Platycodon grandiflorus Root Extract Improves Learning and Memory by Enhancing Synaptogenesis in Mice Hippocampus

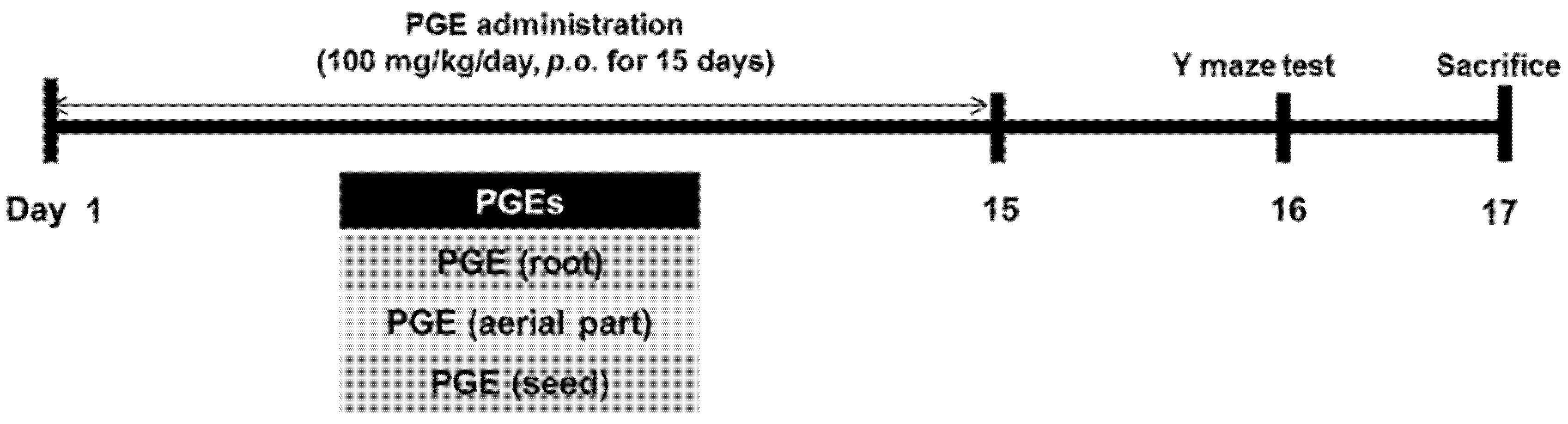{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Plant Material
2.3. Extraction from Root, Aerial Parts, and Seeds of PG
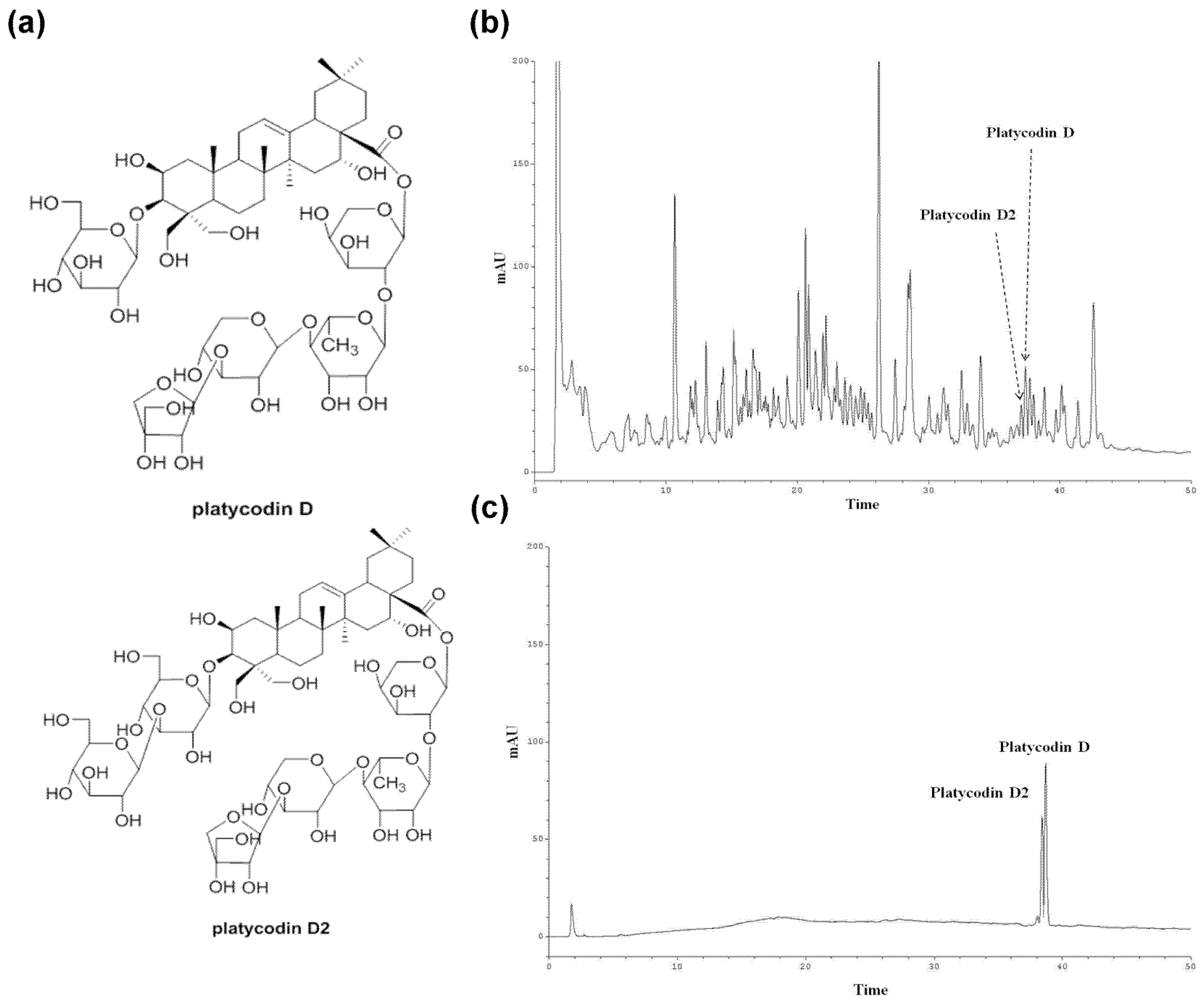
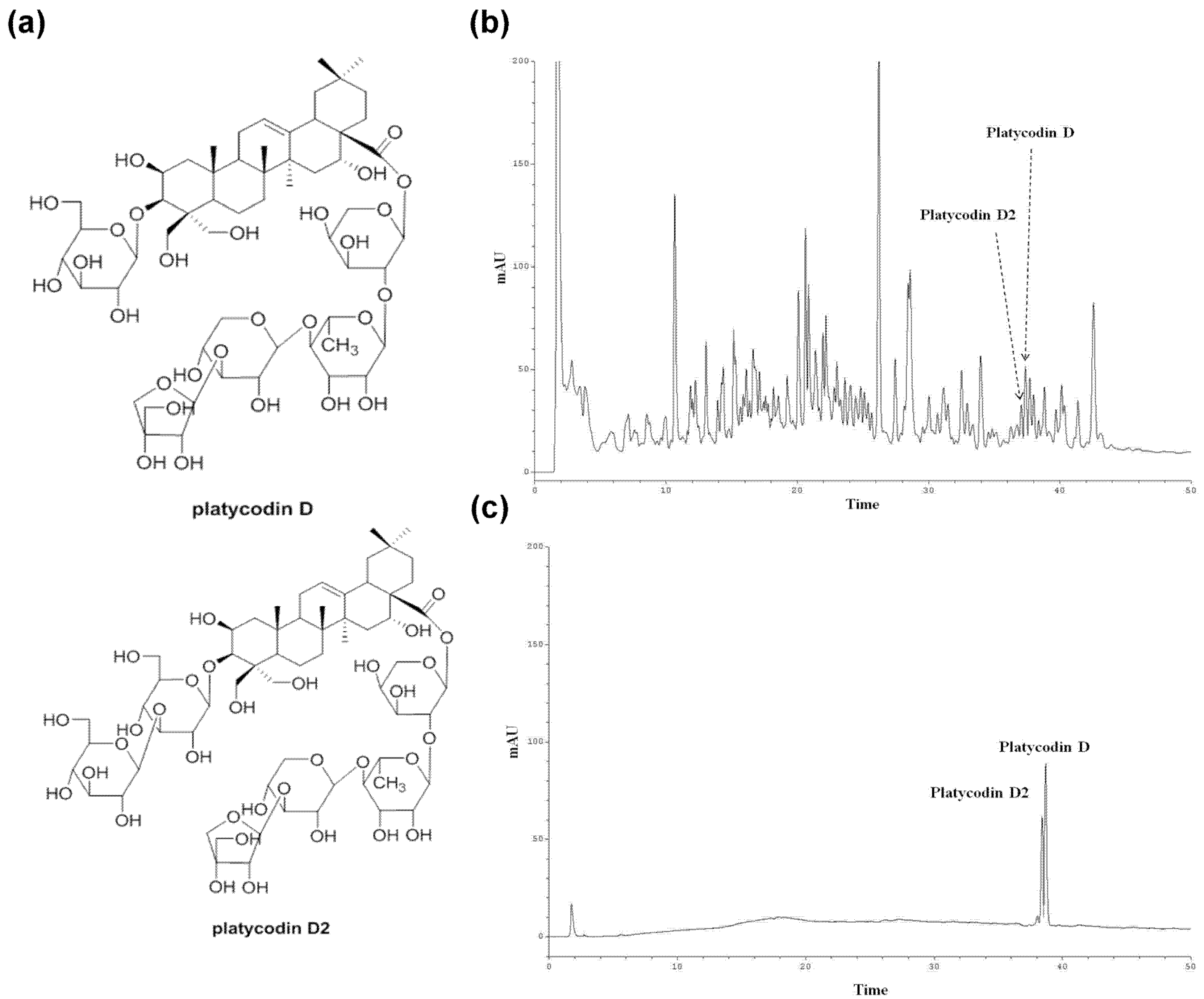
2.4. Isolation of Platycodin D and Platycodin D2
2.5. HPLC Analysis of the PGE (Root)
2.6. Animals and PGE Administration
2.7. Brain Tissue Preparation
2.8. Immunohistochemistry Analysis and Quantification
2.9. Western Blot Analysis
2.10. Y-Maze Test
2.11. Cell Viability Assay
2.12. PC12 Cell Culture and Treatment of PGE, Platycodin D, and Platycodin D2
2.13. Quantification of Neurite Outgrowth of PC12 Cells
2.14. Statistical Analysis
3. Results
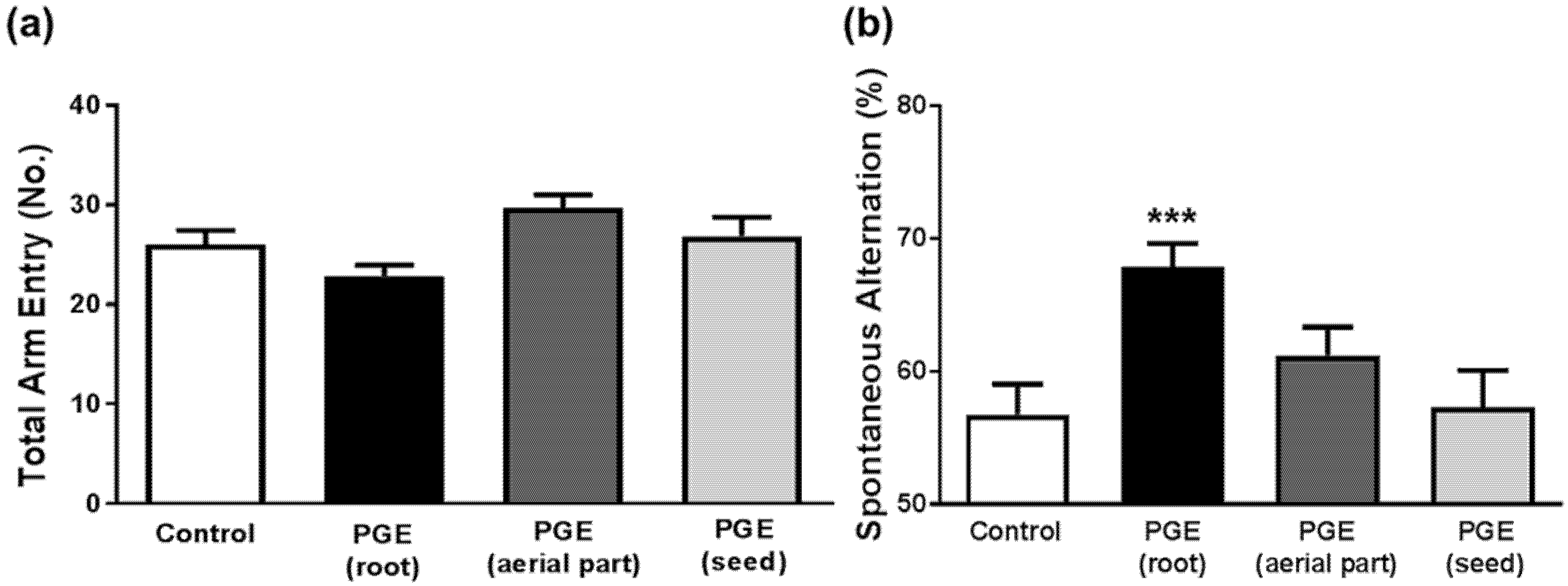
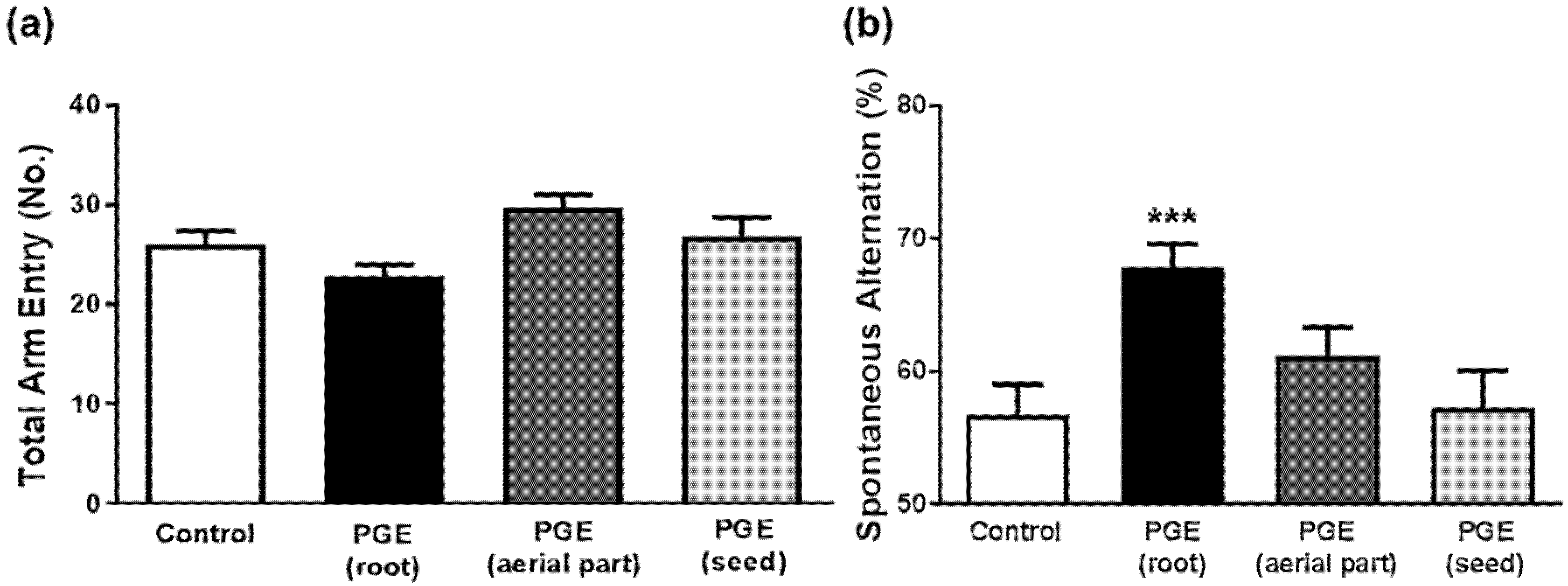
3.1. Behavioral Tests in Mice to Determine Cognitive Function after Administration of PGEs
3.2. Standardization of Platycodin D and Platycodin D2 in PGE (Root) Using HPLC
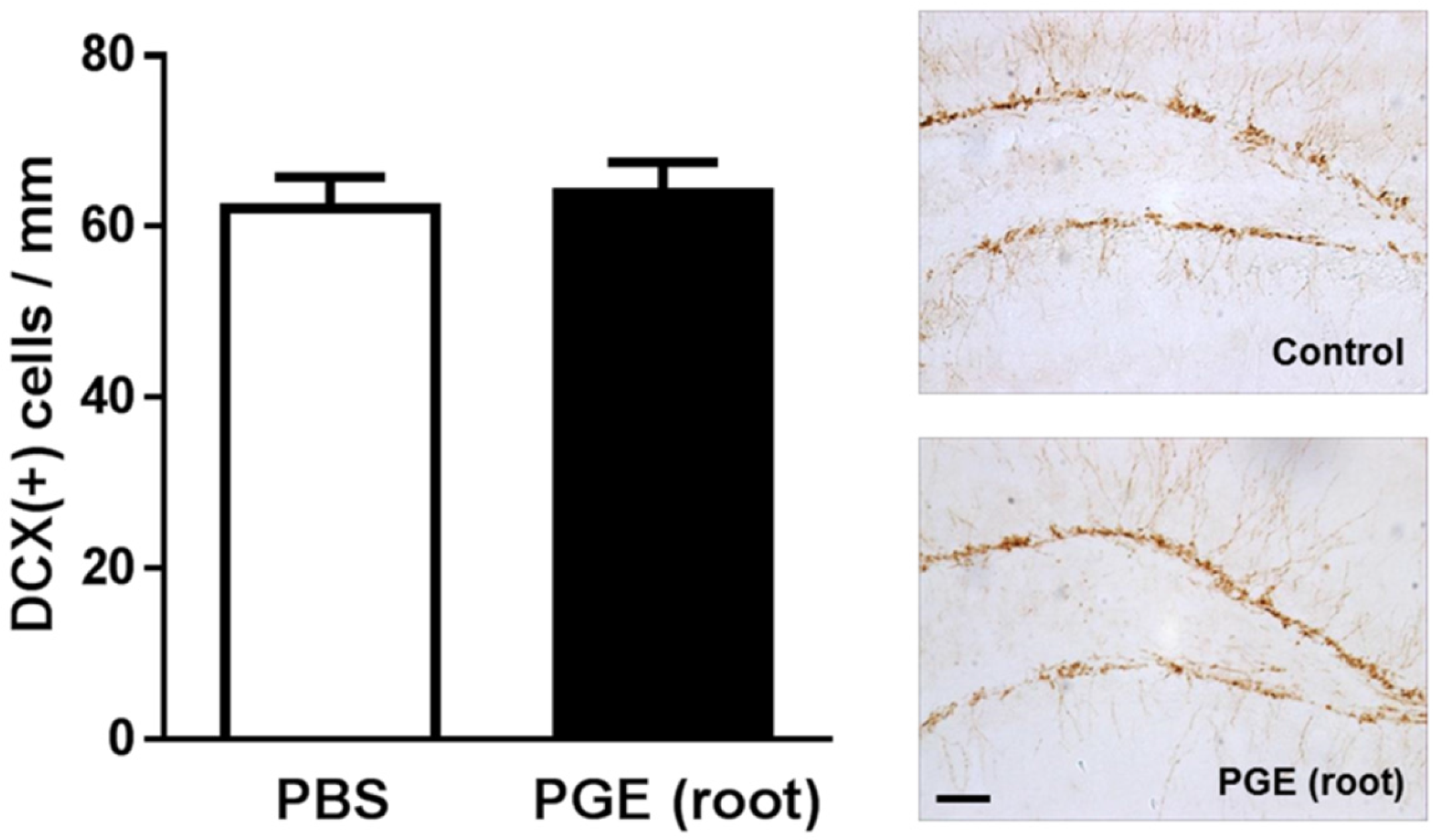
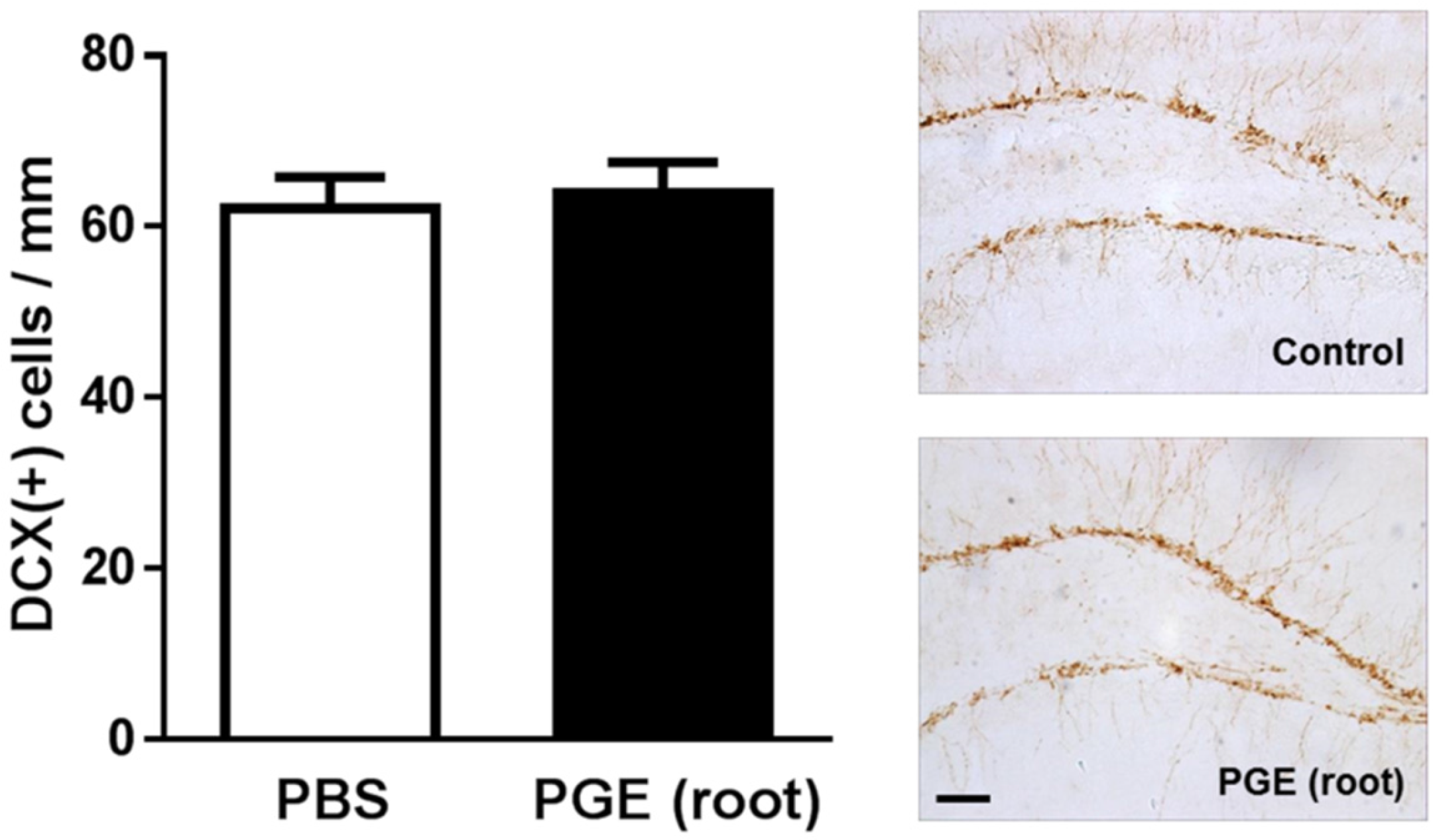
3.3. Immunohistochemical Analysis of the effect of PGE (root) on Hippocampal Neurogenesis
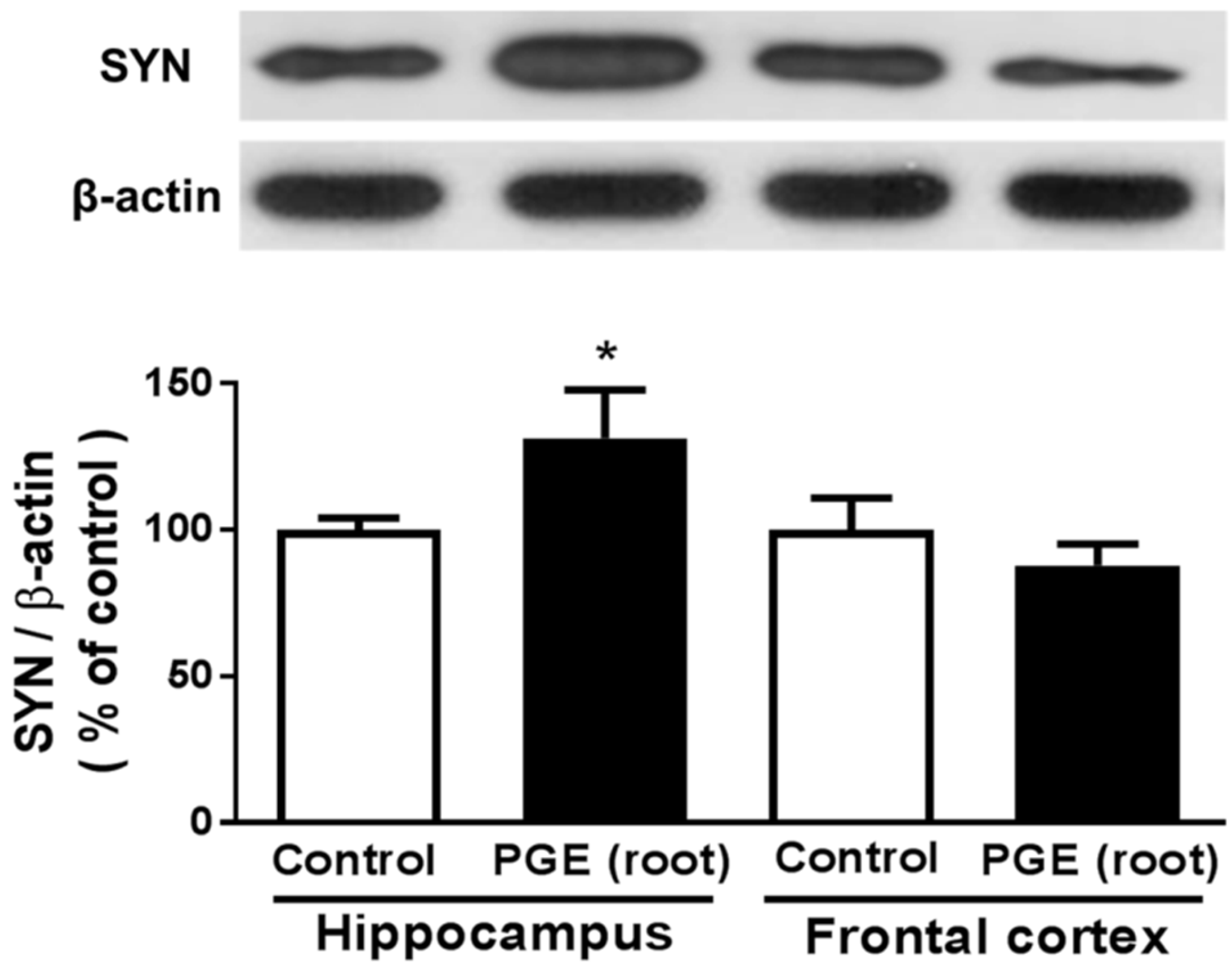
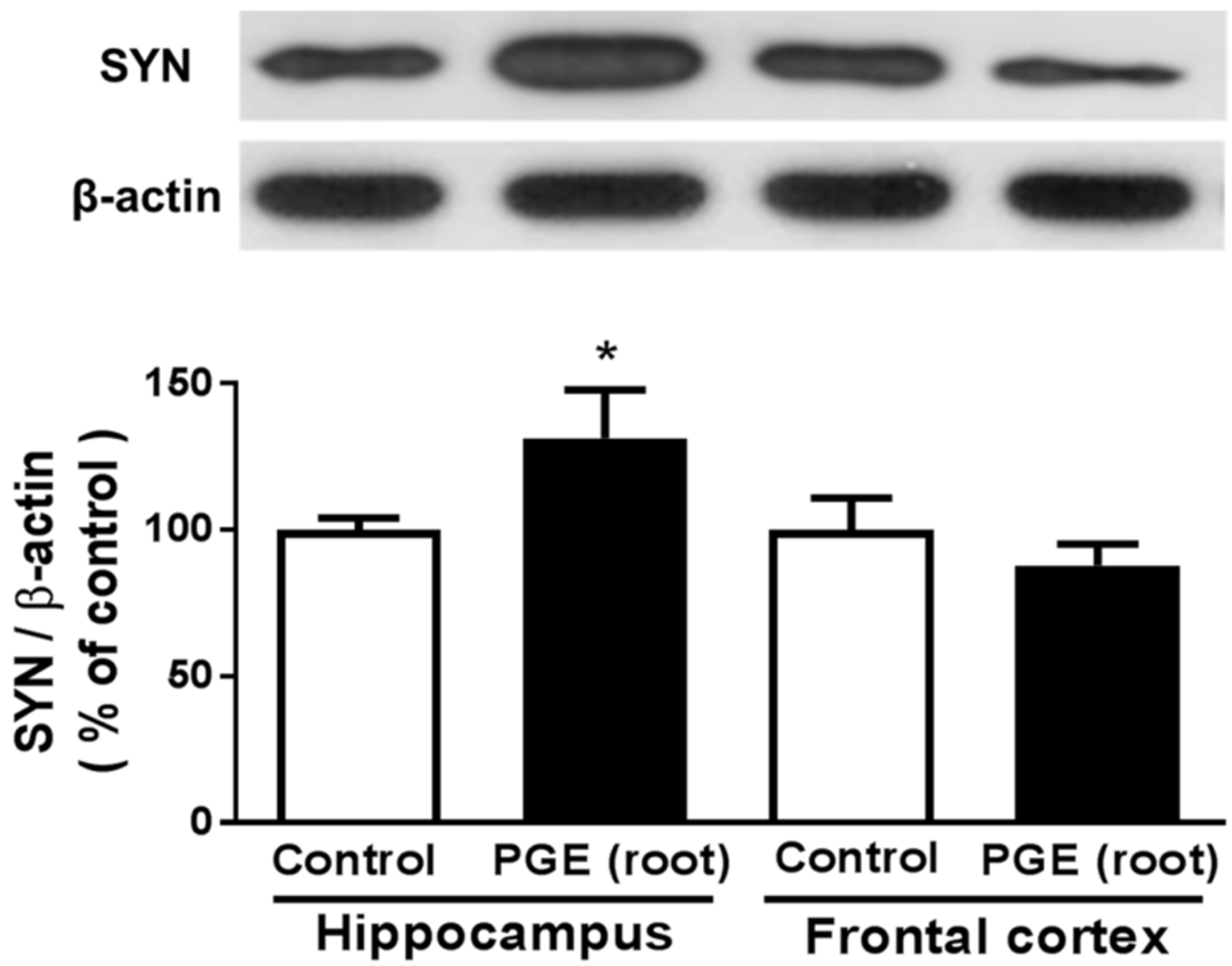
3.4. Immunoblotting Analysis of the Effect of PGE (root) on Synaptic Sprouting
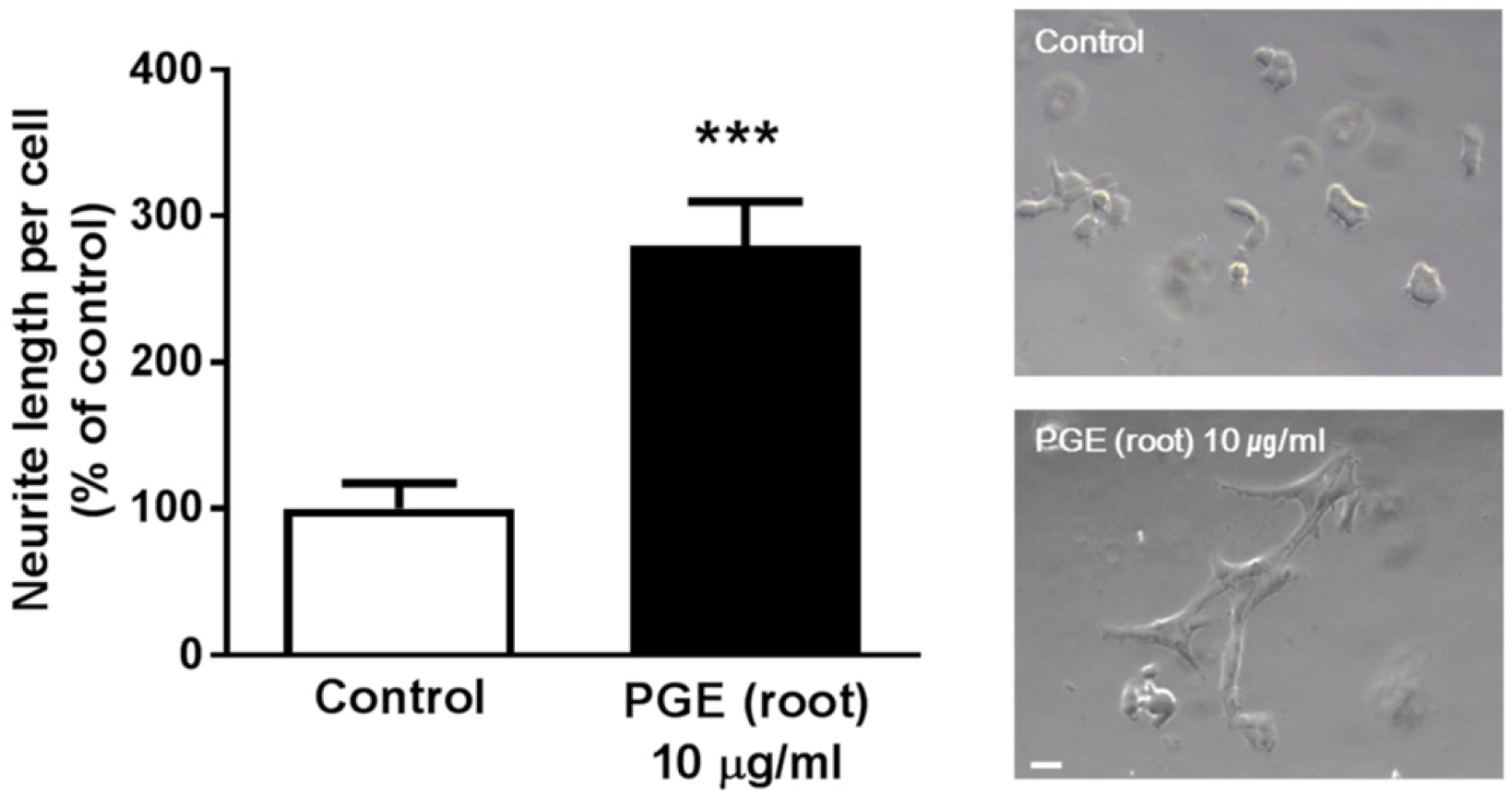
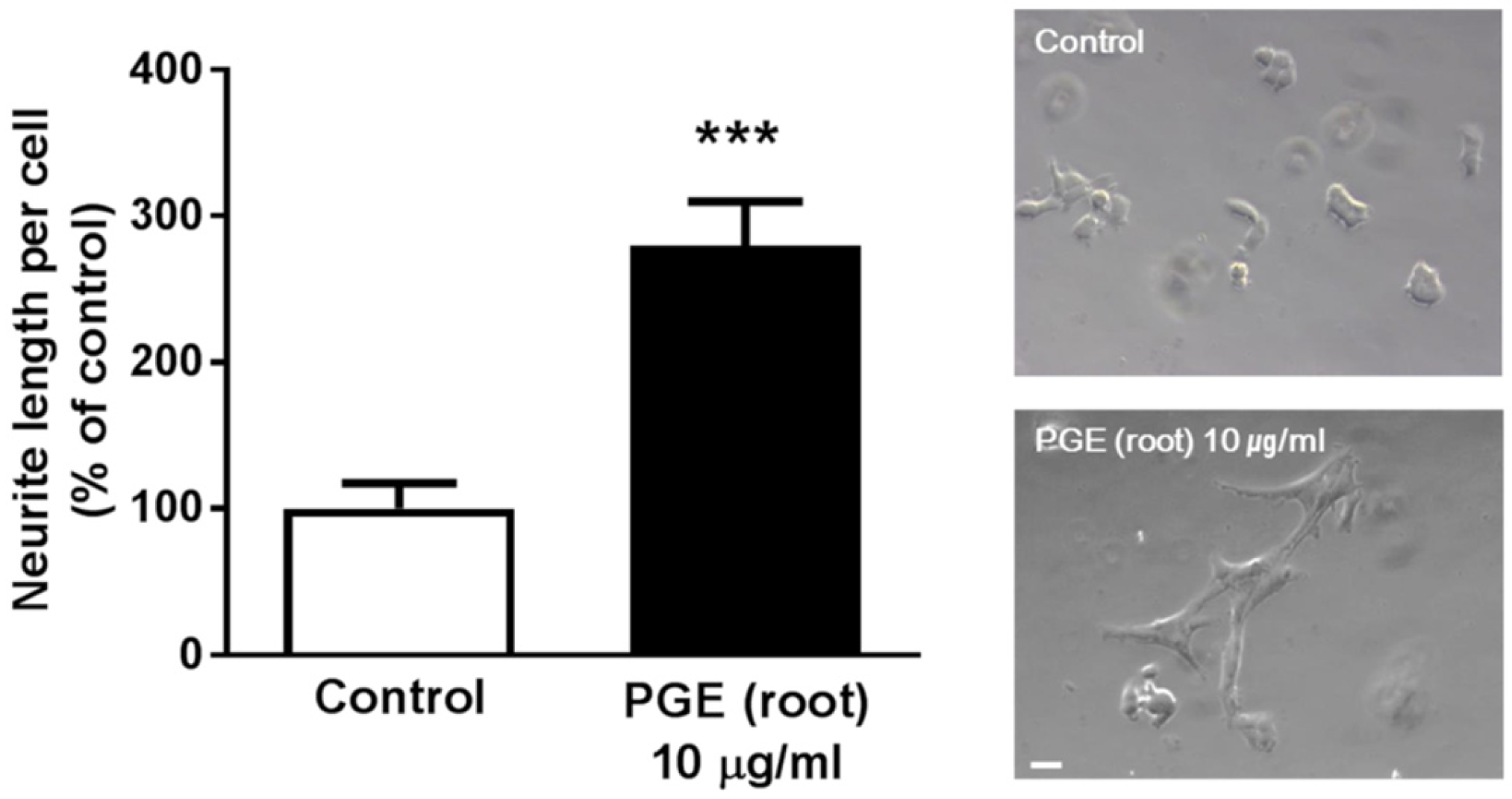
3.5. PGE (Root) Increases Neuritogenesis In Vitro
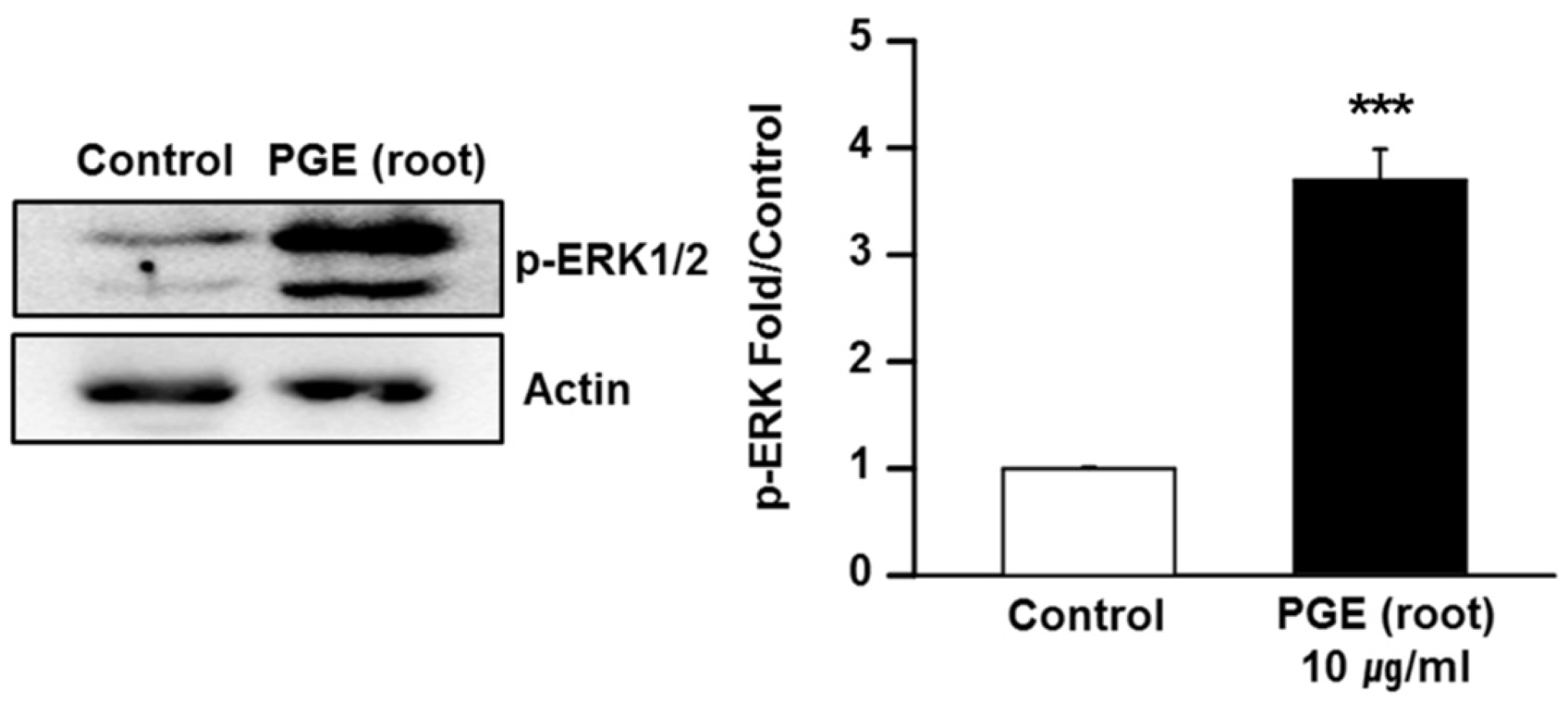
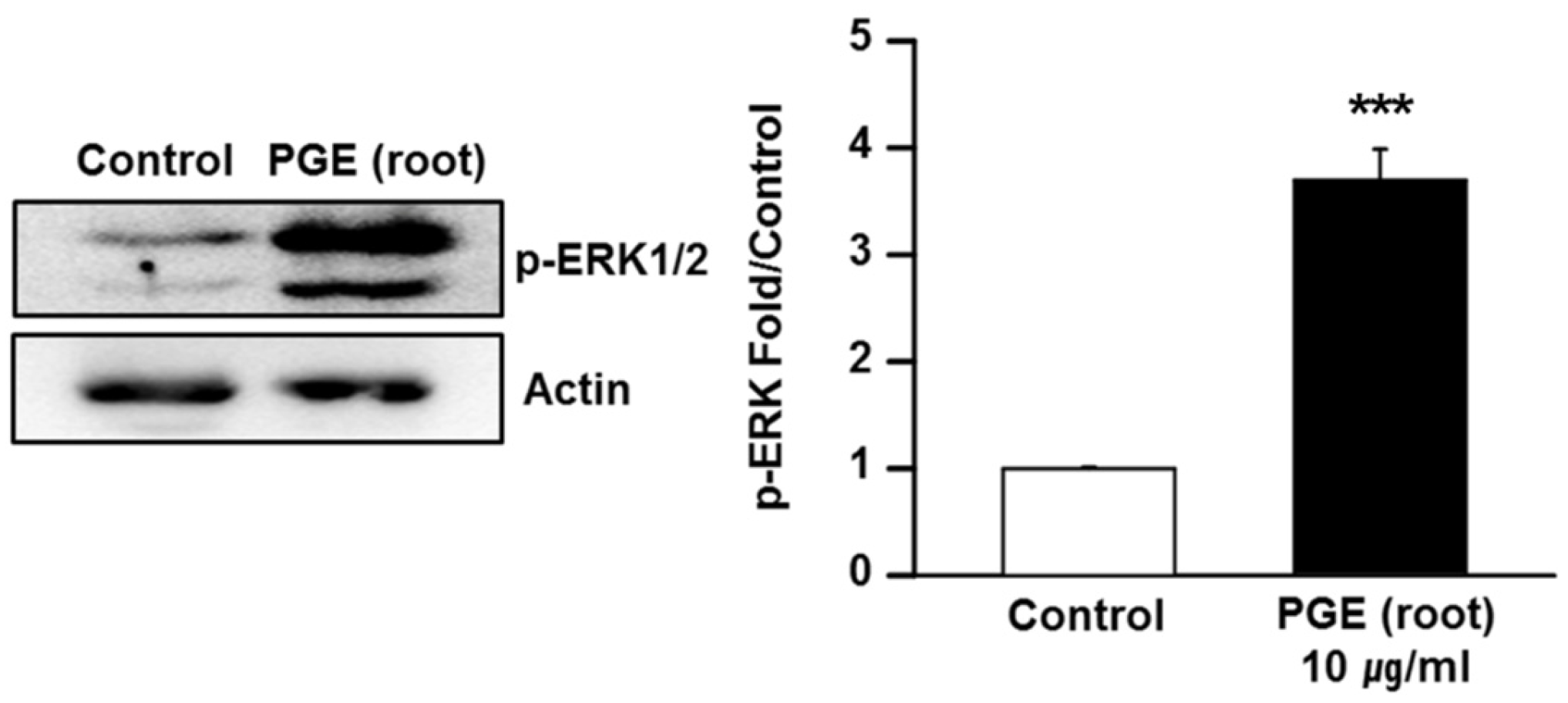
3.6. PGE (Root) Activates the MAPK/ERK Signaling Pathway
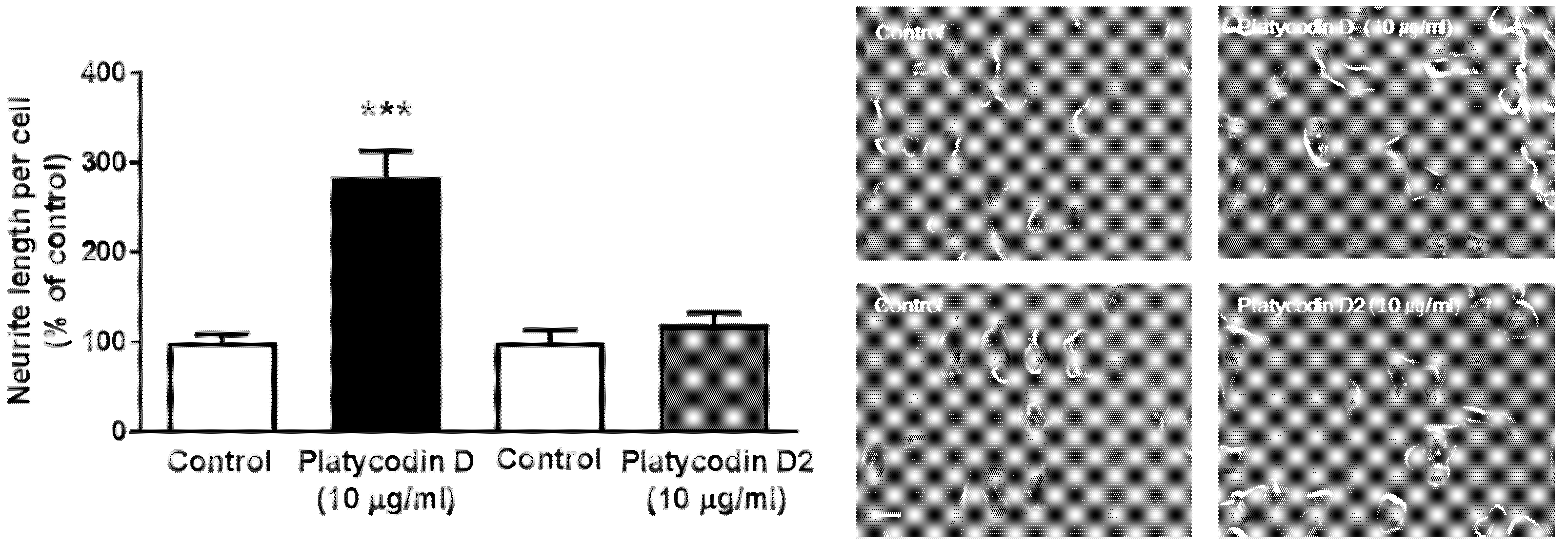
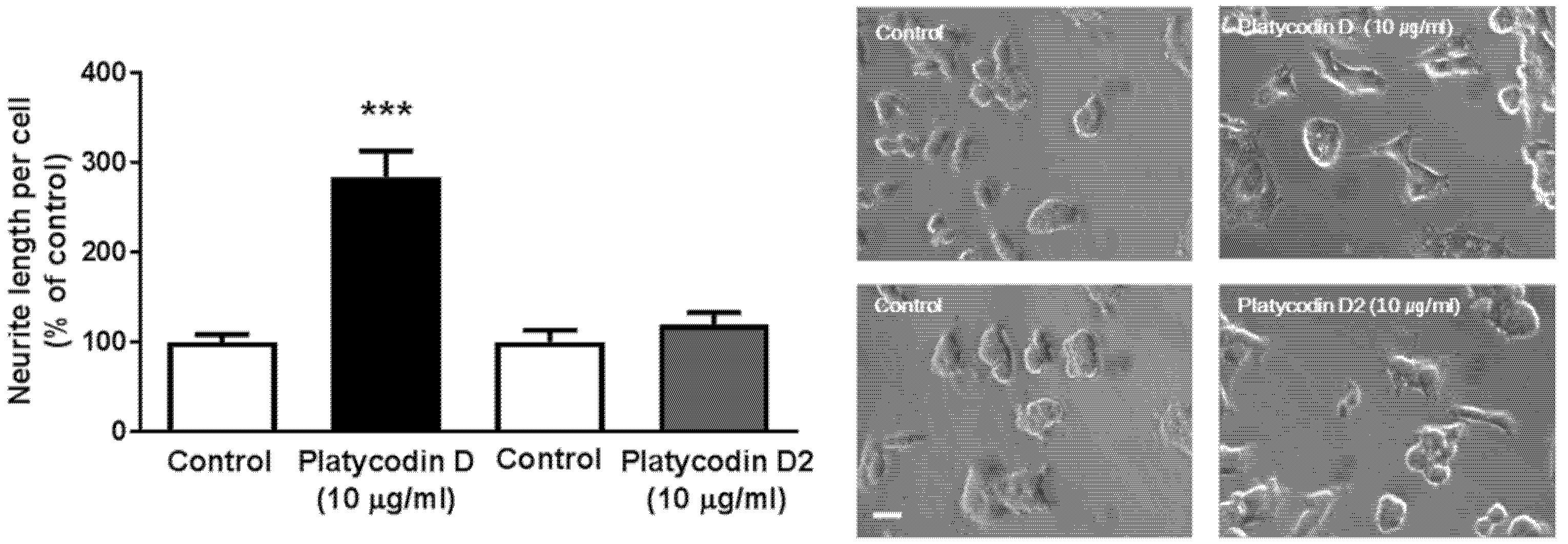
3.7. Platycodin D, a Bioactive Ingredient of PG, Increases Neuritogenesis In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kondo, M.; Takei, Y.; Hirokawa, N. Motor protein KIF1A is essential for hippocampal synaptogenesis and learning enhancement in an enriched environment. Neuron 2012, 73, 743–757. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.K.; Hongpaisan, J.; Lim, C.S.; Alkon, D.L. Bryostatin-1 restores hippocampal synapses and spatial learning and memory in adult fragile x mice. J. Pharmacol. Exp. Ther. 2014, 349, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Diorio, J.; Day, J.C.; Francis, D.D.; Meaney, M.J. Maternal care, hippocampal synaptogenesis and cognitive development in rats. Nat. Neurosci. 2000, 3, 799–806. [Google Scholar] [PubMed]
