Suppression of Wnt Signaling and Osteogenic Changes in Vascular Smooth Muscle Cells by Eicosapentaenoic Acid

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiment
2.2. Immunohistochemical Staining
2.3. Cell Culture
2.4. Cell Staining
2.5. Quantitative PCR
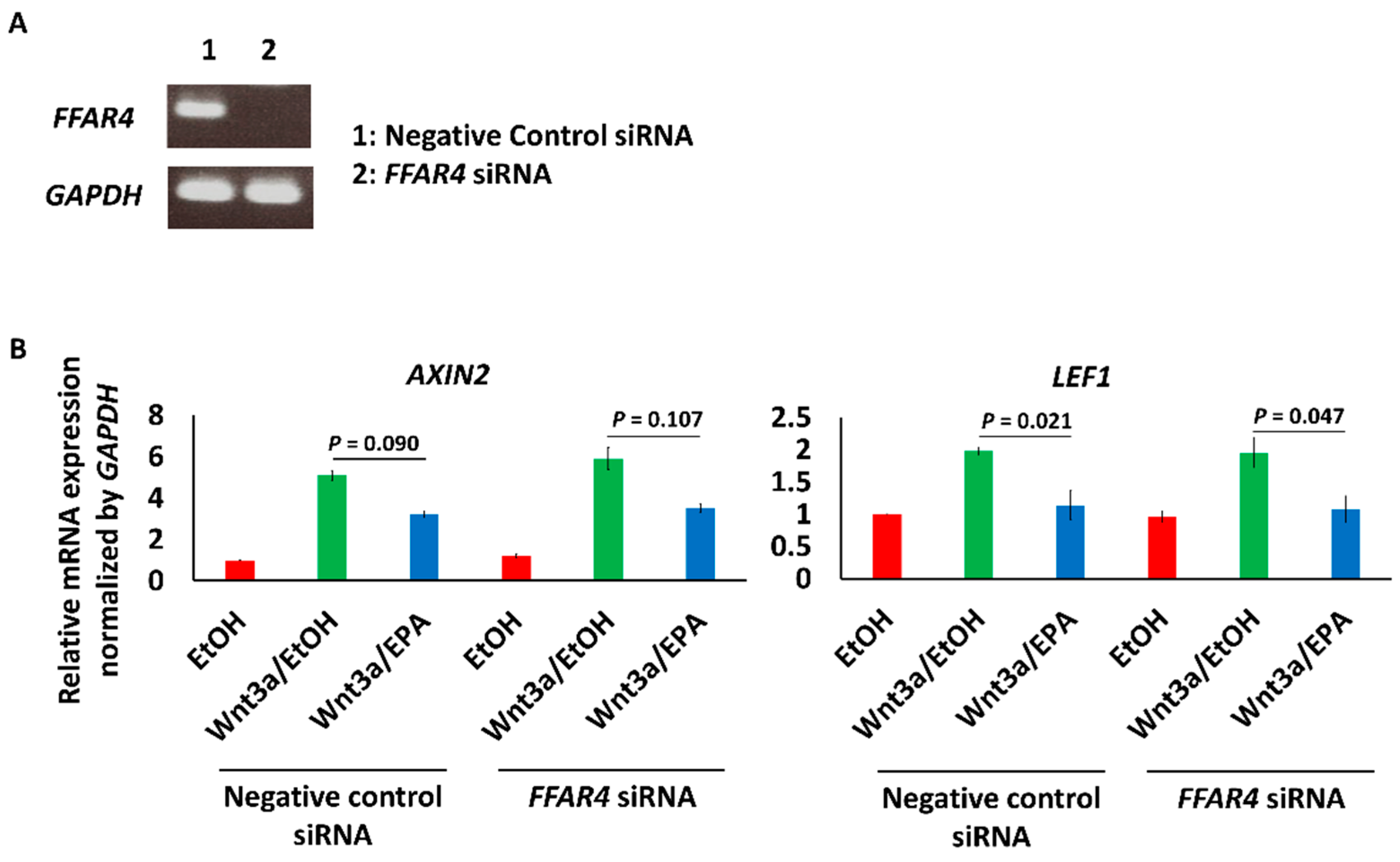
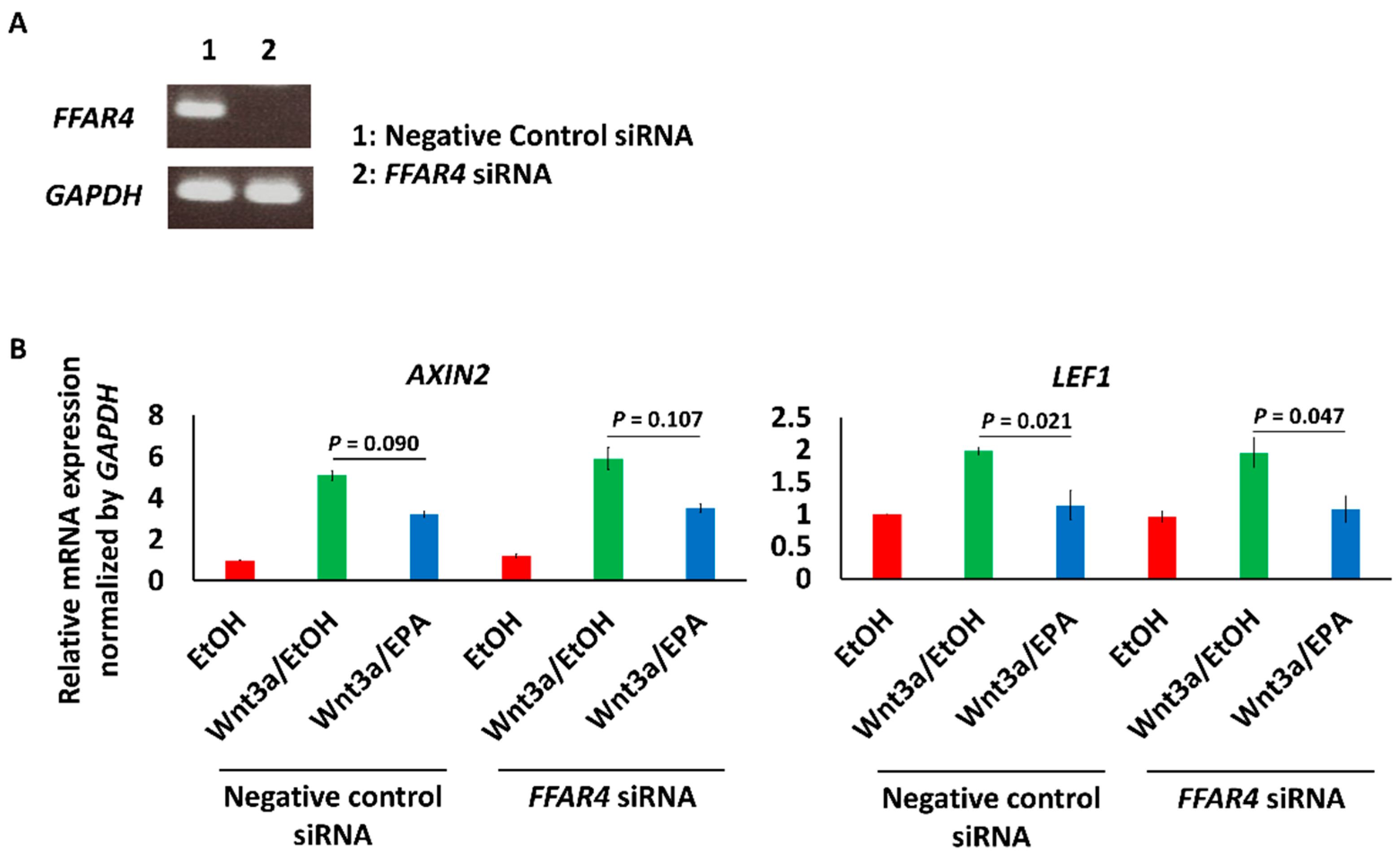
2.6. FFAR4 Knockdown by siRNA
2.7. Statistical Analysis
3. Results
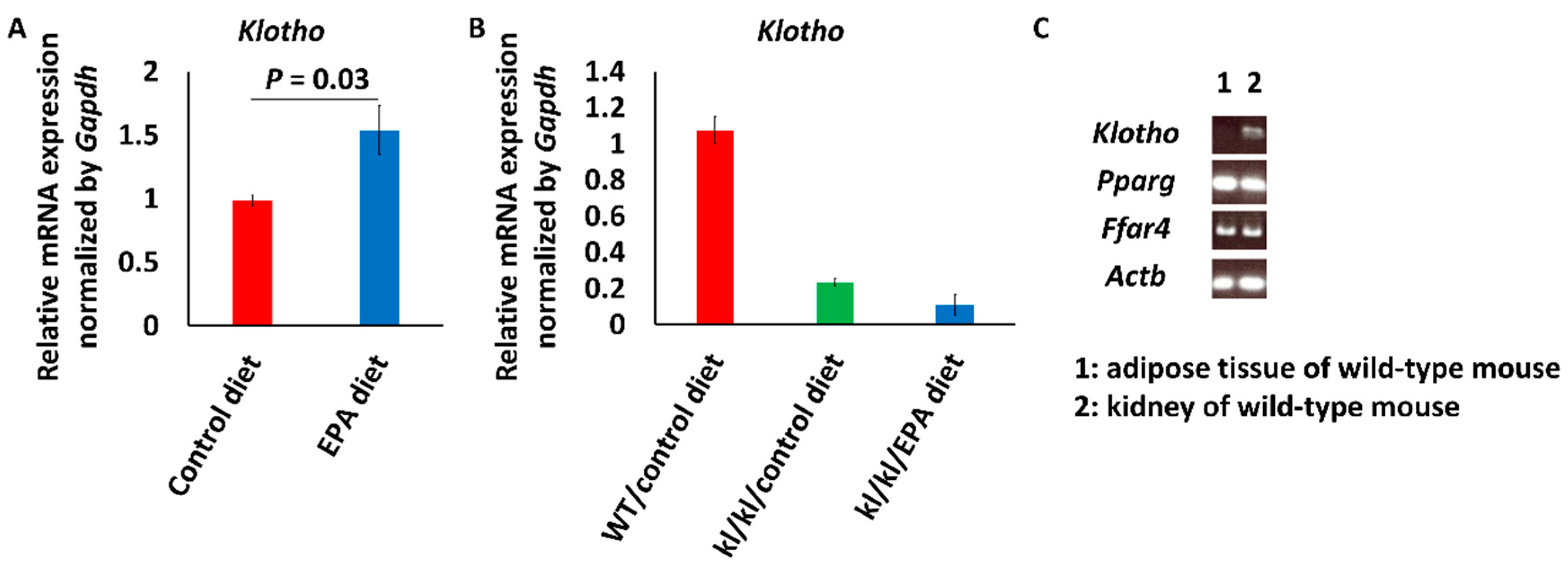
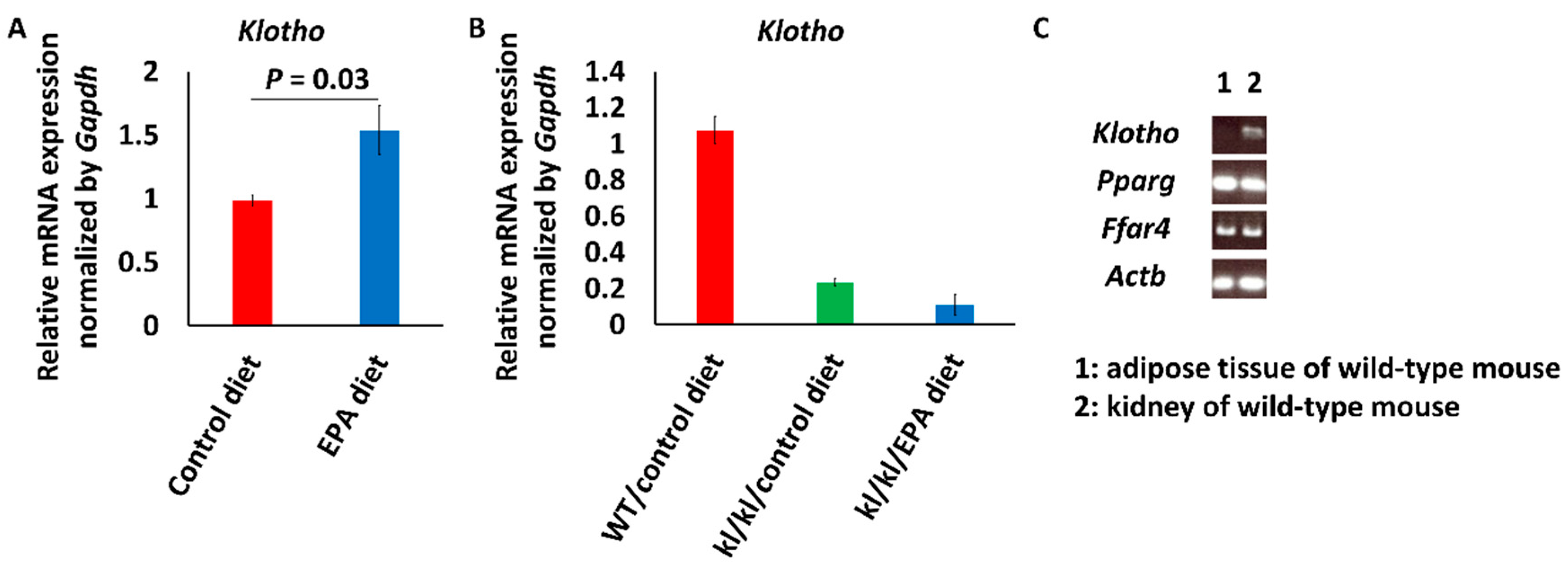
3.1. Intake of EPA Food Upregulated Klotho Expression in Kidneys of Wild-Type Mice
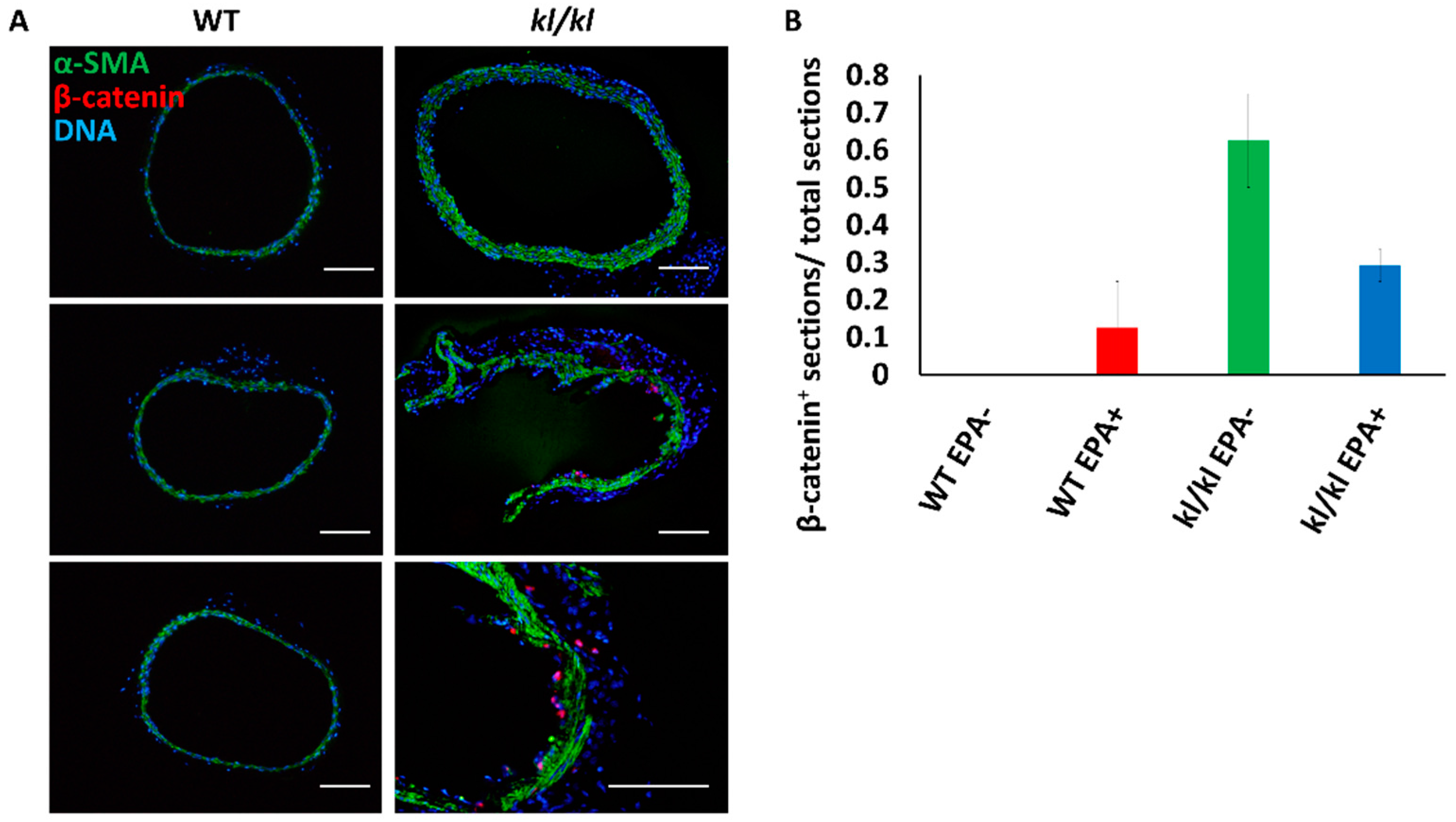
3.2. Beta-Catenin Expression Increased in the Aortas of Kl/kl Mice
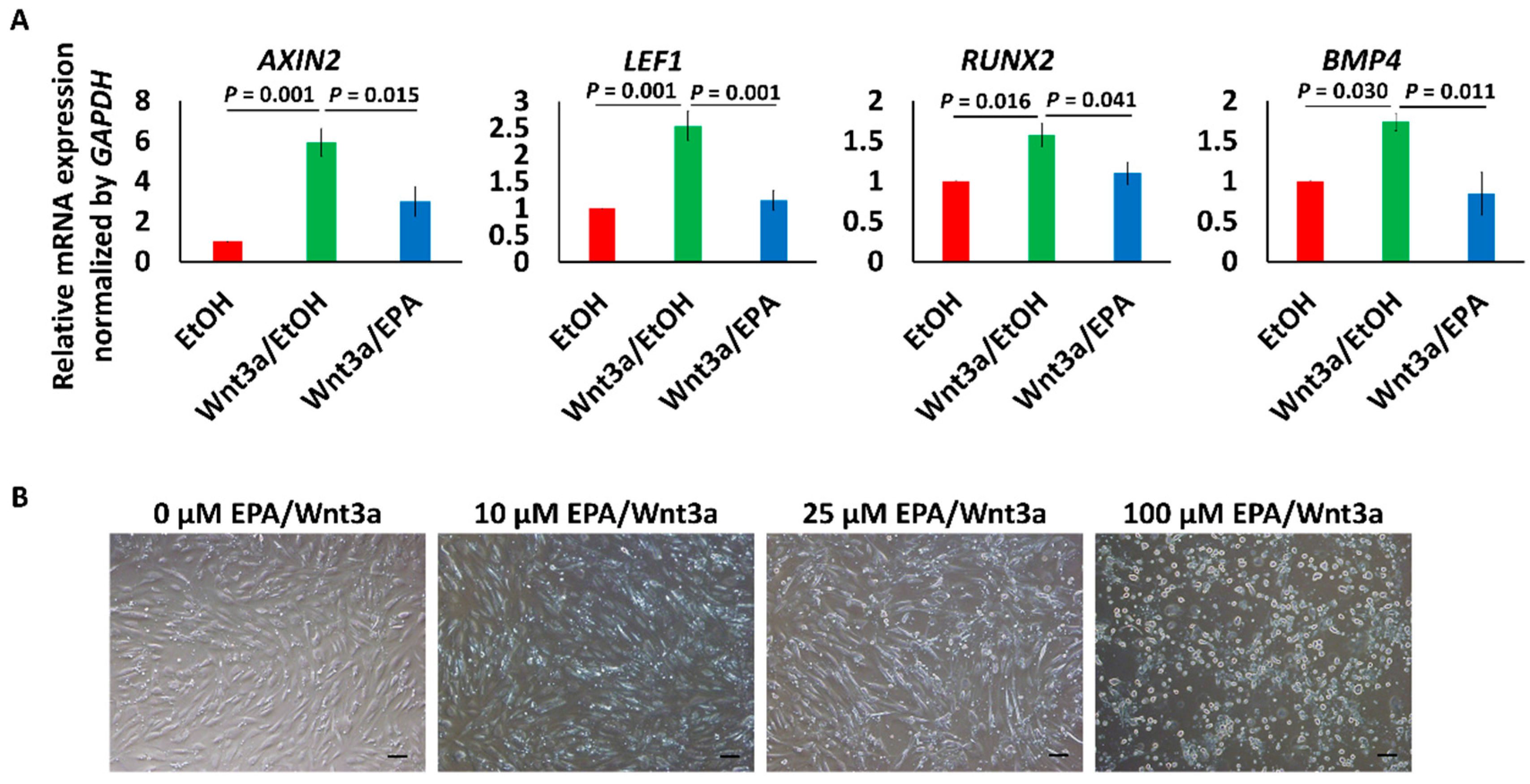
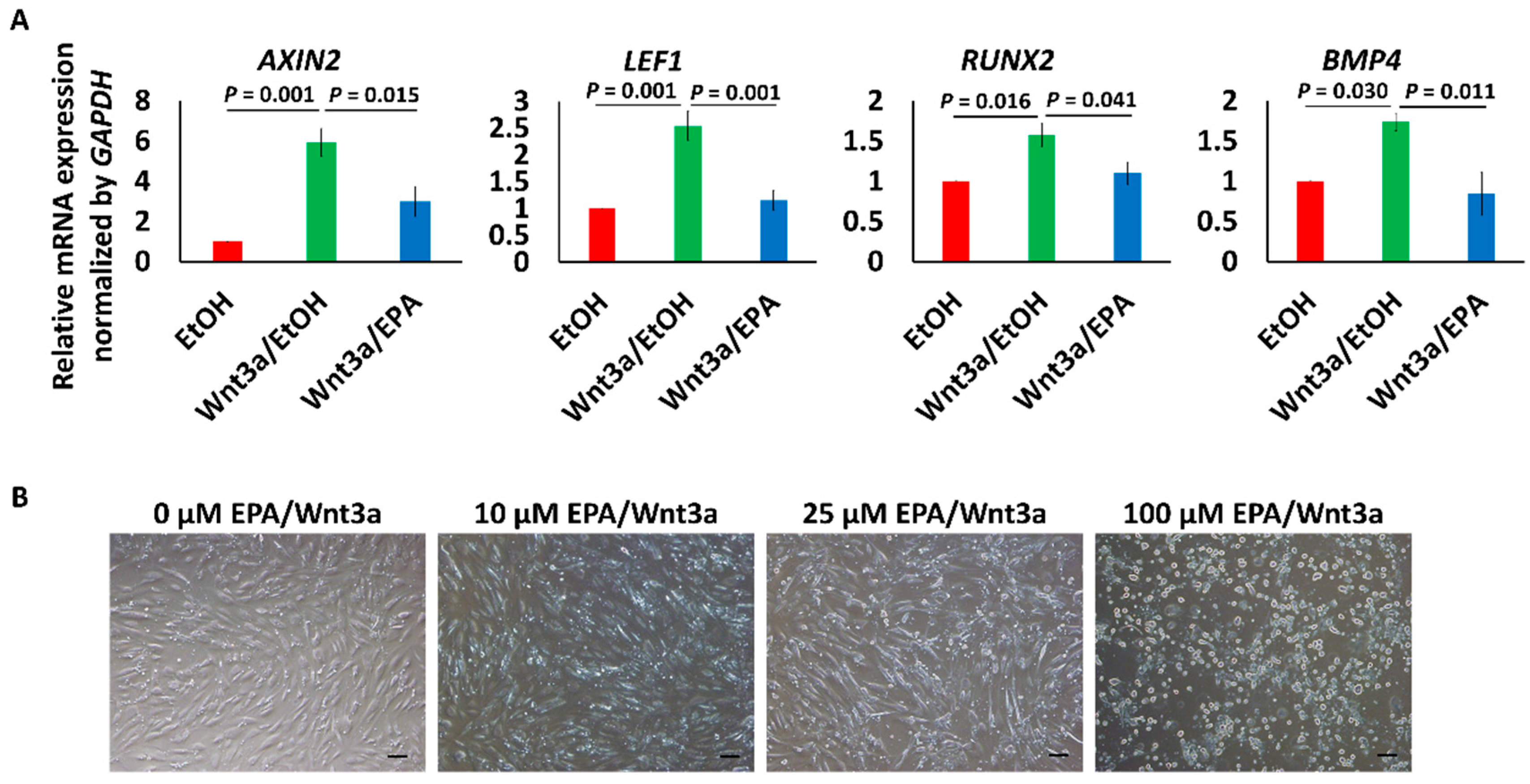
3.3. EPA Suppressed the Expression of Wnt Signaling Marker and Early Osteogenic Genes
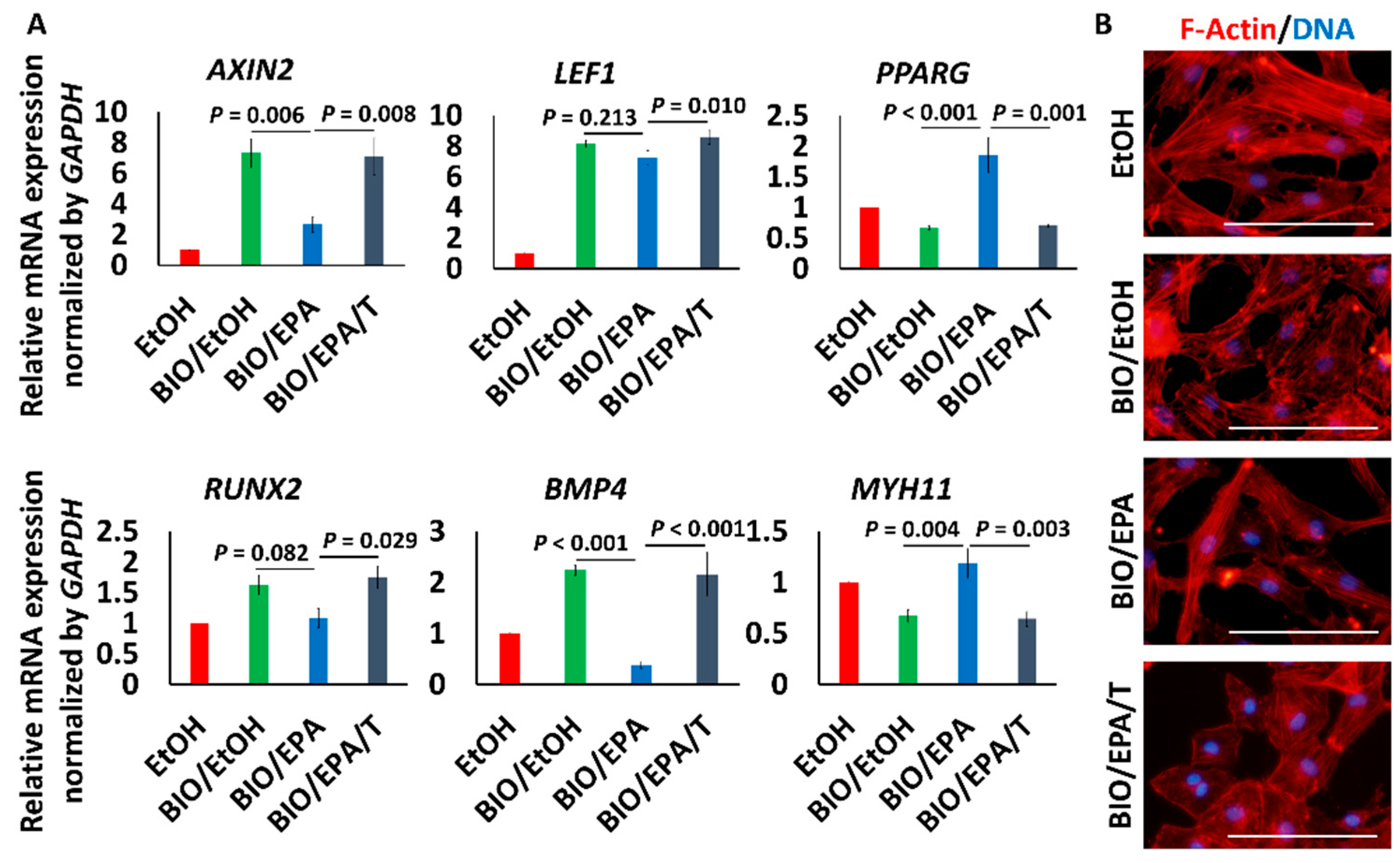
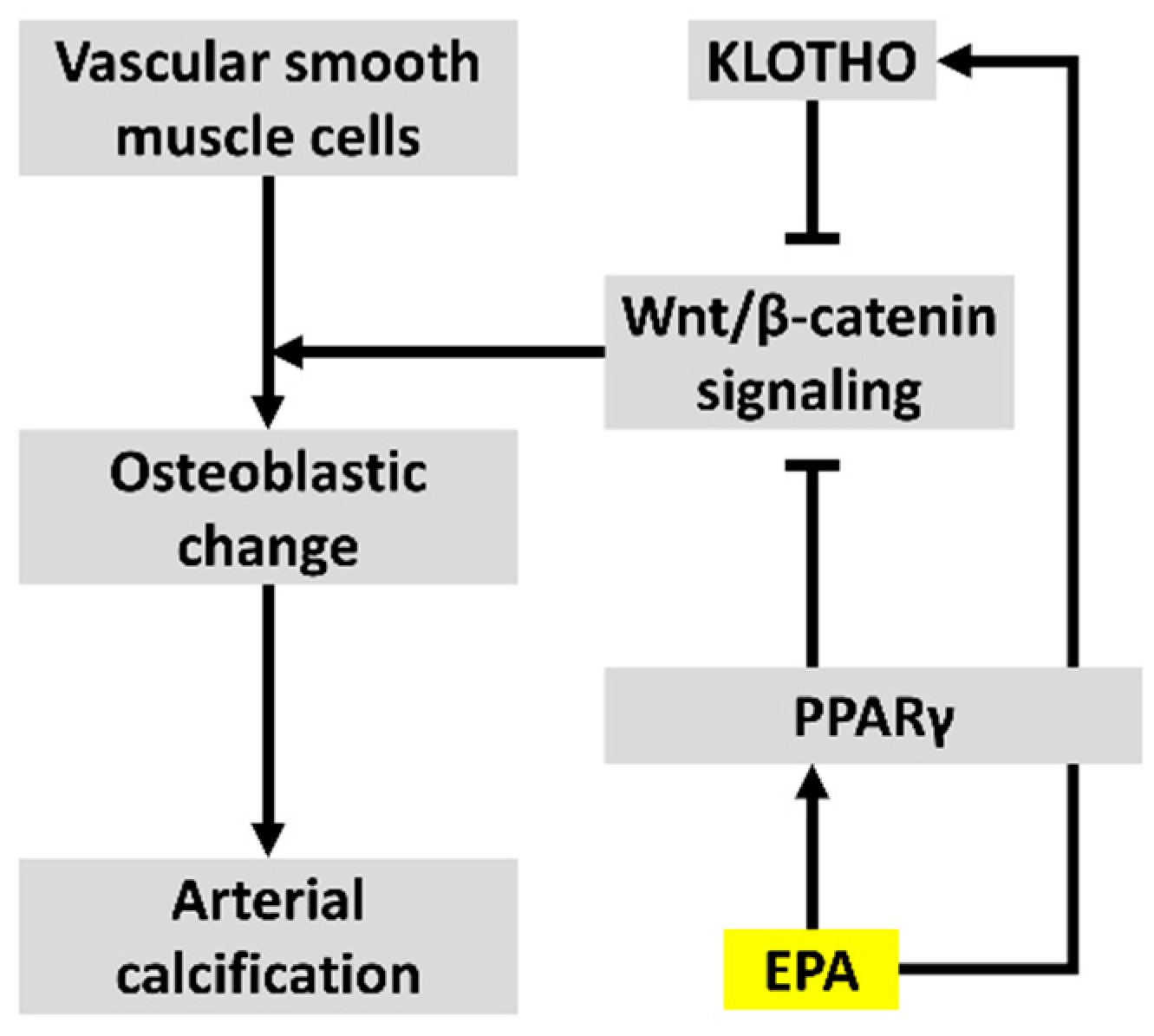
3.4. EPA Suppressed β-Catenin Signaling via PPARγ Activation
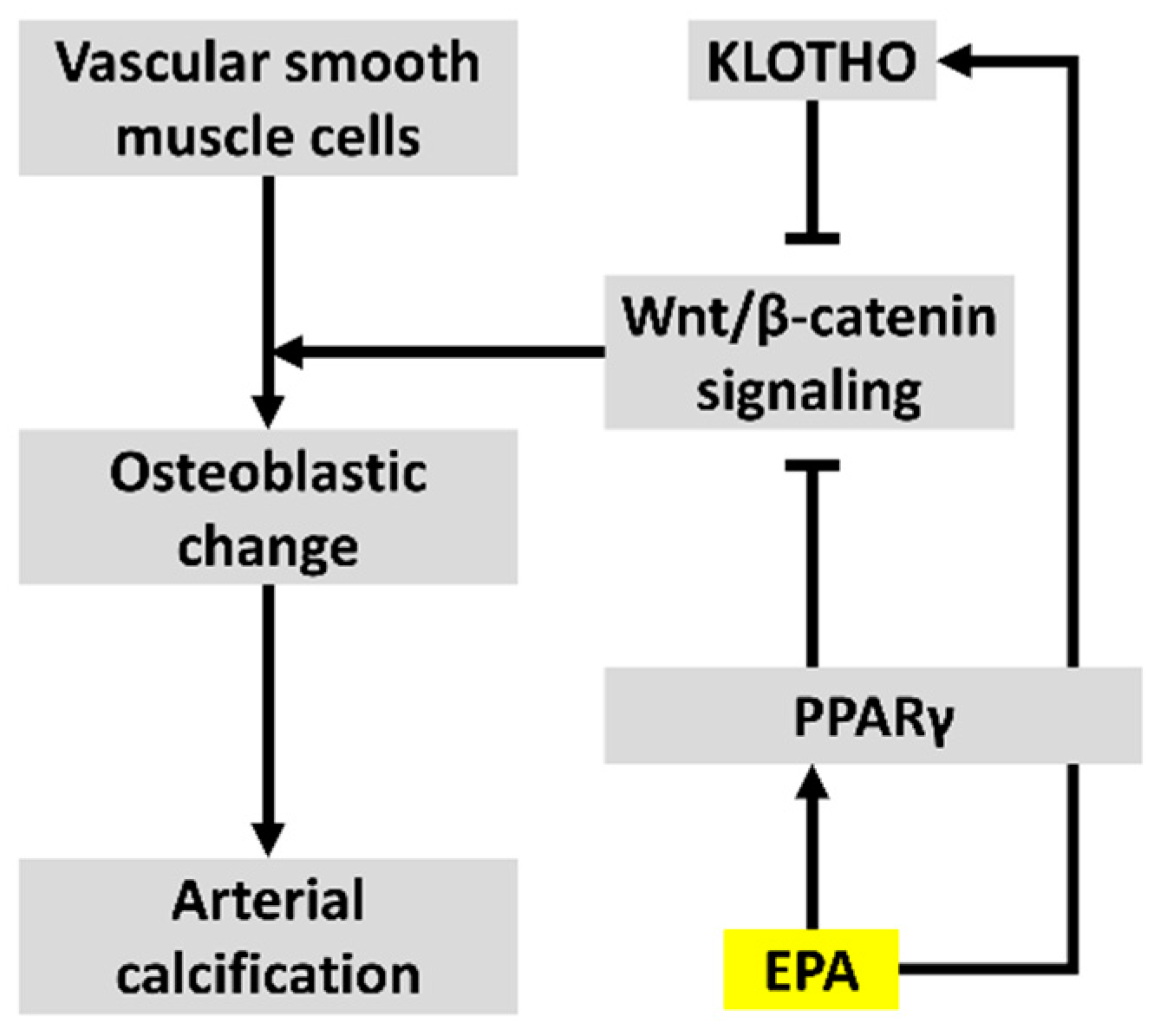
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sangiorgi, G.; Rumberger, J.A.; Severson, A.; Edwards, W.D.; Gregoire, J.; Fitzpatrick, L.A.; Schwartz, R.S. Arterial calcification and not lumen stenosis is highly correlated with atherosclerotic plaque burden in humans: A histologic study of 723 coronary artery segments using nondecalcifying methodology. J. Am. Coll. Cardiol. 1998, 31, 126–133. [Google Scholar] [CrossRef]
- Kelly, R.P.; Tunin, R.; Kass, D.A. Effect of reduced aortic compliance on cardiac efficiency and contractile function of in situ canine left ventricle. Circ. Res. 1992, 71, 490. [Google Scholar] [CrossRef] [PubMed]
- Wayhs, R.; Zelinger, A.; Raggi, P. High coronary artery calcium scores pose an extremely elevated risk for hard events. J. Am. Coll. Cardiol. 2002, 39, 225–230. [Google Scholar] [CrossRef]
- Keelan, P.C.; Bielak, L.F.; Ashai, K.; Jamjoum, L.S.; Denktas, A.E.; Rumberger, J.A.; Sheedy, I.I.P.F.; Peyser, P.A.; Schwartz, R.S. Long-term prognostic value of coronary calcification detected by electron-beam computed tomography in patients undergoing coronary angiography. Circulation 2001, 104, 412. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, S.; Kakihana, M.; Watanabe, H.; Sugishita, Y. Chronically decreased aortic distensibility causes deterioration of coronary perfusion during increased left ventricular contraction. J. Am. Coll. Cardiol. 1994, 24, 1406–1414. [Google Scholar] [CrossRef]
