Effects of Zearalenone Exposure on the TGF-β1/Smad3 Signaling Pathway and the Expression of Proliferation or Apoptosis Related Genes of Post-Weaning Gilts

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Relative mRNA Expressions of the PCNA, BAX, BCL-2, TGF-β1, and Smad3 in Uteri of Post-Weaning Gilts
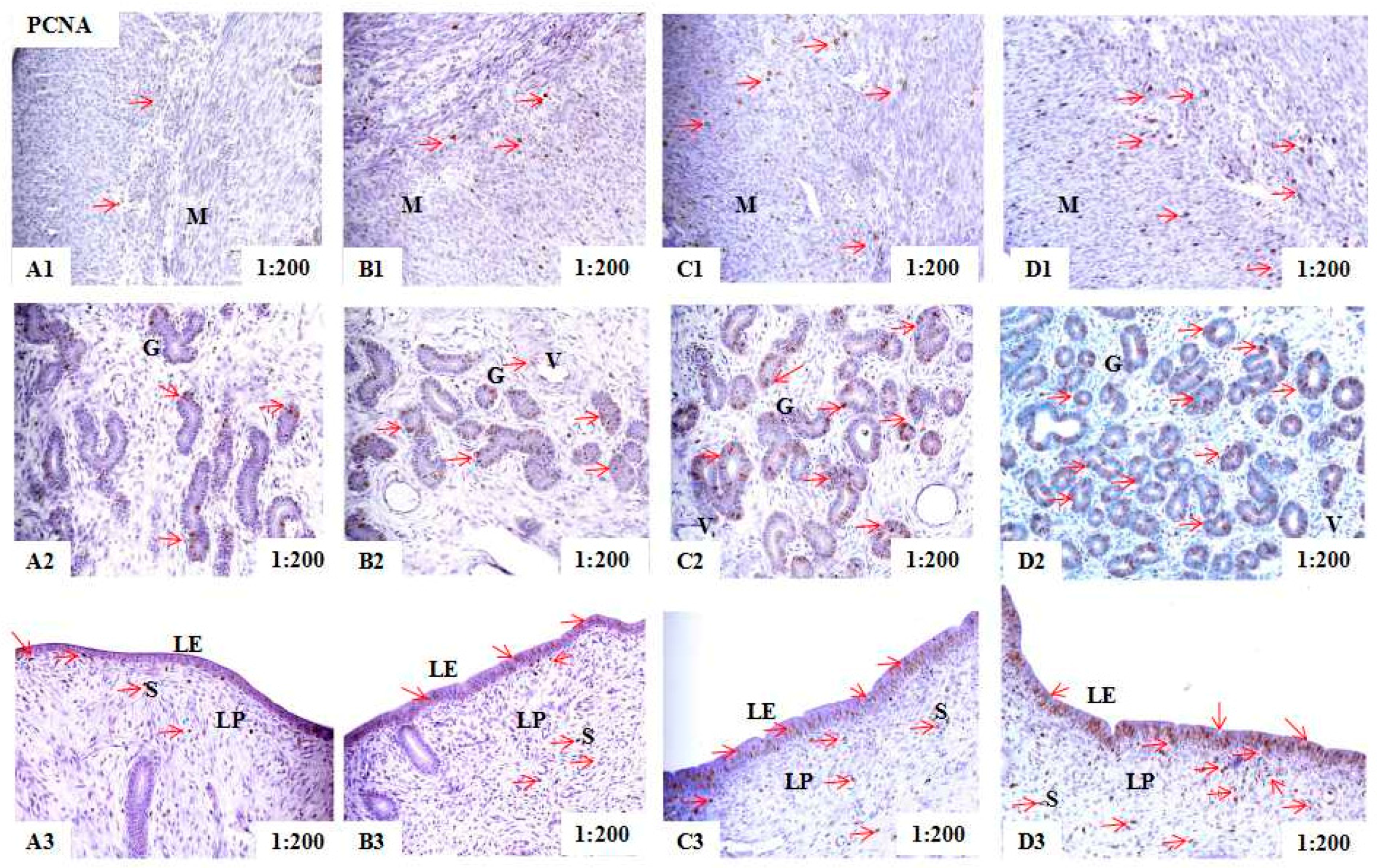
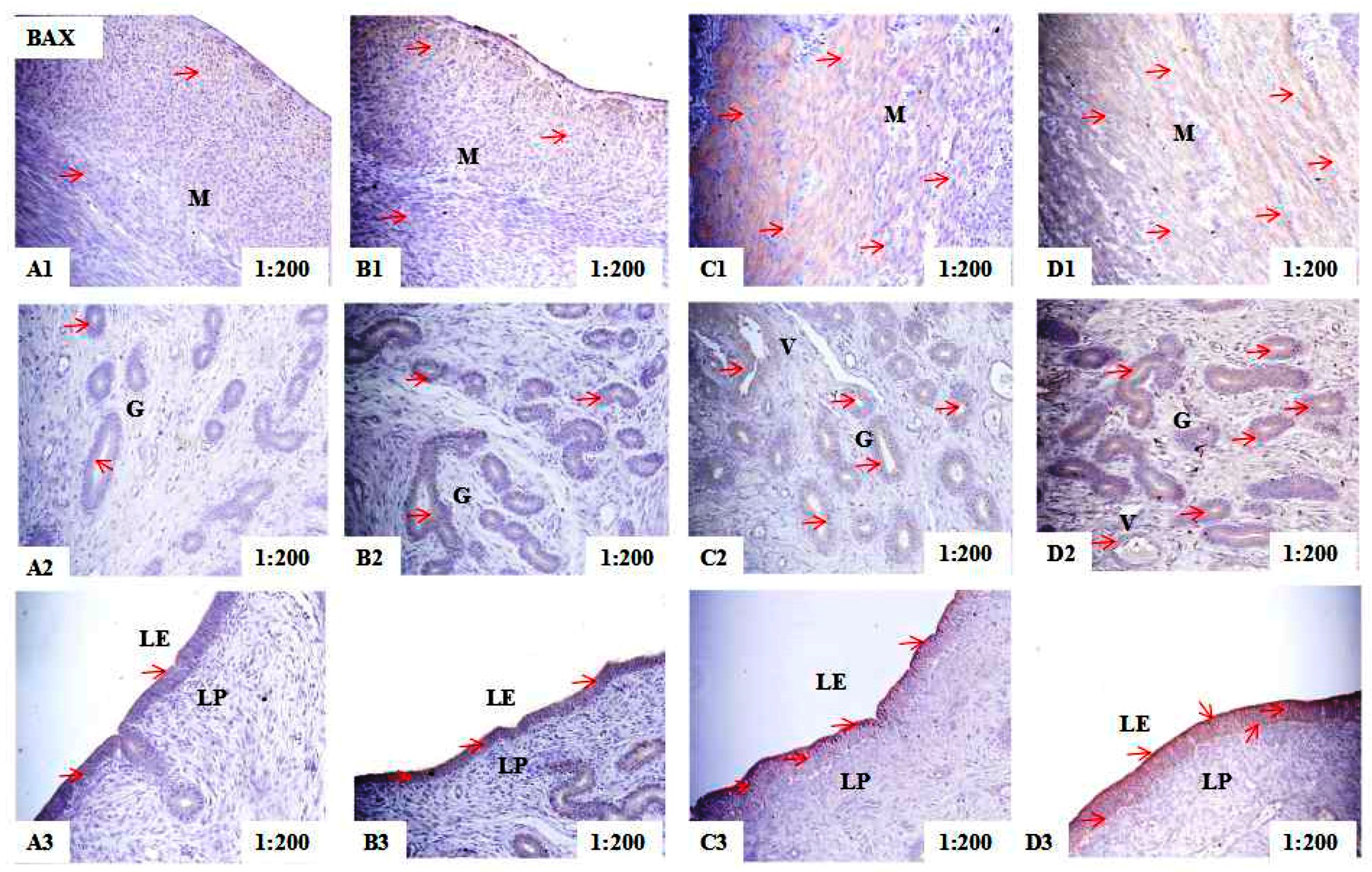
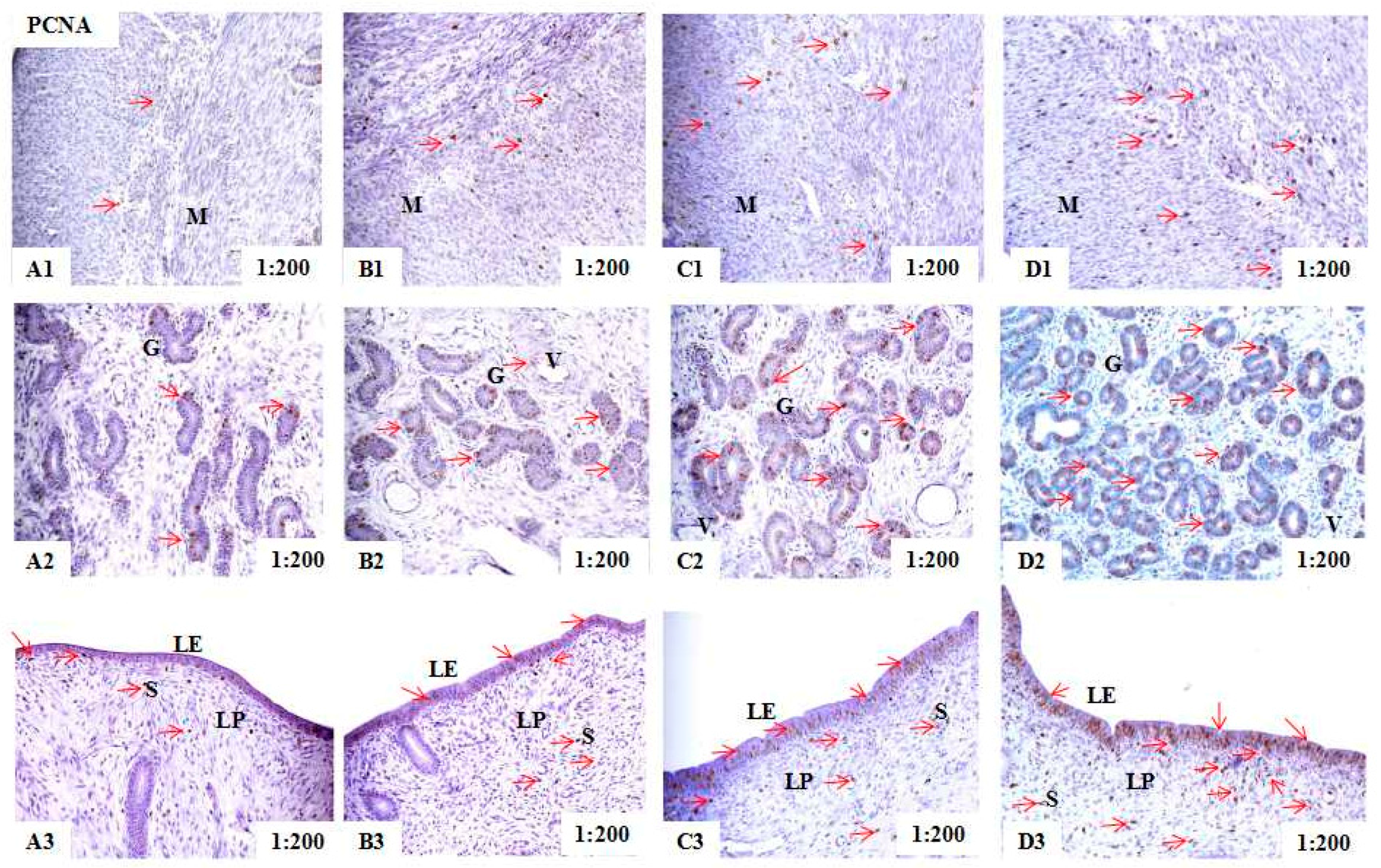
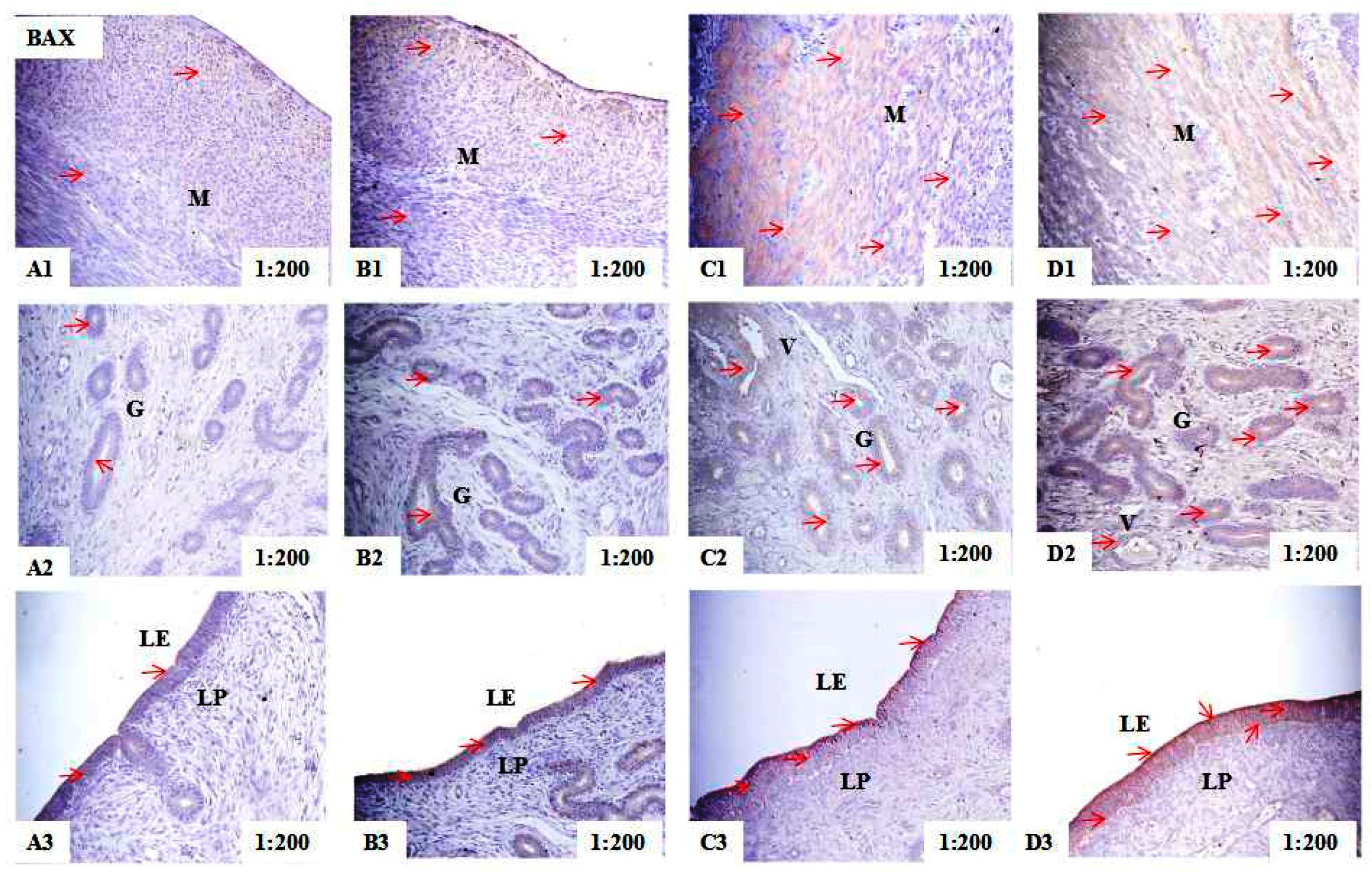
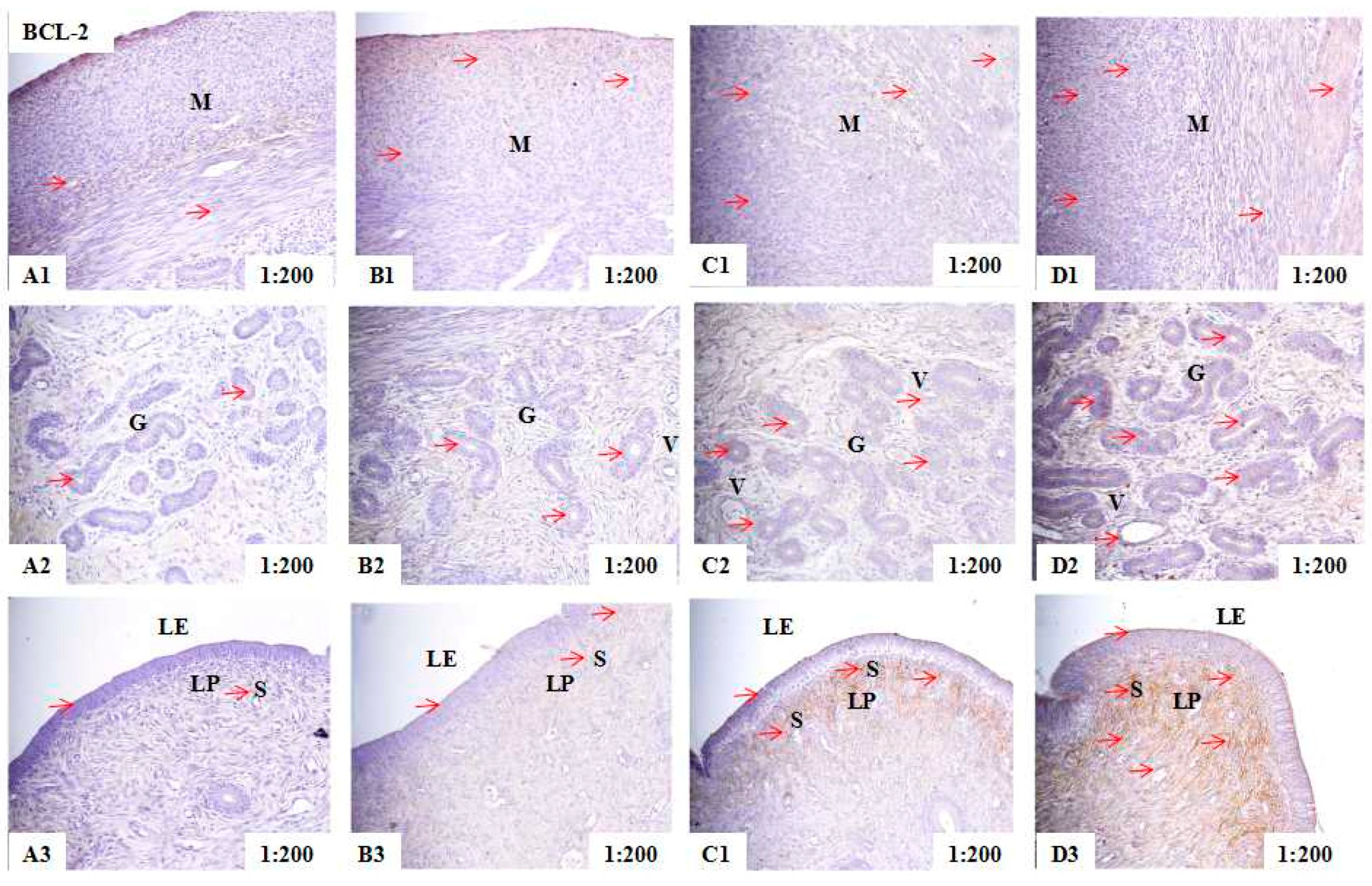
2.2. Localization of PCNA, BAX, and BCL-2 Immunoreactivity in Uteri of Post-Weaning Gilts
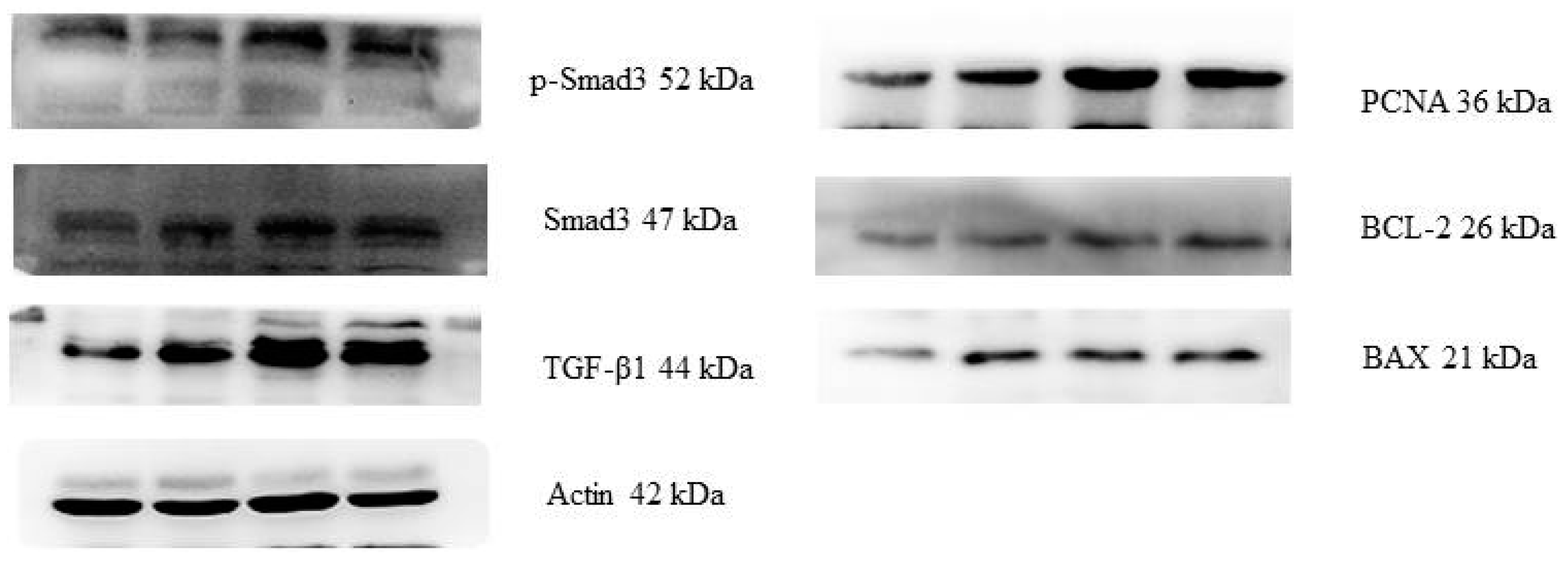
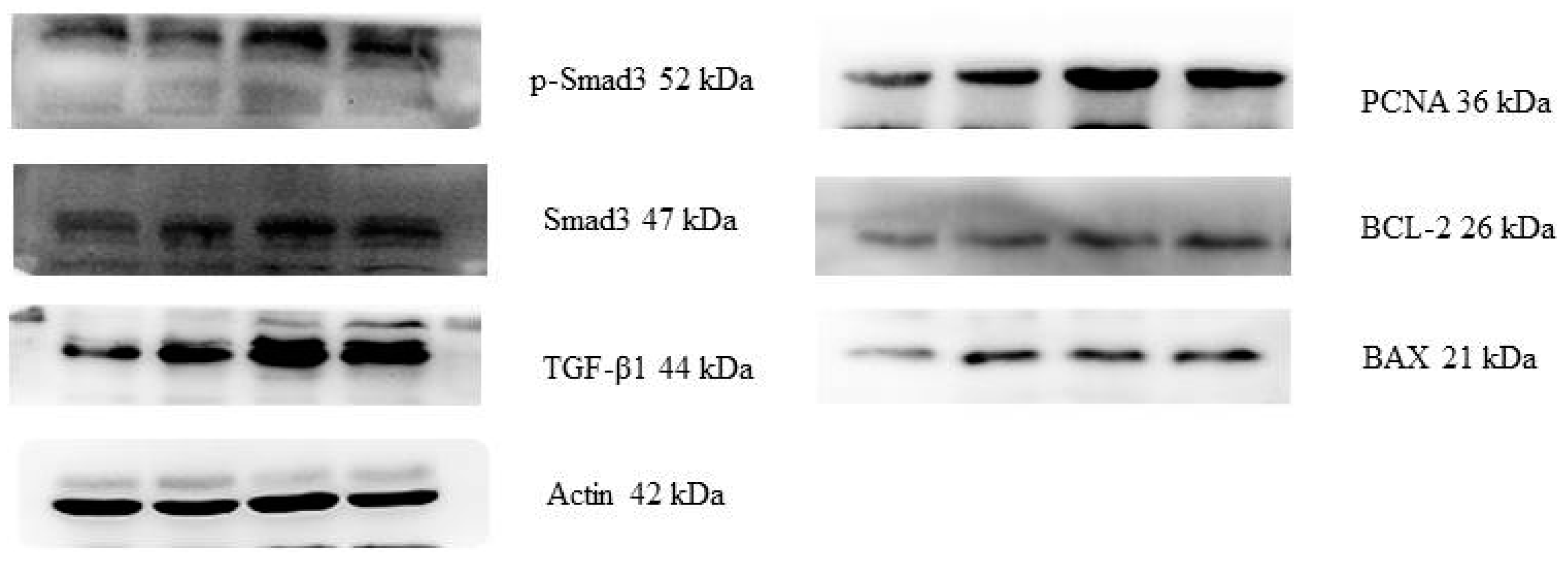
2.3. Protein Expressions of the PCNA, BAX, BCL-2, TGF-β1, Smad3 and p-Smad3
3. Discussion
4. Materials and Methods
4.1. Preparation of Zearalenone-Contaminated Diet
4.2. Animals, Treatments and Feeding Management
4.3. Sample Collection
4.4. Total RNA Extraction, cDNA Preparation, and Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.5. Immunohistochemistry
4.6. Measurement of the Integrated Optical Density of the PCNA, BAX, and BCL-2 Immunohistochemistry
4.7. Western Blotting
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Luo, X.H.; Qi, L.J.; Liu, Y.T.; Wang, R.; Yang, D.; Li, K.; Wang, L.; Li, Y.N.; Zhang, Y.W.; Chen, Z.X. Effects of electron beam irradiation on zearalenone and ochratoxin a in naturally contaminated corn and corn quality parameters. Toxins 2017, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Zaki, M. Mycotoxins in animals: Occurrence, effects, prevention and management. J. Toxicol. Environ. Health Sci. 2012, 4, 13–28. [Google Scholar] [CrossRef]
- Hueza, I.M.; Raspantini, P.C.F.; Raspantini, L.E.R.; Latorre, A.O.; Górniak, S.L. Zearalenone, an estrogenic mycotoxin, is an immunotoxic compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangsila, A.; Faucet-Marquis, V.; Pfohl-Leszkowicz, A.; Itsaranuwat, P. Detoxification of zearalenone by Lactobacillus pentosus strains. Food Control 2016, 62, 187–192. [Google Scholar] [CrossRef]
