Saccharomyces cerevisiae Boulardii Reduces the Deoxynivalenol-Induced Alteration of the Intestinal Transcriptome

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
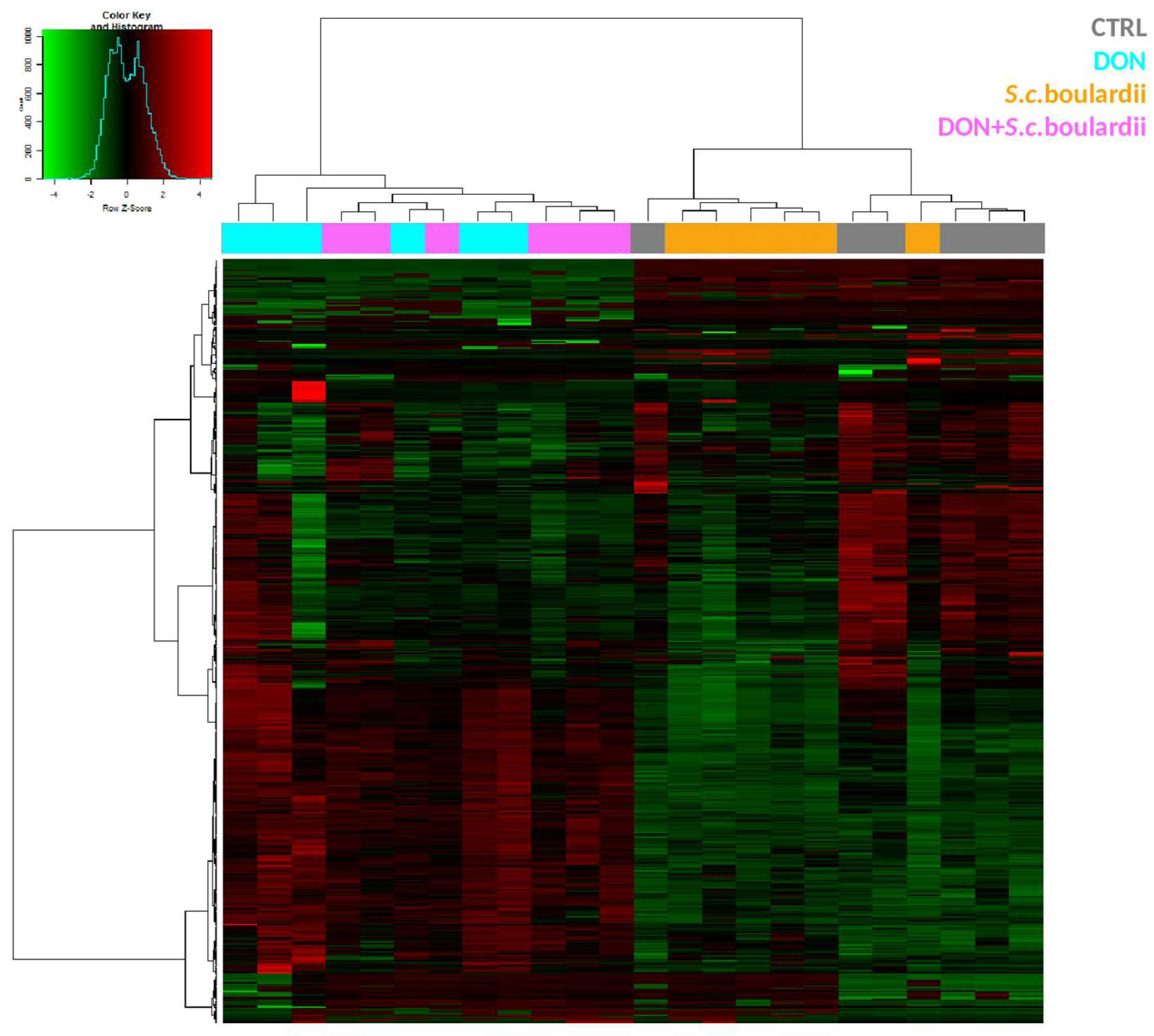
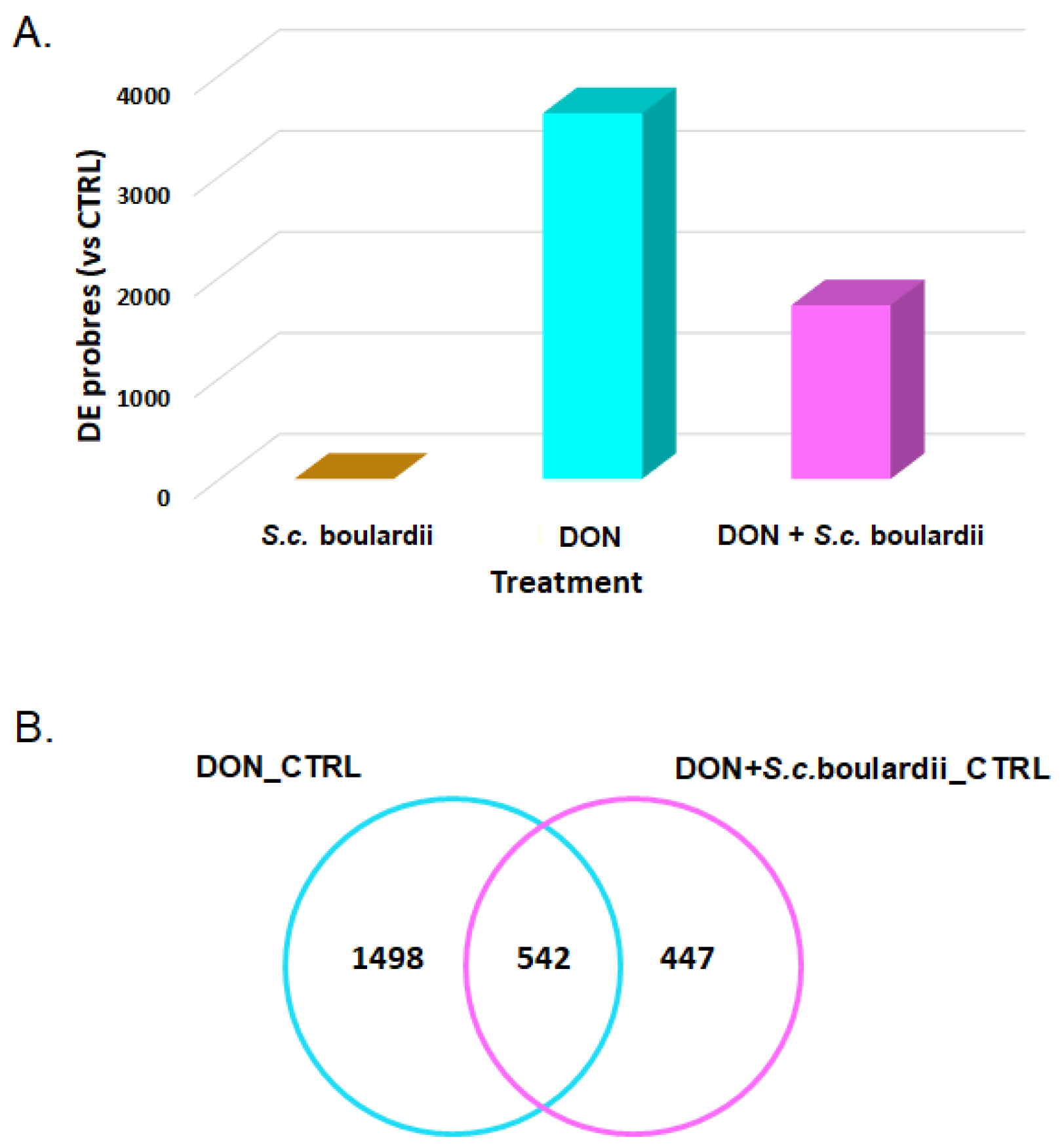
2.1. S. cerevisiae boulardii Reduces the Transcriptomic Impact of DON on the Intestine
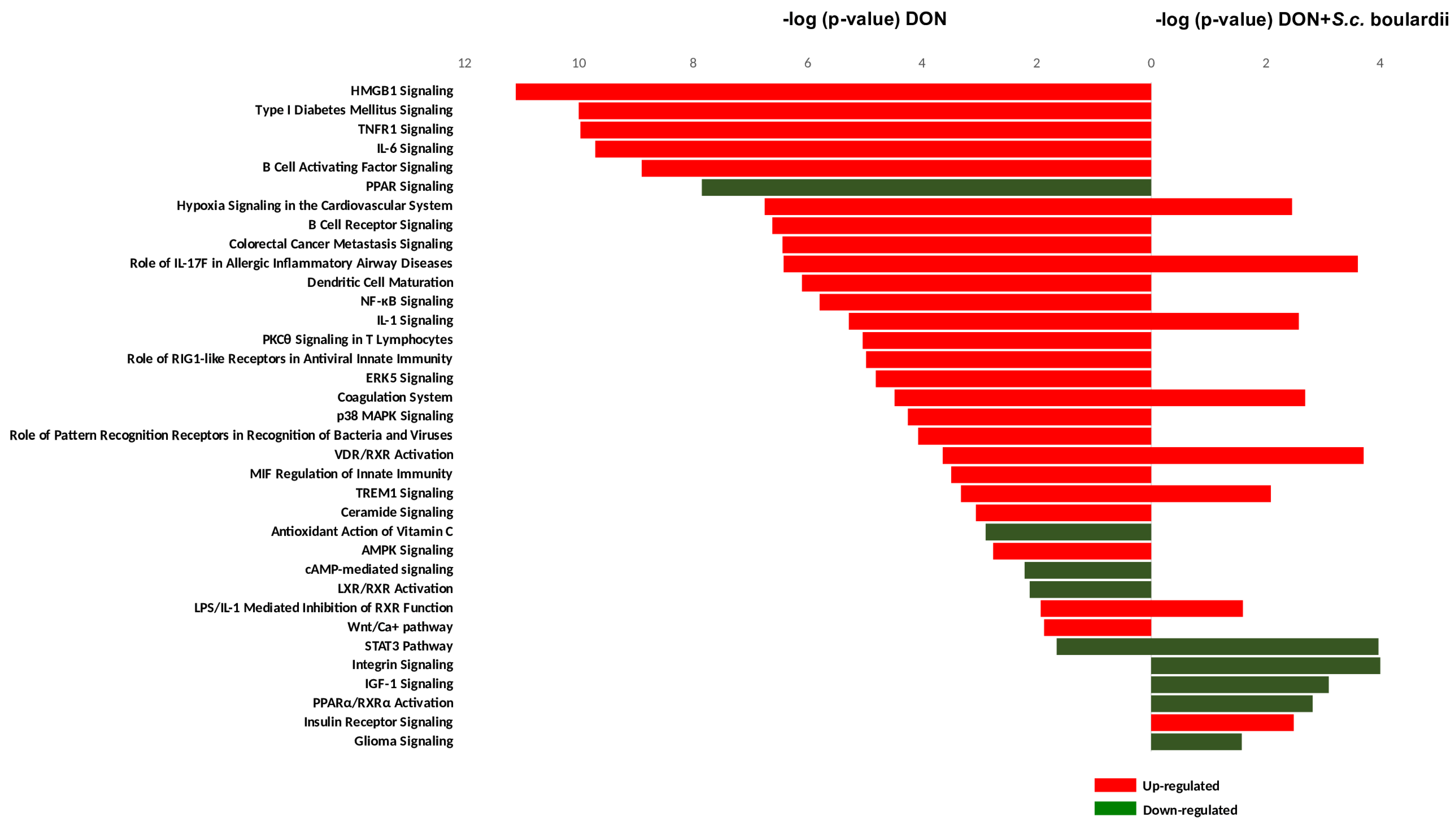
2.2. Functional Analysis of the Intestinal Transcriptome Modulation by DON and/or S. cerevisiae boulardii
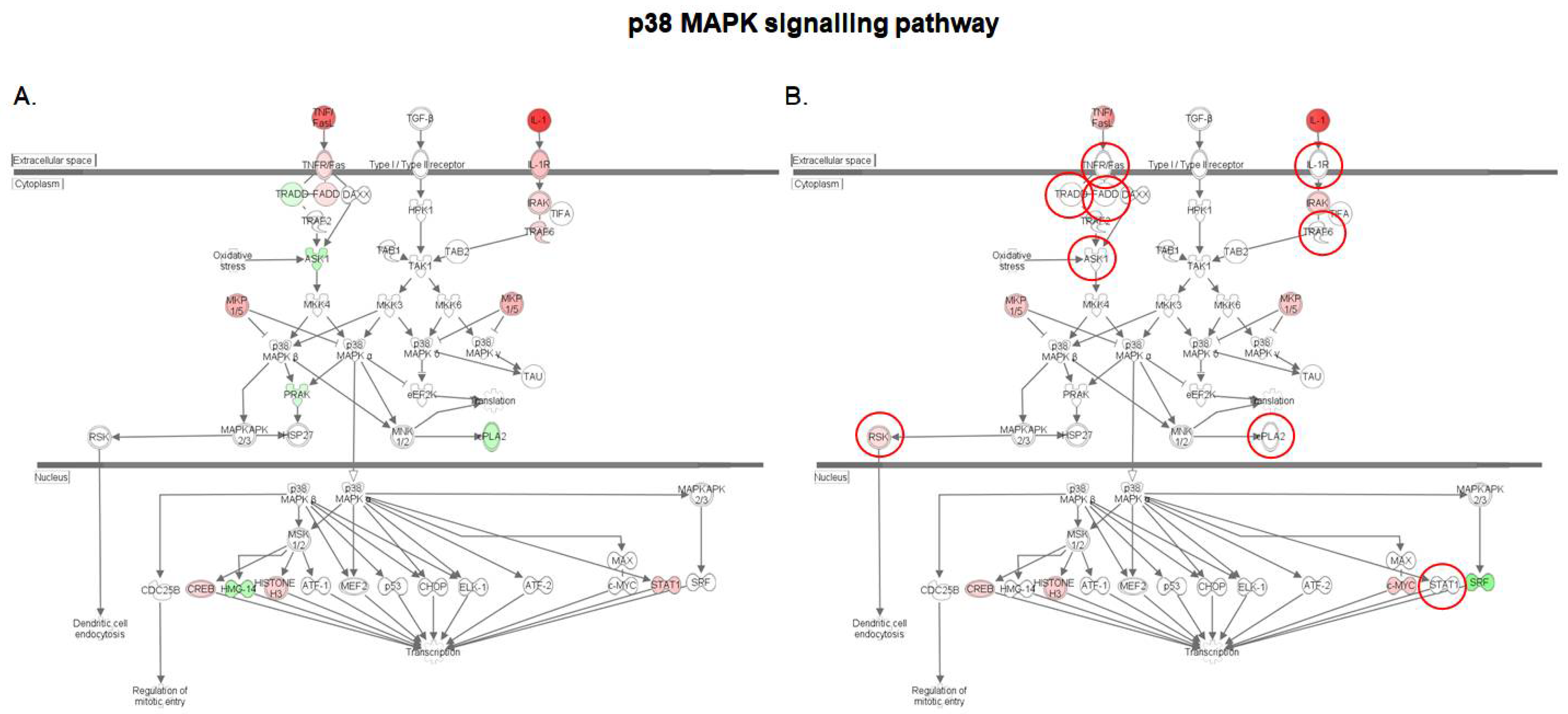
2.3. S. cerevisiae boulardii Reduces the Pro-Inflammatory Effects of DON
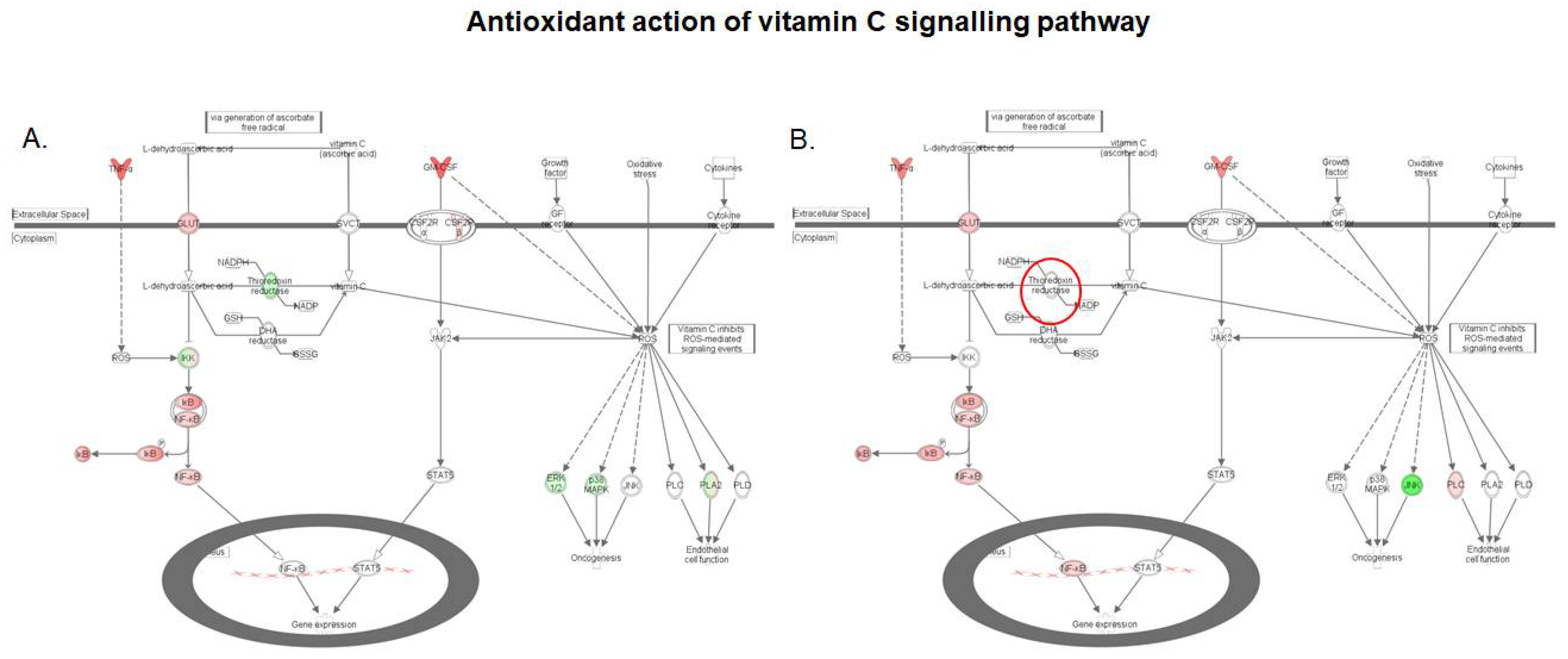
2.4. S. cerevisiae boulardii Reverses the Effect of DON on the Antioxidant Action of Vitamin C
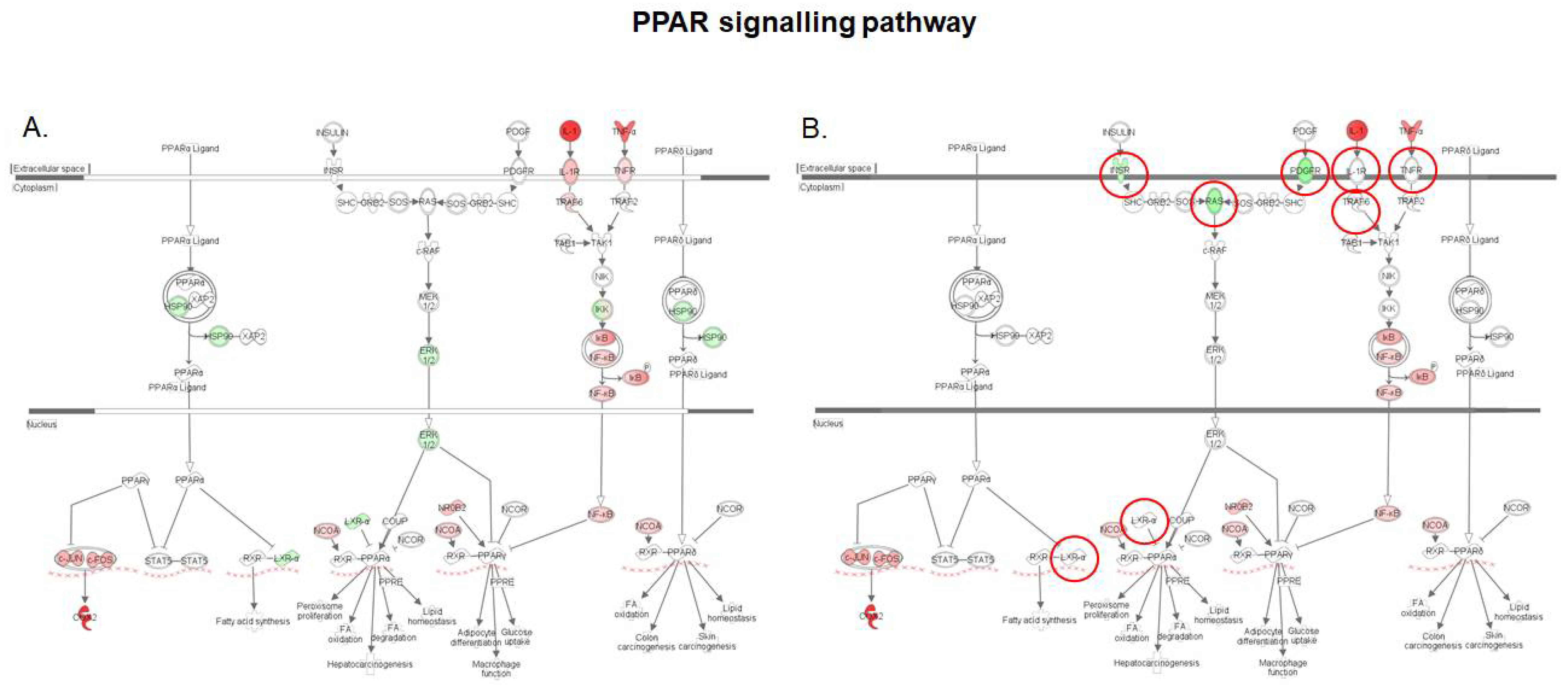
2.5. S. cerevisiae boulardii Restores the Lipid Metabolism Altered by DON
3. Conclusions
4. Materials and Methods
4.1. Toxin
4.2. Yeast Strain and Culture
4.3. Culture of Jejunum Explants
4.4. RNA Extraction
4.5. Microarray Processing and Functional Analysis of Expressed Genes
4.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Robert, H.; Payros, D.; Pinton, P.; Theodorou, V.; Mercier-Bonin, M.; Oswald, I.P. Impact of mycotoxins on the intestine: Are mucus and microbiota new targets? J. Toxicol. Environ. Health Part B Crit. Rev. 2017, 20, 249–275. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Oswald, I.P. Effect of deoxynivalenol and other type B trichothecenes on the intestine: A review. Toxins 2014, 6, 1615–1643. [Google Scholar] [CrossRef] [PubMed]
- Cano, P.M.; Seeboth, J.; Meurens, F.; Cognie, J.; Abrami, R.; Oswald, I.P.; Guzylack-Piriou, L. Deoxynivalenol as a new factor in the persistence of intestinal inflammatory diseases: An emerging hypothesis through possible modulation of TH17-mediated response. PLoS ONE 2013, 8, e53647. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol. Toxins 2013, 5, 784–820. [Google Scholar] [CrossRef] [PubMed]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Mimoun, S.; Murate, L.S.; Loiseau, N.; Lippi, Y.; Bracarense, A.P.; Liaubet, L.; Schatzmayr, G.; Berthiller, F.; Moll, W.D.; et al. Intestinal toxicity of the masked mycotoxin deoxynivalenol-3-beta-d-glucoside. Arch. Toxicol. 2016, 90, 2037–2046. [Google Scholar] [CrossRef] [PubMed]