- Yokoyama, M.; Origasa, H.; Matsuzaki, M.; Matsuzawa, Y.; Saito, Y.; Ishikawa, Y.; Oikawa, S.; Sasaki, J.; Hishida, H.; Itakura, H.; et al. Effects of eicosapentaenoic acid on major coronary events in hypercholesterolaemic patients (JELIS): A randomised open-label, blinded endpoint analysis. Lancet 2007, 369, 1090–1098. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Yokoyama, M.; Saito, Y.; Matsuzaki, M.; Origasa, H.; Oikawa, S.; Sasaki, J.; Hishida, H.; Itakura, H.; Kita, T.; et al. Preventive effects of eicosapentaenoic acid on coronary artery disease in patients with peripheral artery disease. Circ. J. 2010, 74, 1451–1457. [Google Scholar] [CrossRef] [PubMed]
- Doi, M.; Nosaka, K.; Miyoshi, T.; Iwamoto, M.; Kajiya, M.; Okawa, K.; Nakayama, R.; Takagi, W.; Takeda, K.; Hirohata, S.; et al. Early eicosapentaenoic acid treatment after percutaneous coronary intervention reduces acute inflammatory responses and ventricular arrhythmias in patients with acute myocardial infarction: A randomized, controlled study. Int. J. Cardiol. 2014, 176, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Nosaka, K.; Miyoshi, T.; Iwamoto, M.; Kajiya, M.; Okawa, K.; Tsukuda, S.; Yokohama, F.; Sogo, M.; Nishibe, T.; Matsuo, N.; et al. Early initiation of eicosapentaenoic acid and statin treatment is associated with better clinical outcomes than statin alone in patients with acute coronary syndromes: 1-Year outcomes of a randomized controlled study. Int. J. Cardiol. 2017, 228, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Kanai, S.; Uto, K.; Honda, K.; Hagiwara, N.; Oda, H. Eicosapentaenoic acid reduces warfarin-induced arterial calcification in rats. Atherosclerosis 2011, 215, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Eguchi, K.; Matsumoto, S.; Fujiu, K.; Komuro, I.; Nagai, R.; Manabe, I. The ω-3 polyunsaturated fatty acid, eicosapentaenoic acid, attenuates abdominal aortic aneurysm development via suppression of tissue remodeling. PLoS ONE 2014, 9, e96286. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Miura, D.; Saito, Y.; Yunoki, K.; Koyama, Y.; Satoh, M.; Kondo, M.; Osawa, K.; Hatipoglu, O.F.; Miyoshi, T.; et al. Eicosapentaenoic acid prevents arterial calcification in klotho mutant mice. PLoS ONE 2017, 12, e0181009. [Google Scholar] [CrossRef] [PubMed]
