1. Introduction
Fusarium mycotoxins are secondary metabolites produced by
Fusarium species of fungi, which can contaminate animal feed ingredients. Deoxynivalenol (DON) and zearalenone (ZEN) are considered to be the most important
Fusarium mycotoxins due to their high toxicity [
1]. A global survey indicated that
Fusarium mycotoxins DON and ZEN, respectively, contaminated 55% and 36% of feed and feed ingredients in the period 2004–2011 [
2], which results in a large economic loss in the livestock industry. Among farm animals, pigs are particularly sensitive to
Fusarium mycotoxins [
3]. Ingestion of DON is usually associated with feed refusal, low weight gains, low feed efficiency, diarrhea, and organ damage in pigs [
3,
4]. DON also significantly alters functions of the intestinal tract, including decreasing villus surface area and altering the permeability of the intestinal tract in weaning piglets [
5]. Treatment with ZEN leads to hyperestrogenism, precocious puberty, and reproductive disorders [
1,
3]. In many cases, DON and ZEN often simultaneously found in naturally
Fusarium fungi contaminated feed. Therefore, the co-occurrence of mycotoxins may make their toxicity effects on pigs more complicated. The liver is one of the main organs for metabolism of toxic substances, and liver damage might occur when pigs consume overdoses of mycotoxins [
6]. An increasing number of studies have demonstrated that oxidative stress plays a major role in the toxicity of
Fusarium mycotoxins [
7,
8].
Hydrogen gas (H
2), historically thought an inert and nonfunctional gas, is defined as a selective antioxidant in recent years. Molecular hydrogen can selectively neutralize toxic free radicals such as hydroxyl radicals and nitrite anion without disturbing physiological reactive oxygen species (ROS) [
9]. Unlike common antioxidant reagents, hydrogen has a unique advantage that it can penetrate biomembranes and diffuse into cellular components [
10]. As a result, hydrogen has been applied in many animal liver damage models through its anti-inflammatory, anti-apoptotic, and anti-oxidant properties [
11]. Inhalation of H
2 gas suppresses hepatic ischemia/reperfusion injury by reducing oxidative stress in mice [
12]. Hydrogen-rich water (HRW) has also shown remarkable protective effects against obstructive jaundice [
13] and chronic EtOH-induced [
14] liver damage, possibly by activating antioxidant enzymes.
Supplementation of hydrogen-producing prebiotics is also a feasible way of providing functional hydrogen to animals and humans [
11]. Lactulose, a nonabsorbable synthetic disaccharide consisting of fructose and galactose, has been proposed as an indirect antioxidant by mobilizing intestinal hydrogen production [
15]. Lactulose possesses neuroprotective [
16] effects against cerebral ischemia/reperfusion injury and anti-inflammatory [
17,
18] effects on dextran sodium sulfate-induced colon inflammatory, which could be ascribed to the hydrogen production produced during microbial fermentation of lactulose. Lactulose administration accelerates liver regeneration in a rat hepatectomy model by inducing hydrogen [
19]. Previous strategies to mitigate the toxic effects of
Fusarium mycotoxins in animal production involved mycotoxins adsorbent and mycotoxins-degrading enzymes and micro-organisms.