- Ramirez-Amaya, V.; Balderas, I.; Sandoval, J.; Escobar, M.L.; Bermudez-Rattoni, F. Spatial long-term memory is related to mossy fiber synaptogenesis. J. Neurosci. 2001, 21, 7340–7348. [Google Scholar] [PubMed]
- Nordeen, K.W.; Nordeen, E.J. Anatomical and synaptic substrates for avian song learning. J. Neurobiol. 1997, 33, 532–548. [Google Scholar] [CrossRef]
- Dietrich, M.O.; Andrews, Z.B.; Horvath, T.L. Exercise-induced synaptogenesis in the hippocampus is dependent on UCP2-regulated mitochondrial adaptation. J. Neurosci. 2008, 28, 10766–10771. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Akaishi, T.; Matsuki, N.; Ohno, Y.; Nakazawa, K. Beta-estradiol induces synaptogenesis in the hippocampus by enhancing brain-derived neurotrophic factor release from dentate gyrus granule cells. Brain Res. 2007, 30, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Fester, L.; Prange-Kiel, J.; Zhou, L.; Blittersdorf, B.V.; Bohm, J.; Jarry, H.; Schumacher, M.; Rune, G.M. Estrogen-regulated synaptogenesis in the hippocampus: Sexual dimorphism in vivo but not in vitro. J. Steroid Biochem. Mol. Biol. 2012, 131, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Shimada, S. Serotonin and exercise-induced brain plasticity. Neurotransmitter 2015, 2. [Google Scholar] [CrossRef]
- Yang, S.H.; Jeng, C.J.; Chen, C.H.; Chen, Y.; Chen, Y.C.; Wang, S.M. Schisandrin enhances dendrite outgrowth and synaptogenesis in primary cultured hippocampal neurons. J. Sci. Food Agric. 2011, 91, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Tchantchou, F.; Lacor, P.N.; Cao, Z.; Lao, L.; Hou, Y.; Cui, C.; Klein, W.L.; Luo, Y. Stimulation of neurogenesis and synaptogenesis by bilobalide and quercetin via common final pathway in hippocampal neurons. J. Alzheimers Dis. 2009, 18, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.; Perfilieva, E.; Johansson, U.; Orwar, O.; Eriksson, P.S. Enriched environment increases neurogenesis in the adult rat dentate gyrus and improves spatial memory. J. Neurobiol. 1999, 39, 569–578. [Google Scholar] [CrossRef]