- Alassane-Kpembi, I.; Kolf-Clauw, M.; Gauthier, T.; Abrami, R.; Abiola, F.A.; Oswald, I.P.; Puel, O. New insights into mycotoxin mixtures: The toxicity of low doses of type B trichothecenes on intestinal epithelial cells is synergistic. Toxicol. Appl. Pharmacol. 2013, 272, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Alassane-Kpembi, I.; Puel, O.; Pinton, P.; Cossalter, A.M.; Chou, T.C.; Oswald, I.P. Co-exposure to low doses of the food contaminants deoxynivalenol and nivalenol has a synergistic inflammatory effect on intestinal explants. Arch. Toxicol. 2017, 91, 2677–2687. [Google Scholar] [CrossRef] [PubMed]
- Alassane-Kpembi, I.; Puel, O.; Oswald, I.P. Toxicological interactions between the mycotoxins deoxynivalenol, nivalenol and their acetylated derivatives in intestinal epithelial cells. Arch. Toxicol. 2015, 89, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Garcia, G.R.; Payros, D.; Pinton, P.; Dogi, C.A.; Laffitte, J.; Neves, M.; Gonzalez Pereyra, M.L.; Cavaglieri, L.R.; Oswald, I.P. Intestinal toxicity of deoxynivalenol is limited by lactobacillus rhamnosus RC007 in pig jejunum explants. Arch. Toxicol. 2018, 92, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Hemarajata, P.; Versalovic, J. Effects of probiotics on gut microbiota: Mechanisms of intestinal immunomodulation and neuromodulation. Ther. Adv. Gastroenter. 2013, 6, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Pontier-Bres, R.; Rampal, P.; Peyron, J.F.; Munro, P.; Lemichez, E.; Czerucka, D. The Saccharomyces boulardii CNCM I-745 strain shows protective effects against the B. Anthracis lt toxin. Toxins 2015, 7, 4455–4467. [Google Scholar] [CrossRef] [PubMed]