- Kohei, M.; Yasushi, N.; Naoki, M. Metabolism of zearalenone in the course of beer fermentation. Toxins 2011, 3, 134–141. [Google Scholar]
- Fu, G.H.; Ma, J.F.; Wang, L.H.; Yang, X.; Liu, J.; Zhao, X. Effect of Degradation of Zearalenone-Contaminated Feed by Bacillus licheniformis CK1 on Postweaning Female Piglets. Toxins 2016, 8, 300. [Google Scholar] [CrossRef] [PubMed]
- Gromadzka, K.; Waskiewicz, A.; Chelkowski, J.; Golinski, P. Zearalenone and its metabolites: Occurrence, detection, toxicity and guidelines. World Mycotoxin J. 2008, 1, 209–220. [Google Scholar] [CrossRef]
- Kwaśniewska, K.; Gadzała-Kopciuch, R.; Cendrowski, K. Analytical procedure for the determination of zearalenone in environmental and biological samples. Crit. Rev. Anal. Chem. 2015, 45, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, C.; Kersten, S.; Burkhardt-Holm, P.; Valenta, H.; Dänicke, S. Occurrence of deoxynivalenol and zearalenone in commercial fish feed: An initial study. Toxins 2013, 5, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatecka, E.; Ded, L.; Elzeinova, F.; Kubatova, A.; Dorosh, A.; Marqaryan, H.; Dostalova, P.; Korenkova, V.; Hoskova, K.; Peknicova, J. Effect of zearalenone on reproductive parameters and expression of selected testicular genes in mice. Reprod. Toxicol. 2014, 45, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Benzoni, E.; Minervini, F.; Giannoccaro, A.; Fornelli, F.; Vigo, D.; Visconti, A. Influence of in vitro exposure to mycotoxin zearalenone and its derivatives on swine sperm quality. Reprod. Toxicol. 2008, 25, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; Dell’Aquila, M.E. Zearalenone and reproductive function in farm animals. Int. J. Mol. Sci. 2008, 9, 2570–2584. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Rybarczyk, L.; Jakimiuk, E.; Zielonka, Ł.; Obremski, K.; Zwierzchowski, W.; Gajęcki, M. The effect of experimental long-term exposure to low-dose zearalenone on uterine histology in sexually immature gilts. Exp. Toxicol. Pathol. 2012, 64, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.X.; Yang, C.W.; Huang, L.B.; Niu, Q.S.; Jiang, S.Z.; Chi, F. Zearalenone Altered the Serum Hormones, Morphologic and Apoptotic Measurements of Genital Organs in Post-weaning Gilts. Asian-Australas. J. Anim. Sci. 2015, 28, 171–179. [Google Scholar]