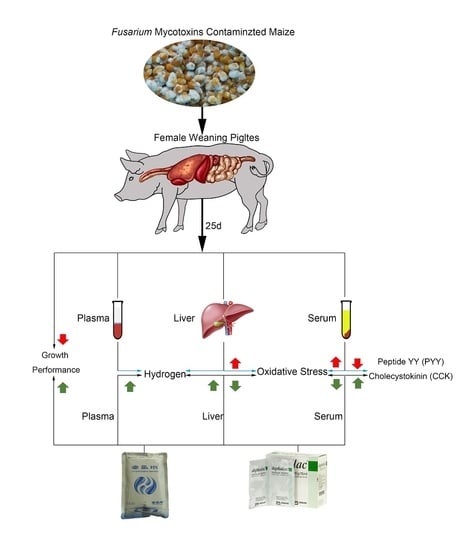
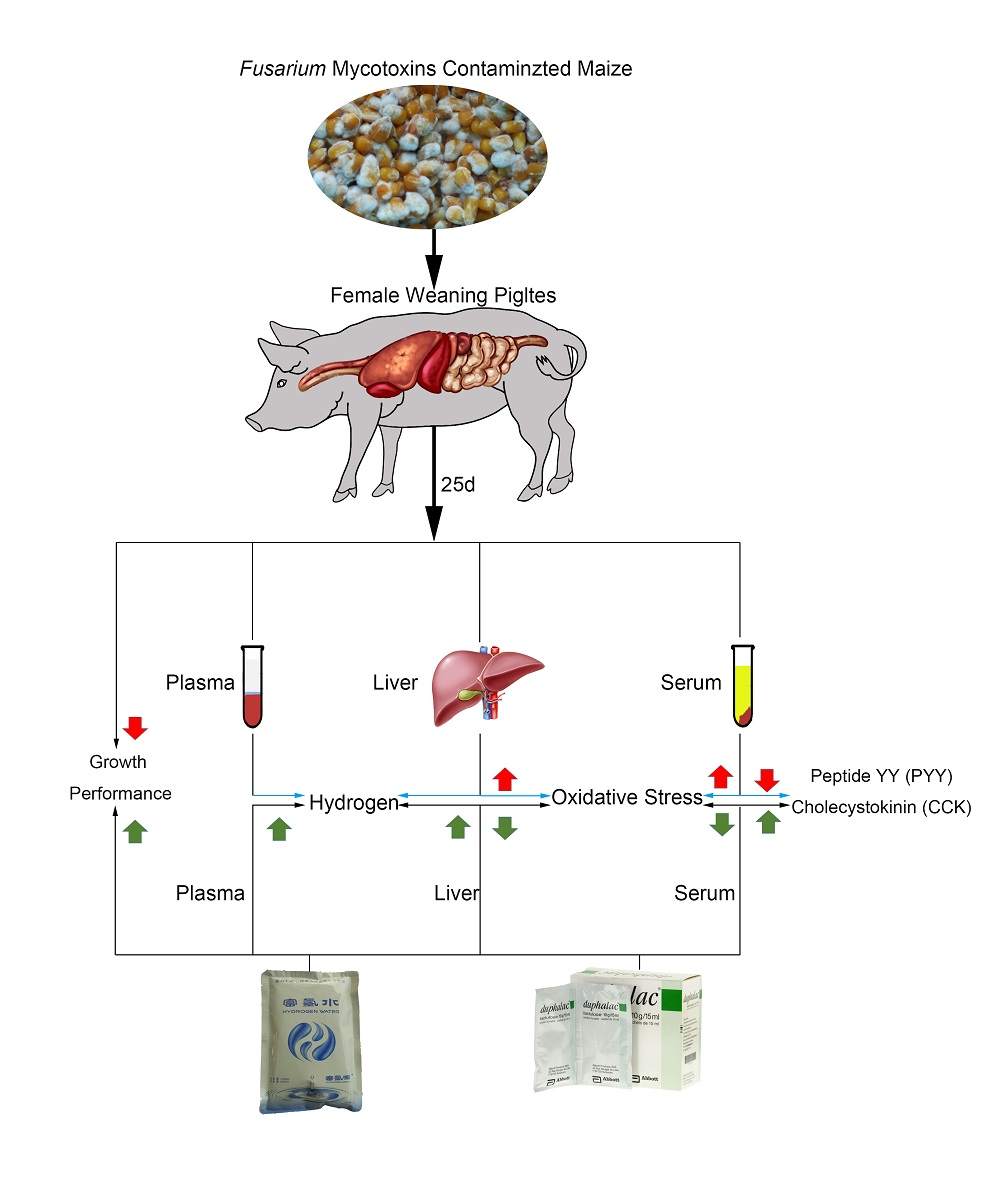
We hypothesize that both HRW and lactulose may protect weaning piglets against growth depression and liver damage caused by Fusarium mycotoxins contaminated diet by increasing hydrogen levels and activating the antioxidant capacity in the body.