- Trevisi, P.; Latorre, R.; Priori, D.; Luise, D.; Archetti, I.; Mazzoni, M.; D’Inca, R.; Bosi, P. Effect of feed supplementation with live yeast on the intestinal transcriptome profile of weaning pigs orally challenged with Escherichia coli f4. Animal 2017, 11, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Weaver, A.C.; See, M.T.; Hansen, J.A.; Kim, Y.B.; De Souza, A.L.; Middleton, T.F.; Kim, S.W. The use of feed additives to reduce the effects of aflatoxin and deoxynivalenol on pig growth, organ health and immune status during chronic exposure. Toxins 2013, 5, 1261–1281. [Google Scholar] [CrossRef] [PubMed]
- Sougioultzis, S.; Simeonidis, S.; Bhaskar, K.R.; Chen, X.; Anton, P.M.; Keates, S.; Pothoulakis, C.; Kelly, C.P. Saccharomyces boulardii produces a soluble anti-inflammatory factor that inhibits NF-kappab-mediated IL-8 gene expression. Biochem. Biophys. Res. Commun. 2006, 343, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Wang, K.; Zhou, S.N.; Wang, X.D.; Wu, J.E. Protective effect of Saccharomyces boulardii on deoxynivalenol-induced injury of porcine macrophage via attenuating p38 MAPK signal pathway. Appl. Biochem. Biotechnol. 2017, 182, 411–427. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Nougayrede, J.P.; Del Rio, J.C.; Moreno, C.; Marin, D.E.; Ferrier, L.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharmacol. 2009, 237, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutsen, H.K.; Alexander, J.; Barregard, L.; Bignami, M.; Bruschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15. [Google Scholar] [CrossRef]
- Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.D.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; Gropp, J.; Kolar, B.; et al. Safety and efficacy of Levucell® SB (Saccharomyces cerevisiae CNCM I-1079) as a feed additive for weaned piglets and sows. EFSA J. 2016, 14, e04478. [Google Scholar]
- Alassane-Kpembi, I.; Gerez, J.R.; Cossalter, A.M.; Neves, M.; Laffitte, J.; Naylies, C.; Lippi, Y.; Kolf-Clauw, M.; Bracarense, A.P.L.; Pinton, P.; et al. Intestinal toxicity of the type B trichothecene mycotoxin fusarenon-x: Whole transcriptome profiling reveals new signaling pathways. Sci. Rep. 2017, 7, 7530. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Mimoun, S.; Murate, L.S.; Loiseau, N.; Lippi, Y.; Bracarense, A.P.; Schatzmayr, G.; He, J.W.; Zhou, T.; Moll, W.D.; et al. Microbial biotransformation of DON: Molecular basis for reduced toxicity. Sci. Rep. 2016, 6, 29105. [Google Scholar] [CrossRef] [PubMed]