- Polonsky, T.S.; McClelland, R.L.; Jorgensen, N.W.; Bild, D.E.; Burke, G.L.; Guerci, A.D.; Greenland, P. Coronary artery calcium score and risk classification for coronary heart disease prediction. JAMA 2010, 303, 1610–1616. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, U.; Massaro, J.M.; D’Agostino, R.B., Sr.; Kathiresan, S.; Fox, C.S.; O’Donnell, C.J. Cardiovascular event prediction and risk reclassification by coronary, aortic, and valvular calcification in the framingham heart study. J. Am. Heart Assoc. 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.; Baarsma, H.A.; Konigshoff, M. Wnt signaling in lung aging and disease. Ann. Am. Thorac. Soc. 2016, 13, S411–S416. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.C.; Fogo, A.B. Fibrosis and renal aging. Kidney Int. 2014, 4, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Brack, A.S.; Conboy, M.J.; Roy, S.; Lee, M.; Kuo, C.J.; Keller, C.; Rando, T.A. Increased wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science 2007, 317, 807. [Google Scholar] [CrossRef] [PubMed]
- Freise, C.; Kretzschmar, N.; Querfeld, U. Wnt signaling contributes to vascular calcification by induction of matrix metalloproteinases. BMC Cardiovasc. Disord. 2016, 16, 185. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.S.; Cheng, S.L.; Pingsterhaus, J.M.; Charlton-Kachigian, N.; Loewy, A.P.; Towler, D.A. Msx2 promotes cardiovascular calcification by activating paracrine wnt signals. J. Clin. Investig. 2005, 115, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Sun, D.; Duan, Y.; Wen, P.; Dai, C.; Yang, J.; He, W. Wnt/beta-catenin signaling promotes VSMCS to osteogenic transdifferentiation and calcification through directly modulating Runx2 gene expression. Exp. Cell Res. 2016, 345, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Kuro-o, M.; Matsumura, Y.; Aizawa, H.; Kawaguchi, H.; Suga, T.; Utsugi, T.; Ohyama, Y.; Kurabayashi, M.; Kaname, T.; Kume, E.; et al. Mutation of the mouse klotho gene leads to a syndrome resembling ageing. Nature 1997, 390, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fergusson, M.M.; Castilho, R.M.; Liu, J.; Cao, L.; Chen, J.; Malide, D.; Rovira, I.I.; Schimel, D.; Kuo, C.J.; et al. Augmented Wnt signaling in a mammalian model of accelerated aging. Science 2007, 317, 803. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, K.; van der Velde, M.; Astor, B.C.; Woodward, M.; Levey, A.S.; de Jong, P.E.; Coresh, J.; Gansevoort, R.T. Association of estimated glomerular filtration rate and albuminuria with all-cause and cardiovascular mortality: A collaborative meta-analysis of general population cohorts. Lancet 2010, 375, 2073–2081. [Google Scholar] [PubMed]