- Galea, L.A.; Wainwright, S.R.; Roes, M.M.; Duarte-Guterman, P.; Chow, C.; Hamson, D.K. Sex, hormones and neurogenesis in the hippocampus: Hormonal modulation of neurogenesis and potential functional implications. J. Neuroendocrinol. 2013, 25, 1039–1061. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Chung, H.; Kim, Y.; Kim, D.H.; Ryu, J.H.; Sato, T.; Kojima, M.; Park, S. Ghrelin directly stimulates adult hippocampal neurogenesis: Implications for learning and memory. Endocr. J. 2013, 60, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.I.; Jeon, S.G.; Kim, K.A.; Kim, Y.J.; Song, E.J.; Choi, J.; Ahn, K.J.; Kim, C.J.; Chung, H.Y.; Moon, M.; et al. The pharmacological stimulation of nurr1 improves cognitive functions via enhancement of adult hippocampal neurogenesis. Stem Cell Res. 2016, 17, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Nokia, M.S.; Lensu, S.; Ahtiainen, J.P.; Johansson, P.P.; Koch, L.G.; Britton, S.L.; Kainulainen, H. Physical exercise increases adult hippocampal neurogenesis in male rats provided it is aerobic and sustained. J. Physiol. 2016, 594, 1855–1873. [Google Scholar] [CrossRef] [PubMed]
- Yau, S.-Y.; Gil-Mohapel, J.; Christie, B.R.; So, K.-F. Physical exercise-induced adult neurogenesis: A good strategy to prevent cognitive decline in neurodegenerative diseases? BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, B.M.; Moreira, F.A.; Massensini, A.R.; Moraes, M.F.; Pereira, G.S. Enriched environment increases neurogenesis and improves social memory persistence in socially isolated adult mice. Hippocampus 2014, 24, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Yoo, K.Y.; Park, O.K.; Hwang, I.K.; Li, H.; Ryu, S.Y.; Kang, I.J.; Yi, J.S.; Bae, Y.S.; Park, J.; Kim, Y.S.; et al. Induction of cell proliferation and neuroblasts in the subgranular zone of the dentate gyrus by aqueous extract from platycodon grandiflorum in middle-aged mice. Neurosci. Lett. 2008, 444, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Giachello, C.N.; Fiumara, F.; Giacomini, C.; Corradi, A.; Milanese, C.; Ghirardi, M.; Benfenati, F.; Montarolo, P.G. Mapk/erk-dependent