- Dai, M.L.; Jiang, S.Z.; Yuan, X.J.; Yang, Z.B.; Yang, W.R.; Huang, L.B. Effects of zearalenone-diet on expression of ghrelin and PCNA genes in ovaries of post-weaning piglets. Anim. Reprod. Sci. 2016, 168, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Clausen, O.P.; Boysen, M. Prognostic value of Bak expression in oral tongue squamous cell carcinomas. Oncol. Rep. 2003, 12, 369–374. [Google Scholar] [CrossRef]
- Cory, S.; Huang, D.C.; Adams, J.M. The Bcl-2 family: Roles in cell survival and oncogenesis. Oncogene 2003, 22, 8590–8607. [Google Scholar] [CrossRef] [PubMed]
- Graves, D. Cytokines that promote periodontal tissue destruction. J. Periodontol. 2008, 79, 1585–1591. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Massagu, J. Mechanisms of TGF-β signaling from cell membrane to the nucleus. Cell 2003, 113, 685–700. [Google Scholar] [CrossRef]
- Jiang, S.Z.; Yang, Z.B.; Yang, W.R.; Gao, J.; Liu, F.X.; Broomhead, J.; Chi, F. Effects of purified zearalenone on growth performance, organ size, serum metabolites, and oxidative stress in postweaning gilts. J. Anim. Sci. 2011, 89, 3008–3015. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Yang, L.J.; Yang, W.R.; Huang, L.B.; Zhou, X.M.; Jiang, S.Z.; Yang, Z.B. Effects of zearalenone on the localization and expression of the growth hormone receptor gene in the uteri of post-weaning piglets. Asian Australas. J. Anim. 2018, 31, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Mackic, S.; Ahmetovic, N. Zearalenone effect on uterine weight of rats. Agric. Conspec. Sci. 2012, 77, 137–141. [Google Scholar]