- Van der Velde, M.; Matsushita, K.; Coresh, J.; Astor, B.C.; Woodward, M.; Levey, A.; de Jong, P.; Gansevoort, R.T.; Chronic Kidney Disease Prognosis Consortium; van der Velde, M.; et al. Lower estimated glomerular filtration rate and higher albuminuria are associated with all-cause and cardiovascular mortality. A collaborative meta-analysis of high-risk population cohorts. Kidney Int. 2011, 79, 1341–1352. [Google Scholar] [PubMed]
- Hu, M.C.; Kuro-o, M.; Moe, O.W. Secreted klotho and chronic kidney disease. Adv. Exp. Med. Biol. 2012, 728, 126–157. [Google Scholar] [PubMed]
- Zhou, L.; Li, Y.; Zhou, D.; Tan, R.J.; Liu, Y. Loss of klotho contributes to kidney injury by derepression of Wnt/Beta-catenin signaling. J. Am. Soc. Nephrol. 2013, 24, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Aizawa, H.; Shiraki-Iida, T.; Nagai, R.; Kuro-o, M.; Nabeshima, Y.-I. Identification of the human klotho gene and its two transcripts encoding membrane and secreted klotho protein. Biochem. Biophys. Res. Commun. 1998, 242, 626–630. [Google Scholar] [CrossRef] [PubMed]
- Shiraki-Iida, T.; Aizawa, H.; Matsumura, Y.; Sekine, S.; Iida, A.; Anazawa, H.; Nagai, R.; Kuro-o, M.; Nabeshima, Y.-I. Structure of the mouse klotho gene and its two transcripts encoding membrane and secreted protein 1. FEBS Lett. 1998, 424, 6–10. [Google Scholar] [CrossRef]
- Ritter, C.S.; Zhang, S.; Delmez, J.; Finch, J.L.; Slatopolsky, E. Differential expression and regulation of klotho by paricalcitol in the kidney, parathyroid, and aorta of uremic rats. Kidney Int. 2015, 87, 1141–1152. [Google Scholar] [CrossRef] [PubMed]
- Mencke, R.; Harms, G.; Mirkovic, K.; Struik, J.; Van Ark, J.; Van Loon, E.; Verkaik, M.; De Borst, M.H.; Zeebregts, C.J.; Hoenderop, J.G.; et al. Membrane-bound klotho is not expressed endogenously in healthy or uraemic human vascular tissue. Cardiovasc. Res. 2015, 108, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, K.; Olauson, H.; Amin, R.; Ponnusamy, A.; Goetz, R.; Taylor, R.F.; Mohammadi, M.; Canfield, A.; Kublickiene, K.; Larsson, T.E. Arterial klotho expression and FGF23 effects on vascular calcification and function. PLoS ONE 2013, 8, e60658. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S.; Crooks, H.; Dracheva, T.; Nishanian, T.G.; Singh, B.; Jen, J.; Waldman, T. Oncogenic β-catenin is required for bone morphogenetic protein 4 expression in human cancer cells. Cancer Res. 2002, 62, 2744. [Google Scholar] [PubMed]
