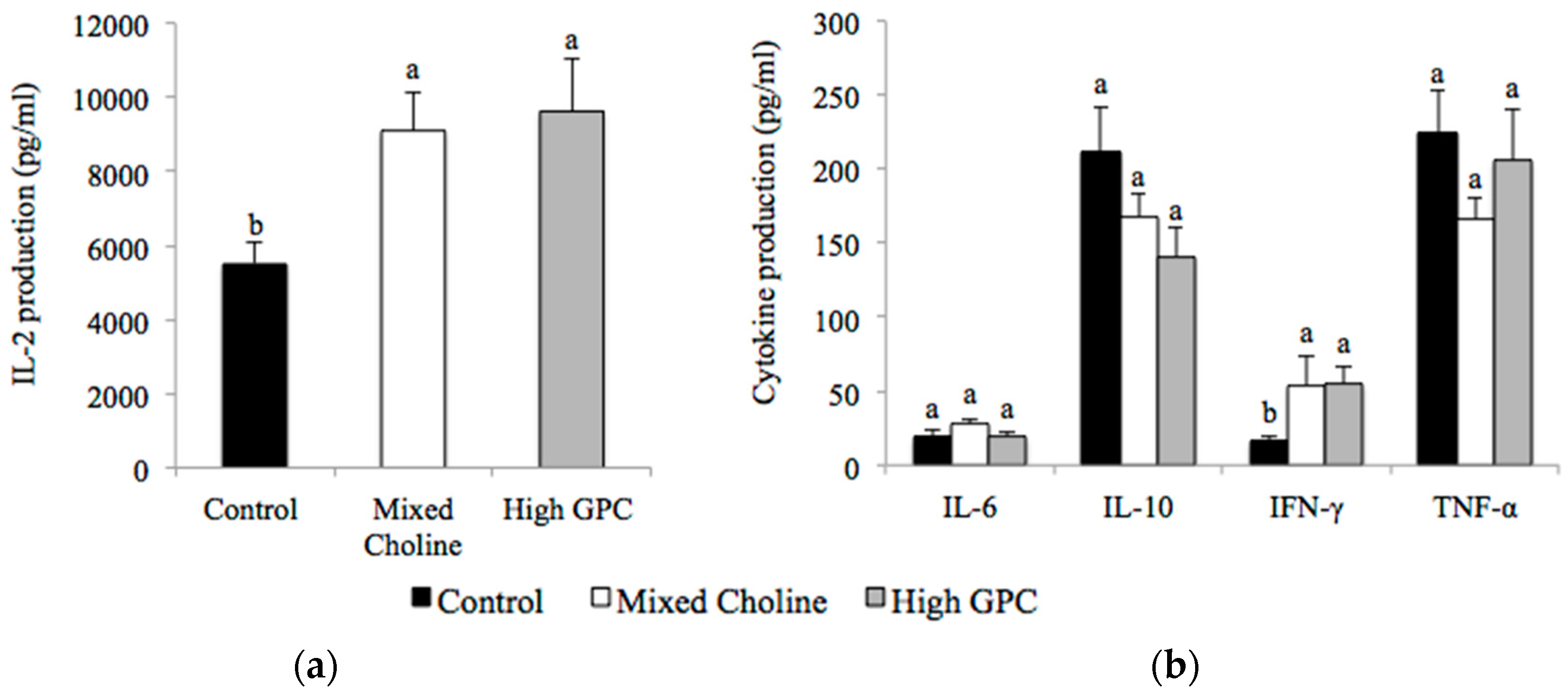
3.5. Ex Vivo Cytokine Production after Stimulation
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dellschaft, N.S.; Ruth, M.R.; Goruk, S.; Lewis, E.D.; Richard, C.; Jacobs, R.L.; Curtis, J.M.; Field, C.J. Choline is required in the diet of lactating dams to maintain maternal immune function. Br. J. Nutr. 2015, 113, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.D.; Goruk, S.; Richard, C.; Dellschaft, N.S.; Curtis, J.M.; Jacobs, R.L.; Field, C.J. Feeding a diet devoid of choline to lactating rodents restricts growth and lymphocyte development in offspring. Br. J. Nutr. 2016, 116, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, R.P.; Kelly, K.B.; Lewis, E.D.; Leonard, K.A.; Goruk, S.; Curtis, J.M.; Vine, D.F.; Proctor, S.D.; Field, C.J.; Jacobs, R.L. Choline deficiency impairs intestinal lipid metabolism in the lactating rat. J. Nutr. Biochem. 2015, 26, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.D.; Subhan, F.B.; Bell, R.C.; McCargar, L.J.; Curtis, J.M.; Jacobs, R.L.; Field, C.J. Estimation of choline intake from 24 h dietary intake recalls and contribution of egg and milk consumption to intake among pregnant and lactating women in Alberta. Br. J. Nutr. 2014, 112, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Mygind, V.L.; Evans, S.E.; Peddie, M.C.; Miller, J.C.; Houghton, L.A. Estimation of usual intake and food sources of choline and betaine in New Zealand reproductive age women. Asia Pac. J. Clin. Nutr. 2013, 22, 319–324. [Google Scholar] [PubMed]
- Yonemori, K.M.; Lim, U.; Koga, K.R.; Wilkens, L.R.; Au, D.; Boushey, C.J.; Le Marchand, L.; Kolonel, L.N.; Murphy, S.P. Dietary choline and betaine intakes vary in an adult multiethnic population. J. Nutr. 2013, 143, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G. Components of the ain-93 diets as improvements in the ain-76a diet. J. Nutr. 1997, 127, 838S–841S. [Google Scholar] [PubMed]
- Holmes-McNary, M.Q.; Cheng, W.L.; Mar, M.H.; Fussell, S.; Zeisel, S.H. Choline and choline esters in human and rat milk and in infant formulas. Am. J. Clin. Nutr. 1996, 64, 572–576. [Google Scholar] [PubMed]