- Stopa, E.; Babin’Ska, I.; Zielonka, Ł.; Gajecki, M.; Gajecka, M. Immunohistochemical evaluation of apoptosis and proliferation in the mucous membrane of selected uterine regions in pre-pubertal bitches exposed to low doses of zearalenone. Pol. J. Vet. Sci. 2016, 19, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Reinen, J.; Kalma, L.L.; Begheijn, S.; Heus, F.; Commandeur, J.N.; Vermeulen, N.P. Application of cytochrome P450 BM3 mutants as biocatalysts for the profiling of estrogen receptor binding metabolites of the mycotoxin zearalenone. Xenobiotica 2011, 41, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.F.; Zhang, N.Y.; Peng, Y.Z.; Qi, D.S. Interaction of zearalenone and soybean isoflavone on the development of reproductive organs, reproductive hormones and estrogen receptor expression in prepubertal gilts. Anim. Reprod. Sci. 2010, 122, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Rybarczyk, L.; Zwierzchowski, W.; Jakimiuk, E.; Zielonka, L.; Obremski, K.; Gajecki, M. The effect of experimental, long-term exposure to low-dose zearalenone mycotoxicosis on the histological condition of ovaries in sexually immature gilts. Theriogenology 2011, 75, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Obremski, K.; Gajecki, M.; Zwierzchowski, W.; Zielonka, L.; Otrockadomagała, I.; Rotkiewicz, T.; Mikołajczyk, A.; Gajecka, M.; Polak, M. Influence of zearalenone on reproductive system cell proliferation in gilts. Pol. J. Vet. Sci. 2003, 6, 239–245. [Google Scholar] [PubMed]
- Gajecka, M.; Obremski, K.; Jakimiuk, E.; Skorskawyszyn´Ska, E.; Zielonka, Ł.; Gajecki, M. Histopathological examination of ovaries in bitches after experimental zearalenone mycotoxicosis. Pol. J. Vet. Sci. 2008, 11, 363–366. [Google Scholar] [PubMed]
- Yuan, H.; Deng, Y.; Yuan, L.; Wu, J.; Yuan, Z.; Yi, J.; Zhang, M.; Guo, C.; Wen, L.; Li, R.; et al. Gynostemma pentaphyllum protects mouse male germ cells against apoptosis caused by zearalenone via BAX and BCL-2 regulation. Toxicol. Mech. Methods 2010, 20, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhang, L.; Wu, D.; Liu, F. Anti-apoptotic action of zearalenone in MCF-7 cells. Ecotoxicol. Environ. Saf. 2005, 62, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Shao, S.L.; Zhang, L.; Zhang, W.W. The Induction of Apoptosis in Human Gastric Cancer Cell Line SGC-7901 through Upregulation of bax and Downregulation bcl-2 Gene by Allicin. In Proceedings of the 10th International Symposium on Biocontrol and Biotechnology, Harbin, China, 27–30 December 2012. [Google Scholar]