- Panizo, S.; Cardus, A.; Encinas, M.; Parisi, E.; Valcheva, P.; Lopez-Ongil, S.; Coll, B.; Fernandez, E.; Valdivielso, J.M. Rankl increases vascular smooth muscle cell calcification through a rank-bmp4-dependent pathway. Circ. Res. 2009, 104, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.; Lu, T.S.; Molostvov, G.; Lee, C.; Lam, F.T.; Zehnder, D.; Hsiao, L.L. Vascular klotho deficiency potentiates the development of human artery calcification and mediates resistance to fibroblast growth factor 23. Circulation 2012, 125, 2243–2255. [Google Scholar] [CrossRef] [PubMed]
- Voelkl, J.; Alesutan, I.; Leibrock, C.B.; Quintanilla-Martinez, L.; Kuhn, V.; Feger, M.; Mia, S.; Ahmed, M.S.; Rosenblatt, K.P.; Kuro, O.M.; et al. Spironolactone ameliorates pit1-dependent vascular osteoinduction in klotho-hypomorphic mice. J. Clin. Investig. 2013, 123, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.U.; Ohmori, K.; Konishi, K.; Igarashi, J.; Hashimoto, T.; Kamitori, K.; Yamaguchi, F.; Tsukamoto, I.; Uyama, T.; Ishihara, Y.; et al. Eicosapentaenoic acid upregulates VEGF-A through both GPR120 and PPARγ mediated pathways in 3T3-L1 adipocytes. Mol. Cell Endocrinol. 2015, 406, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, Y.; Fan, Y.; Wu, J.; Zhao, B.; Guan, Y.; Chien, S.; Wang, N. Klotho is a target gene of PPAR-γ. Kidney Int. 2008, 74, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.E.; Lambert, M.H.; Montana, V.G.; Parks, D.J.; Blanchard, S.G.; Brown, P.J.; Sternbach, D.D.; Lehmann, J.M.; Wisely, G.B.; Willson, T.M.; et al. Molecular recognition of fatty acids by peroxisome proliferator—Activated receptors. Mol. Cell 1999, 3, 397–403. [Google Scholar] [CrossRef]
- Moe, S.M.; Chen, N.X. Mechanisms of vascular calcification in chronic kidney disease. J. Am. Soc. Nephrol. 2008, 19, 213–216. [Google Scholar] [CrossRef] [PubMed]