- Ricote, M.; Valledor, A.F.; Glass, C.K. Decoding transcriptional programs regulated by ppars and LXRS in the macrophage: Effects on lipid homeostasis, inflammation, and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Klune, J.R.; Dhupar, R.; Cardinal, J.; Billiar, T.R.; Tsung, A. Hmgb1: Endogenous danger signaling. Mol. Med. 2008, 14, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Csh Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Pestka, J.J. Comparative induction of 28S ribosomal RNA cleavage by ricin and the trichothecenes deoxynivalenol and T-2 toxin in the macrophage. Toxicol. Sci. 2008, 105, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Cuenda, A.; Rousseau, S. P38 map-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Noguchi, T.; Takeda, K.; Ichijo, H. Pathophysiological roles of ASK1-map kinase signaling pathways. J. Biochem. Mol. Biol. 2007, 40, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Yamamoto, K.; Miki, Y.; Murase, R.; Sato, H.; Taketomi, Y. The roles of the secreted phospholipase A2 gene family in immunology. Adv. Immunol. 2016, 132, 91–134. [Google Scholar] [PubMed]
- Majoros, A.; Platanitis, E.; Kernbauer-Holzl, E.; Rosebrock, F.; Muller, M.; Decker, T. Canonical and non-canonical aspects of JAK-STAT signaling: Lessons from interferons for cytokine responses. Front. Immunol. 2017, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.C. The noncanonical NF-kappab pathway. Immunol. Rev. 2012, 246, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Osselaere, A.; Santos, R.; Hautekiet, V.; De Backer, P.; Chiers, K.; Ducatelle, R.; Croubels, S. Deoxynivalenol impairs hepatic and intestinal gene expression of selected oxidative stress, tight junction and inflammation proteins in broiler chickens, but addition of an adsorbing agent shifts the effects to the distal parts of the small intestine. PLoS ONE 2013, 8, e69014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Dwivedi, P.D.; Pandey, H.P.; Das, M. Role of oxidative stress in deoxynivalenol induced toxicity. Food Chem. Toxicol. 