- Kawashima, K.; Fujii, T.; Moriwaki, Y.; Misawa, H. Critical roles of acetylcholine and the muscarinic and nicotinic acetylcholine receptors in the regulation of immune function. Life Sci. 2012, 91, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- James, S.J.; Yin, L. Diet-induced DNA damage and altered nucleotide metabolism in lymphocytes from methyl-donor-deficient rats. Carcinogenesis 1989, 10, 1209–1214. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.D.; Richard, C.; Goruk, S.; Dellschaft, N.S.; Curtis, J.M.; Jacobs, R.L.; Field, C.J. The form of choline in the maternal diet affects immune development in suckled rat offspring. J. Nutr. 2016, 146, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Dellschaft, N.S.; Richard, C.; Lewis, E.D.; Goruk, S.; Jacobs, R.L.; Curtis, J.M.; Field, C.J. The dietary form of choline during lactation affects maternal immune function in rats. Eur. J. Nutr. 2017. [Google Scholar] [CrossRef]
- Patterson, K.Y.; Bhagwat, S.A.; Williams, J.R.; Howe, J.R.; Holden, J.M.; Zeisel, S.H.; Dacosta, K.A.; Mar, M.-H. USDA Database for the Choline Content of Common Foods Release Two; Nutrient Data Laboratory, Beltsville Human Nutrition Research Center, ARS, USDA: Beltsville, MD, USA, 2008. [Google Scholar]
- Richard, C.; Lewis, E.D.; Zhao, Y.Y.; Asomaning, J.; Jacobs, R.L.; Field, C.J.; Curtis, J.M. Measurement of the total choline content in 48 commercial dairy products or dairy alternatives. J. Food Compos. Anal. 2016, 45, 1–8. [Google Scholar] [CrossRef]
- Burg, M.B.; Ferraris, J.D. Intracellular organic osmolytes: Function and regulation. J. Biol. Chem. 2008, 283, 7309–7313. [Google Scholar] [CrossRef] [PubMed]
- Konger, R.L.; Marathe, G.K.; Yao, Y.; Zhang, Q.; Travers, J.B. Oxidized glycerophosphocholines as biologically active mediators for ultraviolet radiation-mediated effects. Prostaglandins Other Lipid Mediat. 2008, 87, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Nagase, T.; Shimizu, T. Platelet-activating factor receptor. Prostaglandins Other Lipid Mediat. 2002, 68–69, 599–609. [Google Scholar] [CrossRef]
- Richard, C.; Lewis, E.D.; Goruk, S.; Wadge, E.; Curtis, J.M.; Jacobs, R.L.; Field, C.J. Feeding a mixture of choline forms to lactating dams improves the development of the immune system in sprague-dawley rat offspring. Nutrients 2017, 9, 567. [Google Scholar] [CrossRef] [PubMed]
- Bernhart, F.W.; Tomarelli, R.M. A salt mixture supplying the national research council estimates of the mineral requirements of the rat. J. Nutr. 1966, 89, 495–500. [Google Scholar] [PubMed]