phosphorylation of synapsin mediates formation of functional synapses and short-term homosynaptic plasticity. J. Cell Sci. 2010, 123, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Shioda, N.; Han, F.; Fukunaga, K. Role of AKT and erk signaling in the neurogenesis following brain ischemia. Int. Rev. Neurobiol. 2009, 85, 375–387. [Google Scholar] [PubMed]
- Selcher, J.C.; Weeber, E.J.; Christian, J.; Nekrasova, T.; Landreth, G.E.; Sweatt, J.D. A role for ERK MAP kinase in physiologic temporal integration in hippocampal area ca1. Learn. Mem. 2003, 10, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed]
- Hans, A.; Bajramovic, J.J.; Syan, S.; Perret, E.; Dunia, I.; Brahic, M.; Gonzalez-Dunia, D. Persistent, noncytolytic infection of neurons by borna disease virus interferes with ERK 1/2 signaling and abrogates bdnf-induced synaptogenesis. FASEB J. 2004, 18, 863–865. [Google Scholar] [PubMed]
- Alonso, M.; Medina, J.H.; Pozzo-Miller, L. ERK1/2 activation is necessary for BDNF to increase dendritic spine density in hippocampal ca1 pyramidal neurons. Learn. Mem. 2004, 11, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Creson, T.; Zhang, L.; Li, P.; Du, F.; Yuan, P.; Gould, T.D.; Manji, H.K.; Chen, G. Mood stabilizer valproate promotes ERK pathway-dependent cortical neuronal growth and neurogenesis. J. Neurosci. 2004, 24, 6590–6599. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Zhou, X.Y.; Hou, J.C.; Zhu, H.; Wang, Z.; Liu, J.X.; Zheng, Y.Q. Ginsenoside rd promotes neurogenesis in rat brain after transient focal cerebral ischemia via activation of PI3K/Akt pathway. Acta Pharmacol. Sin. 2015, 36, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Nyakudya, E.; Jeong, J.H.; Lee, N.K.; Jeong, Y.S. Platycosides from the roots of platycodon grandiflorum and their health benefits. Prev. Nutr. Food Sci. 2014, 19, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Kang, M.; Park, Y.-H.; Joung, H.; Yang, Y.-K.; Paik, H.Y. Perception of common korean dishes and foods among professionals in related fields. Korean J. Nutr. 2012, 45, 562–576. [Google Scholar] [CrossRef]