- De Montes Oca, A.; Guerrero, F.; Martinez-Moreno, J.M.; Madueno, J.A.; Herencia, C.; Peralta, A.; Almaden, Y.; Lopez, I.; Aguilera-Tejero, E.; Gundlach, K.; et al. Magnesium inhibits Wnt/beta-catenin activity and reverses the osteogenic transformation of vascular smooth muscle cells. PLoS ONE 2014, 9, e89525. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, A.; Matsui, H.; Ohta, M.; Sambuichi, K.; Kawano, H.; Notsu, T.; Imada, K.; Yokoyama, T.; Kurabayashi, M. Palmitic acid induces osteoblastic differentiation in vascular smooth muscle cells through ACSL3 and NF-kB, novel targets of eicosapentaenoic acid. PLoS ONE 2013, 8, e68197. [Google Scholar] [CrossRef] [PubMed]
- Song, K.-S.; Jing, K.; Kim, J.-S.; Yun, E.-J.; Shin, S.; Seo, K.-S.; Park, J.-H.; Heo, J.-Y.; Kang, J.X.; Suh, K.-S.; et al. Omega-3-polyunsaturated fatty acids suppress pancreatic cancer cell growth in vitro and in vivo via downregulation of Wnt/Beta-catenin signaling. Pancreatology 2011, 11, 574–584. [Google Scholar] [CrossRef] [PubMed]
- D’Eliseo, D.; Velotti, F. Omega-3 fatty acids and cancer cell cytotoxicity: Implications for multi-targeted cancer therapy. J. Clin. Med. 2016, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- De Carlo, F.; Witte, T.R.; Hardman, W.E.; Claudio, P.P. Omega-3 eicosapentaenoic acid decreases CD133 colon cancer stem-like cell marker expression while increasing sensitivity to chemotherapy. PLoS ONE 2013, 8, e69760. [Google Scholar] [CrossRef] [PubMed]
- Saremi, A.; Bahn, G.; Reaven, P.D.; VADT Investiqators. Progression of vascular calcification is increased with statin use in the Veterans Affairs Diabetes Trial (VADT). Diabetes Care 2012, 35, 2390–2392. [Google Scholar] [CrossRef] [PubMed]
- Henein, M.; Granåsen, G.; Wiklund, U.; Schmermund, A.; Guerci, A.; Erbel, R.; Raggi, P. High dose and long-term statin therapy accelerate coronary artery calcification. Int. J. Cardiol. 2015, 184, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Nishio, R.; Shinke, T.; Otake, H.; Nakagawa, M.; Nagoshi, R.; Inoue, T.; Kozuki, A.; Hariki, H.; Osue, T.; Taniguchi, Y.; et al. Stabilizing effect of combined eicosapentaenoic acid and statin therapy on coronary thin-cap fibroatheroma. Atherosclerosis 2014, 234, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Niki, T.; Wakatsuki, T.; Yamaguchi, K.; Taketani, Y.; Oeduka, H.; Kusunose, K.; Ise, T.; Iwase, T.; Yamada, H.; Soeki, T.; et al. Effects of the addition of eicosapentaenoic acid to strong statin therapy on inflammatory cytokines and coronary plaque components assessed by integrated backscatter intravascular ultrasound. Circ. J. 