- Kaplan, B.J.; Giesbrecht, G.F.; Leung, B.M.; Field, C.J.; Dewey, D.; Bell, R.C.; Manca, D.P.; O'Beirne, M.; Johnston, D.W.; Pop, V.J.; et al. The alberta pregnancy outcomes and nutrition (APrON) cohort study: Rationale and methods. Matern. Child. Nutr. 2014, 10, 44–60. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Zhao, Y.Y.; Goruk, S.; Oilund, K.; Field, C.J.; Jacobs, R.L.; Curtis, J.M. Validation of an LC-MS/MS method for the quantification of choline-related compounds and phospholipids in foods and tissues. J. Chromatogr. B Biomed. Sci. Appl. 2012, 911, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; Xiong, Y.; Curtis, J.M. Measurement of phospholipids by hydrophilic interaction liquid chromatography coupled to tandem mass spectrometry: The determination of choline containing compounds in foods. J. Chromatogr. A 2011, 1218, 5470–5479. [Google Scholar] [CrossRef] [PubMed]
- Field, C.J.; Wu, G.; Metroz-Dayer, M.D.; Montambault, M.; Marliss, E.B. Lactate production is the major metabolic fate of glucose in splenocytes and is altered in spontaneously diabetic BB rats. Biochem. J. 1990, 272, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Field, C.J.; Thomson, C.A.; van Aerde, J.E.; Parrott, A.; Euler, A.; Lien, E.; Clandinin, M.T. Lower proportion of CD45R0+ cells and deficient interleukin-10 production by formula-fed infants, compared with human-fed, is corrected with supplementation of long-chain polyunsaturated fatty acids. J. Pediatr. Gastroenterol. Nutr. 2000, 31, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Blewett, H.J.; Gerdung, C.A.; Ruth, M.R.; Proctor, S.D.; Field, C.J. Vaccenic acid favourably alters immune function in obese JCR:LA-cp rats. Br. J. Nutr. 2009, 102, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Moukarzel, S.; Soberanes, L.; Dyer, R.A.; Albersheim, S.; Elango, R.; Innis, S.M. Relationships among different water-soluble choline compounds differ between human preterm and donor milk. Nutrients 2017, 9, 369. [Google Scholar] [CrossRef] [PubMed]
- Schoenborn, J.R.; Wilson, C.B. Regulation of interferon-gamma during innate and adaptive immune responses. Adv. Immunol. 2007, 96, 41–101. [Google Scholar] [PubMed]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Couper, K.N.; Blount, D.G.; Riley, E.M. Il-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef] [PubMed]
- Kohler, S.; Thiel, A.; Rudwaleit, M.; Sieper, J.; Braun, J. CD27+ memory and CD27- effector CD8+ T cells are responsible for a decreased production of proinflammatory cytokines in HLA B27-positive subjects. Clin. Exp. Rheumatol. 2005, 23, 840–846. [Google Scholar] [PubMed]