2014, 72, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.H.; Wang, X.; Yang, W.; Nussler, A.K.; Xiong, L.Y.; Kuca, K.; Dohnal, V.; Zhang, X.J.; Yuan, Z.H. Oxidative stress-mediated cytotoxicity and metabolism of T-2 toxin and deoxynivalenol in animals and humans: An update. Arch. Toxicol. 2014, 88, 1309–1326. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, A.F.; Atroshi, F.; Ahotupa, M.; Sankari, S.; Elovaara, E. Protective effect of antioxidants against free radical-mediated lipid peroxidation induced by don or T-2 toxin. Zentralb Vet. Reihe A 1994, 41, 81–90. [Google Scholar] [CrossRef]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. Rsc Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef]
- Chinetti, G.; Lestavel, S.; Bocher, V.; Remaley, A.T.; Neve, B.; Torra, I.P.; Teissier, E.; Minnich, A.; Jaye, M.; Duverger, N.; et al. PPAR-alpha and PPAR-gamma activators induce cholesterol removal from human macrophage foam cells through stimulation of the abca1 pathway. Nat. Med. 2001, 7, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Burns, K.A.; Vanden Heuvel, J.P. Modulation of ppar activity via phosphorylation. Biochim. Biophys. Acta 2007, 1771, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.; Reginato, M.J.; Shao, D.; Lazar, M.A.; Chatterjee, V.K. Transcriptional activation by peroxisome proliferator-activated receptor gamma is inhibited by phosphorylation at a consensus mitogen-activated protein kinase site. J. Biol. Chem. 1997, 272, 5128–5132. [Google Scholar] [CrossRef] [PubMed]
- Hu, E.; Kim, J.B.; Sarraf, P.; Spiegelman, B.M. Inhibition of adipogenesis through map kinase-mediated phosphorylation of ppargamma. Science 1996, 274, 2100–2103. [Google Scholar] [CrossRef] [PubMed]
- Canonici, A.; Pellegrino, E.; Siret, C.; Terciolo, C.; Czerucka, D.; Bastonero, S.; Marvaldi, J.; Lombardo, D.; Rigot, V.; Andre, F. Saccharomyces boulardii improves intestinal epithelial cell restitution by inhibiting alphaVSeta5 integrin activation state. PLoS ONE 2012, 7, e45047. [Google Scholar] [CrossRef] [PubMed]