2016, 80, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.L.; Lamon-Fava, S.; Matthan, N.R.; Wu, D.; Lichtenstein, A.H. EPA and DHA exposure alters the inflammatory response but not the surface expression of Toll-like receptor 4 in macrophages. Lipids 2015, 50, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, D.; Lamon-Fava, S.; Matthan, N.R.; Honda, K.L.; Lichtenstein, A.H. In vitro fatty acid enrichment of macrophages alters inflammatory response and net cholesterol accumulation. Br. J. Nutr. 2009, 102, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T.; Naoe, S.; Nakakuki, M.; Kawano, H.; Imada, K. Eicosapentaenoic acid prevents saturated fatty acid-induced vascular endothelial dysfunction: Involvement of long-chain acyl-Coa synthetase. J. Atheroscler. Thromb. 2015, 22, 1172–1185. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Hottiger, M.O. Crosstalk between Wnt/Beta-catenin and NF-kB signaling pathway during inflammation. Front. Immunol. 2016, 7, 378. [Google Scholar] [CrossRef] [PubMed]
- Sumida, T.; Naito, A.T.; Nomura, S.; Nakagawa, A.; Higo, T.; Hashimoto, A.; Okada, K.; Sakai, T.; Ito, M.; Yamaguchi, T.; et al. Complement C1q-induced activation of beta-catenin signalling causes hypertensive arterial remodelling. Nat. Commun. 2015, 6, 6241. [Google Scholar] [CrossRef] [PubMed]
- Matthijs Blankesteijn, W.; Hermans, K.C. Wnt signaling in atherosclerosis. Eur. J. Pharmacol. 2015, 763, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Dyall, S.C. Long-chain omega-3 fatty acids and the brain: A review of the independent and shared effects of EPA, DPA and DHA. Front. Aging Neurosci. 2015, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Duvall, M.G.; Levy, B.D. DHA- and EPA-derived resolvins, protectins, and maresins in airway inflammation. Eur. J. Pharmacol. 2016, 785, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.F.; Pillai, P.S.; Recchiuti, A.; Yang, R.; Serhan, C.N. Pro-resolving actions and stereoselective biosynthesis of 18S E-series resolvins in human leukocytes and murine inflammation. J. Clin. Investig. 2011, 121, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Basil, M.C.; Levy, B.D. Specialized pro-resolving mediators: Endogenous regulators of infection and inflammation. Nat. Rev. Immunol. 2016, 16, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Romano, M.; Cianci, E.; Simiele, F.; Recchiuti, A. Lipoxins and aspirin-triggered lipoxins in resolution of inflammation. Eur. J. Pharmacol. 2015, 760, 49–63. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | Annealing Temperature (°C) | Product Size (bp) | ||