- Reddy, M.; Eirikis, E.; Davis, C.; Davis, H.M.; Prabhakar, U. Comparative analysis of lymphocyte activation marker expression and cytokine secretion profile in stimulated human peripheral blood mononuclear cell cultures: An in vitro model to monitor cellular immune function. J. Immunol. Methods 2004, 293, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Buettner, M.; Bode, U. Lymph node dissection—understanding the immunological function of lymph nodes. Clin. Exp. Immunol. 2012, 169, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Newberry, R.D.; Lorenz, R.G. Organizing a mucosal defense. Immunol. Rev. 2005, 206, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Lin, J.X.; Leonard, W.J. Interleukin-2 at the crossroads of effector responses, tolerance, and immunotherapy. Immunity 2013, 38, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Malek, T.R. The main function of IL-2 is to promote the development of T regulatory cells. J. Leukoc. Biol. 2003, 74, 961–965. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Harrison, O.J.; Powrie, F.M. Regulatory T cells and immune tolerance in the intestine. Cold Spring Harb. Perspect. Biol. 2013, 5, a018341. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Immunological parameters: What do they mean? J. Nutr. 2007, 137, 773S–780S. [Google Scholar] [PubMed]
- Kasahara, T.; Hooks, J.J.; Dougherty, S.F.; Oppenheim, J.J. Interleukin 2-mediated immune interferon (IFN-gamma) production by human T cells and T cell subsets. J. Immunol. 1983, 130, 1784–1789. [Google Scholar] [PubMed]
- Foulds, L.M.; Boysen, R.I.; Crane, M.; Yang, Y.; Muir, J.A.; Smith, A.I.; de Kretser, D.M.; Hearn, M.T.; Hedger, M.P. Molecular identification of lyso-glycerophosphocholines as endogenous immunosuppressives in bovine and rat gonadal fluids. Biol. Reprod. 2008, 79, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Tokes, T.; Tuboly, E.; Varga, G.; Major, L.; Ghyczy, M.; Kaszaki, J.; Boros, M. Protective effects of L-alpha-glycerylphosphorylcholine on ischaemia-reperfusion-induced inflammatory reactions. Eur. J. Nutr. 2015, 54, 109–118. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Component (g/kg Diet) | Control Diet | MC Diet | HGPC Diet |
|---|---|---|---|
| Casein | 270 | 270 | 270 |
| Starch | 240 | 240 | 240 |
| Sucrose | 126 | 126 | 126 |
| Vitamin mix (AIN-93-Vx) 2 | 19 | 19 | 19 |
| Mineral mix 3 | 50 | 50 | 50 |
| Calcium phosphate dibasic | 3.4 | 3.4 | 3.4 |
| Inositol | 6.3 | 6.3 | 6.3 |
| Cellulose | 80.0 | 80.0 | 80.0 |
| l-cysteine | 1.8 | 1.8 | 1.8 |
| Fat mixture | |||
| Canola oil | 40 | 31 | 39 |
| Olive oil | 13 | 13.8 | 8.5 |