- Czerucka, D.; Piche, T.; Rampal, P. Review article: Yeast as probiotics—Saccharomyces boulardii. Aliment. Pharmacol. Ther. 2007, 26, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Kolf-Clauw, M.; Castellote, J.; Joly, B.; Bourges-Abella, N.; Raymond-Letron, I.; Pinton, P.; Oswald, I.P. Development of a pig jejunal explant culture for studying the gastrointestinal toxicity of the mycotoxin deoxynivalenol: Histopathological analysis. Toxicol. In Vitro 2009, 23, 1580–1584. [Google Scholar] [CrossRef] [PubMed]
- Lucioli, J.; Pinton, P.; Callu, P.; Laffitte, J.; Grosjean, F.; Kolf-Clauw, M.; Oswald, I.P.; Bracarense, A.P. The food contaminant deoxynivalenol activates the mitogen activated protein kinases in the intestine: Interest of ex vivo models as an alternative to in vivo experiments. Toxicon 2013, 66, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Devriendt, B.; Gallois, M.; Verdonck, F.; Wache, Y.; Bimczok, D.; Oswald, I.P.; Goddeeris, B.M.; Cox, E. The food contaminant fumonisin B1 reduces the maturation of porcine CD11R1+ intestinal antigen presenting cells and antigen-specific immune responses, leading to a prolonged intestinal ETEC infection. Vet. Res. 2009, 40, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Graziani, F.; Pujol, A.; Nicoletti, C.; Paris, O.; Ernouf, P.; Di Pasquale, E.; Perrier, J.; Oswald, I.P.; Maresca, M. Deoxynivalenol inhibits the expression by goblet cells of intestinal mucins through a PKR and MAP kinase dependent repression of the resistin-like molecule beta. Mol. Nutr. Food Res. 2015, 59, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Bracarense, A.P.; Schwartz, H.E.; Trumel, C.; Cossalter, A.M.; Schatzmayr, G.; Kolf-Clauw, M.; Moll, W.D.; Oswald, I.P. The low intestinal and hepatic toxicity of hydrolyzed fumonisin B1 correlates with its inability to alter the metabolism of sphingolipids. Biochem. Pharmacol. 2012, 83, 1465–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DON | DON + S. cerevisiae boulardii | |||||