- Baharara, J.; Amini, E. The Potential of Brittle Star Extracted Polysaccharide in Promoting Apoptosis via Intrinsic Signaling Pathway. Avicenna J. Med. Biotechnol. 2015, 7, 151–158. [Google Scholar] [PubMed]
- Soares, N.D.C.P.; Teodoro, A.J.; Oliveira, F.L.; Takiya, C.M.; Junior, A.P.; Nasciutti, L.E.; Lotsch, P.F. Lycopene induce apoptosis in human prostate cells and alters the expression of Bax and Bcl-2 genes. LWT Food Sci. Technol. 2014, 59, 1290–1297. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Gao, R.; Liu, M.; Shi, B.J.; Shan, A.S.; Cheng, B.J. Use of modified halloysite nanotubes in the feed reduces the toxic effects of zearalenone on sow reproduction and piglet development. Theriogenology 2015, 83, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Samik, A.; Safitri, E. Potency of mycotoxin binders on MDA level, expressions of caspase 9 and caspase 3 in the uterus of mice exposed to zearalenone. Iraqi J. Vet. Sci. 2017, 31, 29–33. [Google Scholar]
- Yu, Y.L.; Zhou, H.C.; Sun, W.W.; Zhang, G.S.; Jia, C.; Li, Y.Q. Expression of Bax and Bcl-2 mycotoxicosis of ovaries in SD rats exposured zearalenone. Chin. J. Vet. Sci. 2012, 32, 1015–1019. [Google Scholar]
- Zhang, D.Q.; Zhang, J.J.; Zhong, G.S.; Wang, J.X.; Hu, S.M.; Gao, X.M. Effects of Qilantangzhining Capsule on Hepatocyte Apoptosis and Expression of Bax and Bcl-2 in Rats with Diabetes Mellitus and Hyperlipemia. Chin. J. Tradit. Med. 2005, 20, 211–213. [Google Scholar]
- Cory, S.; Adams, J.M. The Bcl-2 family: Regulations of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Undevia, N.S.; Dorscheid, D.R.; Marroquin, B.A.; Guqliotta, W.L.; Tse, R.; White, S.R. Smad and p38-MAPK signaling mediates apoptotic effects of transforming growth factor-beta1 in human airway epithelial cells. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2004, 287, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.P. Research progress of transforming growth factor beta and regulation of reproductive function. J. Int. Reprod. Health/Fam. Plan. 2003, 22, 17–20. [Google Scholar]