|---|---|---|---|---|
| Mouse | ||||
| Klotho | forward | CAAAGTCTTCGGCCTTGTTC | 60 | 111 |
| reverse | CTCCCCAAGCAAAGTCACA | |||
| Pparg | forward | ATCATCTACACGATGCTGGCC | 60 | 81 |
| reverse | CTCCCTGGTCATGAATCCTTG | |||
| Ffar4 | forward | TGCCCCTCTGCATCTTGTTC | 60 | 202 |
| reverse | CGCGATGCTTTCGTGATCTG | |||
| Actb | forward | GGAGGGGGTTGAGGTGTT | 60 | 70 |
| reverse | GTGTGCACTTTTATTGGTCTCAA | |||
| Human | ||||
| AXIN2 | forward | AGTGTGAGGTCCACGGAAAC | 58 | 103 |
| reverse | CTGGTGCAAAGACATAGCCA | |||
| LEF1 | forward | AATGAGAGCGAATGTCGTTGC | 60 | 137 |
| reverse | GCTGTCTTTCTTTCCGTGCTA | |||
| RUNX2 | forward | TCTTAGAACAAATTCTGCCCTTT | 58 | 136 |
| reverse | TGCTTTGGTCTTGAAATCACA | |||
| BMP4 | forward | GATCCACAGCACTGGTCTTG | 60 | 150 |
| reverse | GGGATGCTGCTGAGGTTAAA | |||
| PPARG | forward | GGCTTCATGACAAGGGAGTTTC | 60 | 74 |
| reverse | AACTCAAACTTGGGCTCCATAAAG | |||
| MYH11 | forward | AGATGGTTCTGAGGAGGAAACG | 60 | 85 |
| reverse | AAAACTGTAGAAAGTTGCTTATTCACT | |||
| FFAR4 | forward | CTGTGCAGGAATGAGTGGAAG | 60 | 197 |
| reverse | CTGATGGAGGGTACTGGAAATG | |||
| GAPDH | forward | GCGAGATCCCTCCAAAATCAA | 58 | 172 |
| reverse | GTTCACACCCATGACGAACAT |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saito, Y.; Nakamura, K.; Miura, D.; Yunoki, K.; Miyoshi, T.; Yoshida, M.; Kawakita, N.; Kimura, T.; Kondo, M.; Sarashina, T.; et al. Suppression of Wnt Signaling and Osteogenic Changes in Vascular Smooth Muscle Cells by Eicosapentaenoic Acid. Nutrients 2017, 9, 858. https://doi.org/10.3390/nu9080858
Saito Y, Nakamura K, Miura D, Yunoki K, Miyoshi T, Yoshida M, Kawakita N, Kimura T, Kondo M, Sarashina T, et al. Suppression of Wnt Signaling and Osteogenic Changes in Vascular Smooth Muscle Cells by Eicosapentaenoic Acid. Nutrients. 2017; 9(8):858. https://doi.org/10.3390/nu9080858
Chicago/Turabian StyleSaito, Yukihiro, Kazufumi Nakamura, Daiji Miura, Kei Yunoki, Toru Miyoshi, Masashi Yoshida, Norifumi Kawakita, Tomonari Kimura, Megumi Kondo, Toshihiro Sarashina, and et al. 2017. "Suppression of Wnt Signaling and Osteogenic Changes in Vascular Smooth Muscle Cells by Eicosapentaenoic Acid" Nutrients 9, no. 8: 858. https://doi.org/10.3390/nu9080858