|---|---|---|---|---|---|---|
| Gene Symbol | Fold Change * | Adjusted p Value | Gene Symbol | Fold Change * | Adjusted p Value | |
| Up–regulated | IL-1β | 11.37 | 6.27 × 10−13 | IL-1β | 9.56 | 6.01 × 10−12 |
| IL-22 | 5.62 | 3.24 × 10−8 | IL-22 | 5.56 | 7.73 × 10−8 | |
| NOR-1 | 5.09 | 1.07 × 10−9 | PTGS2 | 3.96 | 1.32 × 10−6 | |
| PTGS2 | 5.01 | 3.63 × 10−8 | IL-1α | 3.96 | 2.90 × 10−8 | |
| CXCL2 | 4.65 | 0.00045897 | CCL20 | 3.96 | 3.85 × 10−7 | |
| IL-1α | 4.17 | 5.78 × 10−9 | NOR-1 | 3.88 | 7.73 × 10−8 | |
| CCL20 | 4.17 | 8.96 × 10−8 | CXCL2 | 3.22 | 0.01419023 | |
| CCR7 | 3.36 | 1.36 × 10−7 | IL8 | 2.95 | 0.0004445 | |
| HAMP | 3.34 | 2.46 × 10−5 | ARX 10G | 2.87 | 7.48 × 10−7 | |
| PLK2 | 3.28 | 1.02 × 10−11 | PRDM1 | 2.86 | 8.79 × 10−12 | |
| AREG | 3.26 | 7.49 × 10−8 | FOSL1 | 2.82 | 0.00026329 | |
| PRDM1 | 3.17 | 6.27 × 10−13 | GADD45G | 2.7 | 9.54 × 10−6 | |
| FOSL1 | 3.11 | 4.71 × 10−5 | CCR7 | 2.63 | 1.35 × 10−5 | |
| CSF2 | 2.93 | 4.60 × 10−5 | PLK2 | 2.6 | 2.32 × 10−9 | |
| ADAMTS1 | 2.92 | 9.33 × 10−10 | GADD45A | 2.6 | 2.12 × 10−6 | |
| CRSP-2 | 2.9 | 1.31 × 10−5 | PRDM1 | 2.59 | 9.84 × 10−10 | |
| CD274 | 2.74 | 3.47 × 10−7 | ATF3 | 2.57 | 4.04 × 10−6 | |
| RRAD | 2.73 | 0.00150172 | PLK2 | 2.5 | 3.89 × 10−9 | |
| IFRD1 | 2.66 | 1.47 × 10−8 | RND1 | 2.48 | 3.07 × 10−6 | |
| GADD45G | 2.64 | 7.57 × 10−6 | HAMP | 1.31 | 0.001954 | |
| Down–regulated | CTBS | −3.43 | 0.0323898 | ACTN2 | −4.83 | 0.03632277 |
| C1QTNF3 | −2.38 | 0.02819327 | ACTA1 | −4.28 | 0.03241036 | |
| PDLIM3 | −2.2 | 0.04545171 | TPM2 | −2.88 | 0.04875964 | |
| BTC | −2 | 0.04718046 | TPM1 | −2.81 | 0.04897366 | |
| TMEFF2 | −1.97 | 0.02364126 | MYL9 | −2.66 | 0.04636262 | |
| HAND1 | −1.89 | 0.04906228 | LIMS2 | −2.57 | 0.04897366 | |
| SMPX | −1.82 | 0.01890697 | MYLK | −2.53 | 0.04955867 | |
| BPI | −1.76 | 0.03066332 | C1QTNF3 | −2.49 | 0.03403947 | |
| LYZ | −1.72 | 0.02546107 | SPOCK3 | −2.41 | 0.02910998 | |
| LECT1 | −1.7 | 0.04314249 | BTC | −2.26 | 0.04788811 | |
| ASPN | −1.67 | 0.04519455 | TPM1 | −2.23 | 0.04179965 | |
| OGN | −1.66 | 0.03041459 | HAND1 | −2.2 | 0.02447994 | |