- Nasu, K.; Nishida, M.; Matsumoto, H.; Binq, S.; Inoue, C.; Kawano, Y.; Miyakawa, I. Regulation of proliferation, motility, and contractivity of cultured human endometrial stromal cells by transforming growth factor-beta isoforms. Fertil. Steril. 2005, 84, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Reis, F.M.; Ribeiro, M.F.; Maia, A.L.; Spritzer, P.M. Regulation of human endometrial transforming growth factor beta1 and beta3 isoforms through menstrual cycle and medroxyprogesterone acetate treatment. Histol. Histopathol. 2002, 17, 739–745. [Google Scholar] [PubMed]
- Jiang, X.; Dong, J. Effects of estrogen on proliferation and mRNA expression of TGF-β1 and IGF-1 in osteoblasts. Guide Chian Med. 2013, 11, 88–90. [Google Scholar]
- Ungefroren, H.; Groth, S.; Sebens, S.; Lehnert, H.; Gieseler, F.; Fändrich, F. Differential roles of Smad2 and Smad3 in the regulation of TGF-β1-mediated growth inhibition and cell migration in pancreatic ductal adenocarcinoma cells: Control by Rac1. Mol. Cancer 2011, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Wrighton, K.H.; Lin, X.; Feng, X.H. Phospho-control of TGF-beta superfamily signaling. Cell Res. 2009, 19, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Nasreen, K. TGF-β1 increases proliferation of airway smooth muscle cells by phosphorylation of map kinases. Respir. Res. 2006, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J. Research on the Regulation of TGF-β1/Smad3 Signal Transduction Pathway on Derma Fibroblast during the Wound Healing Process; Second Military Medical University: Shanghai, China, 2007. [Google Scholar]