| Vegetable oil | 14 | 14 | 14.3 |
| Corn oil | 73 | 65 | 71.9 |
| Sunflower oil | 2 | 5 | 2.8 |
| Flax seed oil | 2 | 2 | 2 |
| Hydrogenated canola oil | 56 | 57.5 | 56 |
| DHAsco | 1.5 | 1.5 | 1.5 |
| ARAsco | 1.5 | 1.5 | 1.5 |
| Choline mixture (providing 1 g of total choline/kg diet) | |||
| Soy lecithin (PC) | 0 | 13.1 | 3.2 |
| Choline bitartrate | 2.1 | 0.6 | 0.3 |
| Glycerophosphocholine | 0.0 | 0.6 | 1.9 |
| Control (n = 6) | MC (n = 6) | HGPC (n = 6) | p Value | |
|---|---|---|---|---|
| Body weight (g) | 302 ± 11 | 322 ± 9 | 318 ± 5 | 0.284 |
| Spleen weight (g) | 0.75 ± 0.02 | 0.81 ± 0.05 | 0.79 ± 0.02 | 0.385 |
| Splenocytes/g spleen (×106) | 275 ± 14.9 | 220 ± 12.1 | 296 ± 20.3 | 0.086 |
| Mesenteric lymphocytes/animal (×106) | 6.3 ± 0.8 | 8.0 ± 0.7 | 6.3 ± 0.8 | 0.169 |
| Liver weight (g) | 14.6 ± 1.4 | 16.7 ± 0.8 | 15.1 ± 0.5 | 0.285 |
| Intestine length (cm) | 139 ± 4 | 145 ± 3 | 141 ± 2 | 0.339 |
| Food Intake (g/day) | 47 ± 3 | 46 ± 1 | 47 ± 1 | 0.395 |
| Average pup weight (g) | 60 ± 3 b | 74 ± 3 a | 72 ± 3 a | 0.007 |
| Choline-Containing Metabolite (µg/mg Protein) | Control (n = 6) | MC (n = 6) | HGPC (n = 6) | p Value |
|---|---|---|---|---|
| PC | 19.3 ± 1.6 | 18.4 ± 1.8 | 15.4 ± 1.4 | 0.205 |
| Free choline | 0.45 ± 0.04 | 0.43 ± 0.04 | 0.56 ± 0.07 | 0.232 |
| GPC | 0.30 ± 0.03 | 0.27 ± 0.04 | 0.52 ± 0.13 | 0.084 |
| LysoPC | 0.36 ± 0.04 b | 1.10 ± 0.1 a | 1.07 ± 0.1 a | 0.0001 |
| Sphingomyelin | 3.4 ± 0.3 | 4.1 ± 0.4 | 4.2 ± 0.3 | 0.176 |
| Phosphocholine | 1.1 ± 0.04 b | 1.5 ± 0.04 a | 1.3 ± 0.06 b | 0.0015 |
| Total choline (µg/mg protein) | 4.3 ± 0.2 | 4.6 ± 0.3 | 4.3 ± 0.2 | 0.669 |
| Cell Phenotype | Control (n = 6) | MC (n = 6) | HGPC (n = 6) | p Value |
|---|---|---|---|---|
| % of total lymphocytes | ||||
| Total CD3+ (T cell) | 44.6 ± 1.8 | 40.9 ± 1.8 | 36.9 ± 3.5 | 0.167 |
| % of CD3+ cells | ||||
| CD4+ | 59.3 ± 1.7 a | 57.5 ± 4.6 a | 51.1 ± 2.7 b | 0.011 |
| CD8+ | 34.8 ± 0.9 | 32.1 ± 1.5 | 36.1 ± 1.7 | 0.164 |
| % of CD4+ T cells | ||||
| CD27+ | 73.7 ± 9.5 | 81.8 ± 2.7 | 86.3 ± 2.8 | 0.342 |
| CD28+ | 84.2 ± 6.1 b | 98.9 ± 0.6 a | 94.7 ± 1.7 ab | 0.035 |
| CD71+ | 6.9 ± 1.1 b | 11.1 ± 1.0 a | 13.4 ± 1.2 a | 0.041 |
| CD127+ | 2.9 ± 0.5 | 2.1 ± 0.2 | 2.6 ± 0.6 | 0.558 |
| % of CD8+ T cells | ||||
| CD27+ | 79.3 ± 8.1 b | 98.2 ± 1.3 a | 99.7 ± 0.2 a | 0.013 |
| CD28+ | 71.7 ± 6.3 | 74.1 ± 1.4 | 69.1 ± 2.4 | 0.683 |
| CD71+ | 7.9 ± 0.8 b | 13.2 ± 0.9 ab | 16.4 ± 3.2 a | 0.023 |
| CD127+ | 3.9 ± 0.9 b | 5.1 ± 0.3 a | 8.2 ± 1.2 a | 0.011 |