- Kim, M.H. Contemplation on the emergency foods in korea under the Japanese occupation. J. East Asian Soc. Diet. Life 2015, 25, 721–738. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Yang, D.; Zhang, C.; Zhang, N.; Li, M.; Liu, Y. Platycodon grandiflorus—An ethnopharmacological, phytochemical and pharmacological review. J. Ethnopharmacol. 2015, 164, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.C.; Kang, O.H.; Choi, J.G.; Lee, Y.S.; Brice, O.O.; Jung, H.J.; Hong, S.H.; Lee, Y.M.; Shin, D.W.; Kim, Y.S.; et al. Anti-allergic activity of a platycodon root ethanol extract. Int. J. Mol. Sci. 2010, 11, 2746–2758. [Google Scholar] [CrossRef] [PubMed]
- Bae, N.; Ahn, T.; Chung, S.; Oh, M.S.; Ko, H.; Oh, H.; Park, G.; Yang, H.O. The neuroprotective effect of modified yeoldahanso-tang via autophagy enhancement in models of parkinson’s disease. J. Ethnopharmacol. 2011, 134, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.J.; Kim, H.K.; Han, M.H.; Oh, Y.N.; Yoon, H.M.; Chung, Y.H.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Choi, Y.H. Anti-inflammatory effects of saponins derived from the roots of platycodon grandiflorus in lipopolysaccharidestimulated bv2 microglial cells. Int. J. Mol. Med. 2013, 31, 1357–1366. [Google Scholar] [PubMed]
- Choi, J.H.; Yoo, K.Y.; Park, O.K.; Lee, C.H.; Won, M.H.; Hwang, I.K.; Ryu, S.Y.; Kim, Y.S.; Yi, J.S.; Bae, Y.S.; et al. Platycodin D and 2”-o-acetyl-polygalacin D2 isolated from platycodon grandiflorum protect ischemia/reperfusion injury in the gerbil hippocampus. Brain Res. 2009, 1279, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Son, I.H.; Park, Y.H.; Lee, S.I.; Yang, H.D.; Moon, H.I. Neuroprotective activity of triterpenoid saponins from platycodi radix against glutamate-induced toxicity in primary cultured rat cortical cells. Molecules 2007, 12, 1147–1152. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.G.; Kim, J.H.; Shin, E.H.; Kang, Y.R.; Lee, B.G.; Park, S.H.; Moon, D.I.; Kwon, L.S.; Kim, Y.P.; Choi, M.H.; et al. Improving effects of platycodon extracts jelly on β-amyloid-induced cytotoxicity and scopolamine-induced cognitive impairment animal models. Korean J. Plant Resour. 2013, 26, 417–425. [Google Scholar] [CrossRef]