- Usa, N.C. Nutrient Requirements of Swine; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- AOAC. Official Methods of Analysis of the AOAC; AOAC International: Rockville, MD, USA, 2012. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.; Agnati, L.F.; Horvath, T.L.; Valderrama, J.J.; Calle, A.D.L.; Fuxe, K. Uncoupling protein 2/3 immunoreactivity and theascending dopaminergic and noradrenergic neuronal systems: Relevance for volume transmission. Neuroscience 2006, 137, 1447–1461. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | PCNA | BAX | BCL-2 | TGF-β1 | Smad3 | |
|---|---|---|---|---|---|---|
| Control | 1.00 ± 0.13 c | 1.00 ± 0.11 b | 1.00 ± 0.12 b | 1.00 ± 0.10 c | 1.00 ± 0.15 b | |
| ZEA0.5 | 1.53 ± 0.19 b | 1.23 ± 0.21 ab | 1.19 ± 0.08 ab | 1.45 ± 0.32 b | 1.27 ± 0.08 a | |
| ZEA1.0 | 2.27 ± 0.14 a | 1.37 ± 0.16 ab | 1.29 ± 0.21 a | 1.99 ± 0.09 a | 1.30 ± 0.24 a | |
| ZEA1.5 | 2.38 ± 0.31 a | 1.53 ± 0.42 a | 1.41 ± 0.25 a | 1.85 ± 0.17 a | 1.34 ± 0.07 a | |
| p-values | Treatment | <0.001 | 0.043 | <0.001 | 0.036 | 0.045 |
| Linear | <0.001 | 0.019 | <0.001 | 0.346 | 0.012 | |
| Quadratic | <0.001 | 0.067 | <0.001 | 0.649 | 0.018 | |
| Items | PCNA | BAX | BCL-2 | |
|---|---|---|---|---|
| Control | 63.18 ± 2.11 d | 50.84 ± 2.09 d | 51.35 ± 1.10 d | |
| ZEA0.5 | 71.41 ± 3.16 c | 55.19 ± 3.08 c | 54.40 ± 2.32 c | |
| ZEA1.0 | 90.08 ± 5.16 b | 59.22 ± 4.21 b | 63.19 ± 4.02 b | |
| ZEA1.5 | 95.53 ± 4.42 a | 63.41 ± 3.25 a | 68.05 ± 3.17 a | |
| p-values | Treatment | 0.013 | 0.032 | 0.022 |
| Linear | <0.001 | <0.001 | <0.001 | |
| Quadratic | <0.001 | 0.011 | 0.039 | |
| Items | PCNA/Actin | BAX/Actin | BCL-2/Actin | TGF-β1/Actin | Smad3/Actin | p-Smad3/Actin | |
|---|---|---|---|---|---|---|---|
| Control | 0.27 ± 0.02 c | 0.13 ± 0.02 c | 0.11 ± 0.01 c | 0.39 ± 0.15 b | 0.32 ± 0.01 c | 0.24 ± 0.05 d | |
| ZEA0.5 | 0.62 ± 0.09 b | 0.35 ± 0.03 b | 0.27 ± 0.02 b | 0.91 ± 0.06 a | 0.55 ± 0.03 b | 0.31 ± 0.08 c | |
| ZEA1.0 | 0.95 ± 0.10 a | 0.43 ± 0.11 ab | 0.36 ± 0.01 a | 0.99 ± 0.07 a | 0.82 ± 0.02 a | 0.49 ± 0.06 b | |
| ZEA1.5 | 1.06 ± 0.08 a | 0.46 ± 0.14 a | 0.45 ± 0.12 a | 0.92 ± 0.05 a | 0.75 ± 0.03 a | 0.61 ± 0.02 a | |
| p-values | Treatment | <0.001 | 0.002 | <0.001 | <0.001 | <0.001 | <0.001 |
| Linear | <0.001 | <0.001 | <0.001 | 0.001 | 0.014 | <0.001 | |