| % of total lymphocytes | ||||
| CD68+CD284+ | 8.3 ± 0.3 ab | 7.9 ± 0.2 b | 9.0 ± 0.3 a | 0.045 |
| OX62+OX6+ (Dendritic cell) | 4.4 ± 1.1 b | 8.6 ± 0.5 a | 9.2 ± 0.6 a | 0.001 |
| Total CD45RA+ (B cells) | 30.7 ± 1.0 b | 40.4 ± 1.3 a | 39.1 ± 4.7 a | 0.034 |
| CD80+CD45RA+ | 4.8 ± 0.5 a | 2.9 ± 0.2 b | 3.7 ± 0.2 b | 0.004 |
| IgA+ | 12.1 ± 0.6 | 12.1 ± 0.3 | 13.6 ± 0.6 | 0.105 |
| IgG+ | 10.4 ± 1.1 a | 5.4 ± 0.8 b | 3.2 ± 0.2 b | 0.0003 |
| IgM+ | 49.6 ± 1.5 b | 55.2 ± 0.8 a | 53.1 ± 1.9 ab | 0.045 |
| Cell Phenotype | Control (n = 6) | MC (n = 6) | HGPC (n = 6) | p Value |
|---|---|---|---|---|
| % of total lymphocytes | ||||
| Total CD3+ (T cell) | 57.0 ± 1.6 | 58.5 ± 1.1 | 60.3 ± 1.5 | 0.278 |
| % of CD3+ cells | ||||
| CD4+ | 69.3 ± 0.6 | 70.1 ± 1.4 | 66.0 ± 1.4 | 0.061 |
| CD8+ | 26.0 ± 1.6 | 26.5 ± 3.8 | 22.4 ± 3.7 | 0.628 |
| % of CD4+ T cells | ||||
| CD71+ | 6.2 ± 0.9 | 5.5 ± 0.8 | 6.8 ± 0.6 | 0.484 |
| CD127+ | 0.73 ± 0.2 | 1.5 ± 0.3 | 2.7 ± 1.2 | 0.215 |
| CD152+ (CTLA-4+) | 1.8 ± 1.1 | 0.8 ± 0.1 | 2.4 ± 0.9 | 0.433 |
| % of CD8+ T cells | ||||
| CD71+ | 7.0 ± 0.7 | 7.9 ± 1.3 | 5.7 ± 0.5 | 0.258 |
| CD127+ | 1.0 ± 0.2 | 1.1 ± 0.1 | 1.9 ± 0.5 | 0.104 |
| CD152+ (CTLA-4+) | 5.7 ± 0.5 b | 5.4 ± 0.4 ab | 7.9 ± 0.6 a | 0.005 |
| % of total lymphocytes | ||||
| Total CD45RA+ (B cells) | 26.8 ± 1.3 | 36.1 ± 1.3 | 33.7 ± 0.1 | 0.554 |
| IgA+ | 4.3 ± 0.6 a | 1.6 ± 0.1 b | 2.3 ± 0.3b | 0.001 |
| IgG+ | 6.0 ± 0.9 a | 3.4 ± 0.2 b | 5.3 ± 0.5 ab | 0.024 |
| IgM+ | 38.7 ± 1.9 | 32.1 ± 2.9 | 37.4 ± 1.6 | 0.335 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lewis, E.D.; Richard, C.; Goruk, S.; Wadge, E.; Curtis, J.M.; Jacobs, R.L.; Field, C.J. Feeding a Mixture of Choline Forms during Lactation Improves Offspring Growth and Maternal Lymphocyte Response to Ex Vivo Immune Challenges. Nutrients 2017, 9, 713. https://doi.org/10.3390/nu9070713
Lewis ED, Richard C, Goruk S, Wadge E, Curtis JM, Jacobs RL, Field CJ. Feeding a Mixture of Choline Forms during Lactation Improves Offspring Growth and Maternal Lymphocyte Response to Ex Vivo Immune Challenges. Nutrients. 2017; 9(7):713. https://doi.org/10.3390/nu9070713
Chicago/Turabian StyleLewis, Erin D., Caroline Richard, Susan Goruk, Emily Wadge, Jonathan M. Curtis, René L. Jacobs, and Catherine J. Field. 2017. "Feeding a Mixture of Choline Forms during Lactation Improves Offspring Growth and Maternal Lymphocyte Response to Ex Vivo Immune Challenges" Nutrients 9, no. 7: 713. https://doi.org/10.3390/nu9070713