- Choi, Y.H.; Kim, Y.S.; Yeo, S.J.; Roh, S.H.; Jeong, Y.C.; Kang, J.S.; Ryu, S.Y. Ameliorating effect of balloon flower saponin on the ethanol-induced memory impairment in mice. Phytother. Res. 2008, 22, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Moon, M.K.; Ahn, J.Y.; Kim, S.; Ryu, S.Y.; Kim, Y.S.; Ha, T.Y. Ethanol extract and saponin of platycodon grandiflorum ameliorate scopolamine-induced amnesia in mice. J. Med. Food 2010, 13, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Cha, M.R.; Choi, C.W.; Yoo, D.S.; Choi, Y.H.; Park, B.K.; Kim, E.J.; Kim, Y.S.; Kang, J.S.; Kim, Y.H.; Ryu, S.Y. Cognitive enhancing effect of saponin rich fraction from the roots extract of platycodon grandiflorum in mice. Korean J. Pharmacogn. 2011, 42, 5. [Google Scholar]
- Ha, Y.W.; Na, Y.C.; Seo, J.J.; Kim, S.N.; Linhardt, R.J.; Kim, Y.S. Qualitative and quantitative determination of ten major saponins in platycodi radix by high performance liquid chromatography with evaporative light scattering detection and mass spectrometry. J. Chromatogr. A 2006, 24, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Hien, T.T.; Han, E.H.; Chung, Y.C.; Jeong, H.G. Molecular mechanism of endothelial nitric-oxide synthase activation by platycodon grandiflorum root-derived saponins. Toxicol. Lett. 2010, 195, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Ji, S.H.; Kim, G.S.; Song, K.S.; Um, Y.; Kim, O.T.; Lee, Y.; Hong, C.P.; Shin, D.H.; Kim, C.K.; et al. Global profiling of various metabolites in platycodon grandiflorum by UPLC-QTOF/MS. Int. J. Mol. Sci. 2015, 16, 26786–26796. [Google Scholar] [CrossRef] [PubMed]
- Jeong, C.H.; Shim, K.H. Chemical composition and antioxidative activities of platycodon grandiflorum leaves and stems. J. Korean Soc. Food Sci. Nutr. 2006, 35, 511–515. [Google Scholar]
- Xing, Q.; Fu, Q.; Jin, Y.; Liang, X. preparation of triterpene saponins from platycodon grandiflorum by two-dimensional hydrophilic interaction liquid chromatography-reversed phase liquid chromatography. Se pu. 2014, 32, 767–772. [Google Scholar] [PubMed]
- Jeong, C.-H.; Choi, G.N.; Kim, J.H.; Kwak, J.H.; Kim, D.O.; Kim, Y.J.; Heo, H.J. Antioxidant activities from the aerial parts of platycodon grandiflorum. Food Chem. 2010, 118, 278–282. [Google Scholar] [CrossRef]