| Quadratic | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Ingredients | Content (%) | Nutrients (3) | |
|---|---|---|---|
| Corn | 64.5 | Digestible Energy, MJ/kg | 13.81 |
| Whey powder | 5.0 | Crude Protein (%) | 19.82 |
| Soybean meal | 23.0 | Calcium (%) | 0.70 |
| Fish meal | 5.0 | Total Phosphorus (%) | 0.64 |
| L-Lysine HCl | 0.2 | Lysine (%) | 1.22 |
| CaHPO4 | 0.7 | Sulfur Amino Acid (%) | 0.65 |
| Pulverized Limestone | 0.3 | Threonine (%) | 0.75 |
| NaCl | 0.3 | Trptophan (%) | 0.22 |
| Premix (2) | 1.0 | ||
| Total | 100.0 | ||
| Target Gene | Accession No. | Primer Sequence (5′ to 3′) | Product Size Bp |
|---|---|---|---|
| PCNA | NM_001291925.1 | F:GTGATTCCACCACCATGTC R:TGAGACGAGTCCATGCTCG | 145 |
| BAX | XM_013998624.2 | F:GCCGAAATGTTTGCTGACG R:CAGCCGATCTCGAAGGAAG | 156 |
| BCL-2 | XM_021077298.1 | F:GAGCGTAGACAAGGAGATGC R:TCCGACTGAAGAGCGAAC | 239 |
| TGF-β1 | AF461808 | F:AAAGCGGCAACCAAATCTATGA R:GCTGAGGTAGCGCCAGGAAT | 206 |
| Smad3 | NM_214137 | F:TGGTGCCACGCCACACAGAG R:TCGGGGAGAGGTTTGGAGAA | 213 |
| GAPDH | NM_001206359.1 | F:ATGGTGAAGGTCGGAGTGAA R:CGTGGGTGGAATCATACTGG | 154 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Yang, L.; Shao, M.; Wang, Y.; Yang, W.; Huang, L.; Zhou, X.; Jiang, S.; Yang, Z. Effects of Zearalenone Exposure on the TGF-β1/Smad3 Signaling Pathway and the Expression of Proliferation or Apoptosis Related Genes of Post-Weaning Gilts. Toxins 2018, 10, 49. https://doi.org/10.3390/toxins10020049
Zhou M, Yang L, Shao M, Wang Y, Yang W, Huang L, Zhou X, Jiang S, Yang Z. Effects of Zearalenone Exposure on the TGF-β1/Smad3 Signaling Pathway and the Expression of Proliferation or Apoptosis Related Genes of Post-Weaning Gilts. Toxins. 2018; 10(2):49. https://doi.org/10.3390/toxins10020049
Chicago/Turabian StyleZhou, Min, Lijie Yang, Minghui Shao, Yuxi Wang, Weiren Yang, Libo Huang, Xuemei Zhou, Shuzhen Jiang, and Zaibin Yang. 2018. "Effects of Zearalenone Exposure on the TGF-β1/Smad3 Signaling Pathway and the Expression of Proliferation or Apoptosis Related Genes of Post-Weaning Gilts" Toxins 10, no. 2: 49. https://doi.org/10.3390/toxins10020049