| RYR2 | −1.64 | 0.03593429 | FLNA | −2.1 | 0.03855459 | |
| DSTN | −1.64 | 0.00936255 | TMX 10FF2 | −2.03 | 0.02907563 | |
| FHL1C | −1.63 | 0.0438132 | CPXM2 | −2.02 | 0.02989774 | |
| CXCL12 | −1.63 | 0.01364805 | DNAJA4 | −1.94 | 0.01272008 | |
| CYP4F2 | −1.59 | 0.04673074 | PRUNX 102 | −1.86 | 0.0392422 | |
| ITGB1BP2 | −1.59 | 0.0354242 | SMARCD3 | −1.83 | 0.04490504 | |
| ACTB | −1.59 | 0.02928137 | FHL1C | −1.81 | 0.02399939 | |
| MATN2 | −1.59 | 0.0065206 | ATP2B4 | −1.81 | 0.04252773 | |
| DON | DON + S. cerevisiae boulardii | |||||||
|---|---|---|---|---|---|---|---|---|
| Gene Symbol | FC Microarray * | Adjusted p Value | FC qRT-PCR ** | p Value | FC Microarray * | Adjusted p Value | FC qRT-PCR | p Value |
| IL-1β | 11.37 | 6.27 × 10−13 | 6.08 | 9 × 10−4 | 9.56 | 6.01 × 10−12 | 4.96 | 0.0089 |
| IL-22 | 5.62 | 3.24 × 10−8 | 5.88 | 0.0791 | 5.56 | 7.73 × 10−8 | 3.65 | 0.0964 |
| IL8 | 2.22 | 0.0081 | 3.13 | 1.8 × 10−4 | 2.95 | 4.45 × 10−4 | 2.91 | 0.0025 |
| IL17A | 2.28 | 2.4 × 10−4 | 7.62 | 0.0022 | 1.81 | 0.0025 | 1.75 | 0.018 |
| TNF-α | 2.07 | 1.82 × 10−7 | 2.99 | 0.0085 | 1.71 | 6.14 × 10−5 | 1.61 | 0.01 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alassane-Kpembi, I.; Pinton, P.; Hupé, J.-F.; Neves, M.; Lippi, Y.; Combes, S.; Castex, M.; Oswald, I.P. Saccharomyces cerevisiae Boulardii Reduces the Deoxynivalenol-Induced Alteration of the Intestinal Transcriptome. Toxins 2018, 10, 199. https://doi.org/10.3390/toxins10050199
Alassane-Kpembi I, Pinton P, Hupé J-F, Neves M, Lippi Y, Combes S, Castex M, Oswald IP. Saccharomyces cerevisiae Boulardii Reduces the Deoxynivalenol-Induced Alteration of the Intestinal Transcriptome. Toxins. 2018; 10(5):199. https://doi.org/10.3390/toxins10050199
Chicago/Turabian StyleAlassane-Kpembi, Imourana, Philippe Pinton, Jean-François Hupé, Manon Neves, Yannick Lippi, Sylvie Combes, Mathieu Castex, and Isabelle P. Oswald. 2018. "Saccharomyces cerevisiae Boulardii Reduces the Deoxynivalenol-Induced Alteration of the Intestinal Transcriptome" Toxins 10, no. 5: 199. https://doi.org/10.3390/toxins10050199