- Moon, M.; Song, H.; Hong, H.J.; Nam, D.W.; Cha, M.Y.; Oh, M.S.; Yu, J.; Ryu, H.; Mook-Jung, I. Vitamin D-binding protein interacts with abeta and suppresses abeta-mediated pathology. Cell Death Differ. 2013, 20, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Tori, K.; Tozyo, T.; Yoshimura, Y. Saponins from roots of platycodon grandiflorum. Part 2. Isolation and structure of new triterpene glycosides. J. Chem. Soc. Perkin Trans. 1 1984, 661–668. [Google Scholar] [CrossRef]
- Masliah, E.; Fagan, A.M.; Terry, R.D.; DeTeresa, R.; Mallory, M.; Gage, F.H. Reactive synaptogenesis assessed by synaptophysin immunoreactivity is associated with gap-43 in the dentate gyrus of the adult rat. Exp. Neurol. 1991, 113, 131–142. [Google Scholar] [CrossRef]
- Spellman, T.; Rigotti, M.; Ahmari, S.E.; Fusi, S.; Gogos, J.A.; Gordon, J.A. Hippocampal-prefrontal input supports spatial encoding in working memory. Nature 2015, 522, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.P.; Klumperman, J.; Lukowiak, K.; Syed, N.I. In vitro synaptogenesis between the somata of identified lymnaea neurons requires protein synthesis but not extrinsic growth factors or substrate adhesion molecules. J. Neurosci. 1997, 17, 7839–7849. [Google Scholar] [PubMed]
- Liu, Z. Preparation of botanical samples for biomedical research. Endocr. Metab. Immune Disord. Drug Targets 2008, 8, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Han, L.K.; Zheng, Y.N.; Xu, B.J.; Okuda, H.; Kimura, Y. Saponins from platycodi radix ameliorate high fat diet-induced obesity in mice. J. Nutr. 2002, 132, 2241–2245. [Google Scholar] [PubMed]
- Kim, T.Y.; Shin, Y.W. The effect of femented platycodon grandiflorum on the memory impairment of mice. Korea J. Herbol. 2013, 28, 25–31. [Google Scholar] [CrossRef]
- Ben Abdallah, N.M.; Slomianka, L.; Vyssotski, A.L.; Lipp, H.P. Early age-related changes in adult hippocampal neurogenesis in C57 mice. Neurobiol. Aging 2010, 31, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Liu, L.; Liu, J.; Dou, L.; Wang, G.; Liu, X.; Yuan, Q. Ginsenoside rg1 protects against neurodegeneration by inducing neurite outgrowth in cultured hippocampal neurons. Neural Regen. Res. 2016, 11, 319–325. [Google Scholar] [PubMed]
- Liu, J.; He, J.; Huang, L.; Dou, L.; Wu, S.; Yuan, Q. Neuroprotective effects of ginsenoside RB1 on hippocampal neuronal injury and neurite outgrowth. Neural Regen. Res. 2014, 9, 943–950. [Google Scholar] [PubMed]
- Yan, J.; Liu, Q.; Dou, Y.; Hsieh, Y.; Liu, Y.; Tao, R.; Zhu, D.; Lou, Y. Activating glucocorticoid receptor-erk signaling pathway contributes to ginsenoside RG1 protection against beta-amyloid peptide-induced human endothelial cells apoptosis. J. Ethnopharmacol. 2013, 147, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Yin, Y.; Wei, G.; Cui, J.; Zhang, E.; Guan, Y.; Yan, J.; Guo, C.; Zhu, Y.; Mu, F.; et al. Chikusetsu saponin iva confers cardioprotection via SIRT1/ERK1/2 and homer1a pathway. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.D.; Xia, F.; Lin, X.M.; Duan, K.L.; Wang, F.; Lu, Q.L.; Cao, H.; Qian, Y.H.; Shi, M. Ginsenoside-Rd promotes neurite outgrowth of pc12 cells through MAPK/ERK- and PI3K/AKT-dependent pathways. Int. J. Mol. Sci. 2016, 17, 177. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.J.; Shin, M.G.; Kim, E.S.; Lee, Y.S.; Shin, Y.J.; Jeon, S.Y. Cytoprotective effects of platycodon grandiflorus (jacq.) a.DC on C6 glioma cell apoptosis by oxidative stress. J. Physiol. Pathol. Korean Med. 2014, 28, 396–402. [Google Scholar] [CrossRef]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-i.; Jeon, S.G.; Kim, K.A.; Kim, J.-J.; Song, E.J.; Jeon, Y.; Kim, E.; Lee, K.B.; Kwak, J.H.; Moon, M. Platycodon grandiflorus Root Extract Improves Learning and Memory by Enhancing Synaptogenesis in Mice Hippocampus. Nutrients 2017, 9, 794. https://doi.org/10.3390/nu9070794
Kim J-i, Jeon SG, Kim KA, Kim J-J, Song EJ, Jeon Y, Kim E, Lee KB, Kwak JH, Moon M. Platycodon grandiflorus Root Extract Improves Learning and Memory by Enhancing Synaptogenesis in Mice Hippocampus. Nutrients. 2017; 9(7):794. https://doi.org/10.3390/nu9070794
Chicago/Turabian StyleKim, Jin-il, Seong Gak Jeon, Kyoung Ah Kim, Jwa-Jin Kim, Eun Ji Song, Yukyoung Jeon, Eunbin Kim, Kyung Bok Lee, Jong Hwan Kwak, and Minho Moon. 2017. "Platycodon grandiflorus Root Extract Improves Learning and Memory by Enhancing Synaptogenesis in Mice Hippocampus" Nutrients 9, no. 7: 794. https://doi.org/10.3390/nu9070794