5. Materials and Methods
5.1. Preparation of Fusarium Contaminated Maize
Fusarium graminearum strain 2021 was kindly supplied by Prof. Ming-Guo Zhou, College of Plant Protection, Nanjing Agricultural University, Nanjing, China. The strain was cultured as described in a previous study [
48]. Briefly, the strain was first cultivated on potato sucrose agar (PSA) at 25 °C for 7 days. The hyphae of fungi were obtained, inoculated in mung bean broth (MBB), and cultured with shaking (200 rpm/min at 25 °C) for 7 days for conidia production. Before inoculation, maize was soaked with water for 48 h and autoclaved. The conidia were inoculated to cool autoclaved maize at a concentration of 1 × 10
6 conidia/kg. The
Fusarium contaminated maize was incubated in plastic storage boxes for 30 days (temperature 15–25 °C and humidity 50–85%). The cool autoclaved maize not inoculated with conidia was used as control maize. Finally, control and maize with
Fusarium mold were dried in an oven at 70 °C for 24 h, respectively.
5.2. Experimental Diets and Mycotoxins Analysis
Fusarium mycotoxins-contaminated maize and uncontaminated control maize were used at 50% at the expense of normal maize for the manufacturing of two experimental diets, respectively. The experimental diets were formulated according to the recommendations of the nutrient requirement of swine by National Research Council [
49] and based on a previous study [
41], with a minor modification to the vitamin and mineral premix. No antibiotic, hormone and preservatives were added to the diets.
Table A1 shows the ingredients of the experimental diets used in this study.
Fusarium mycotoxins levels in the two experimental diets were analyzed as previously described [
50]. Briefly, a 10 g diet sample was extracted with 25 mL of acetonitrile:water (80:20,
v/v) at 180 rpm for 30 min. After centrifugation at 3000 rpm for 10 min, 0.5 mL supernatant was diluted with 1.5 mL of acetonitrile:water (80:20,
v/v) and filtered through a nylon filter (13 mm in diameter and 0.22 μm pore size). Then, the filtered solution was analyzed by a high-pressure liquid chromatography/electrospray ionization–tandem mass spectrometry (LC–MS/MS) system consisting of an Agilent 1200 HPLC (Agilent Technology, Shanghai, China), an Agilent 6410B triple-quadrupole mass spectrometer (Agilent Technology, Shanghai, China), and an Agilent MassHunter Workstation running Qualitative Analysis version B.01.03 software (Agilent Technology, Shanghai, China, 2001) for data acquisition and analysis. No significant differences were found in nutrient composition between the NC and MC experimental diets (
Table A1). Levels of the main
Fusarium mycotoxins in 2 experimental diets are summarized in
Table A2. The NC diet contained 221.10 μg/kg DON, 12.12 μg/kg 3-acetyl DON, 32.95 μg/kg 15-acetyl DON, and 266.26 μg/kg total DON, respectively. While the MC diet contained 825.46 μg/kg DON, 212.79 μg/kg 3-acetyl DON, 59.45 μg/kg 15-acetyl DON, 1097.99 μg/kg total DON, and 501.56 μg/kg ZEN, which were all significantly higher (
p < 0.05) than in the NC diet (
Table A2).
5.3. Animals
All experiments and protocols used in this study were approved by the Nanjing Agricultural University Institutional Animal Care and Use Committee (Certification No.: SYXK (Su) 2011-0036, 11 August 2015). A total of 24 clinically healthy female weaning piglets (Landrace × large × white) from 6 littles (4 pigs/little) were individually housed in pens (1.2 by 2.0 m) with 1 feeder and 1 nipple drinker. The piglets had ab libitum access to feed and water.
5.4. Experiment Design and Sample Collection
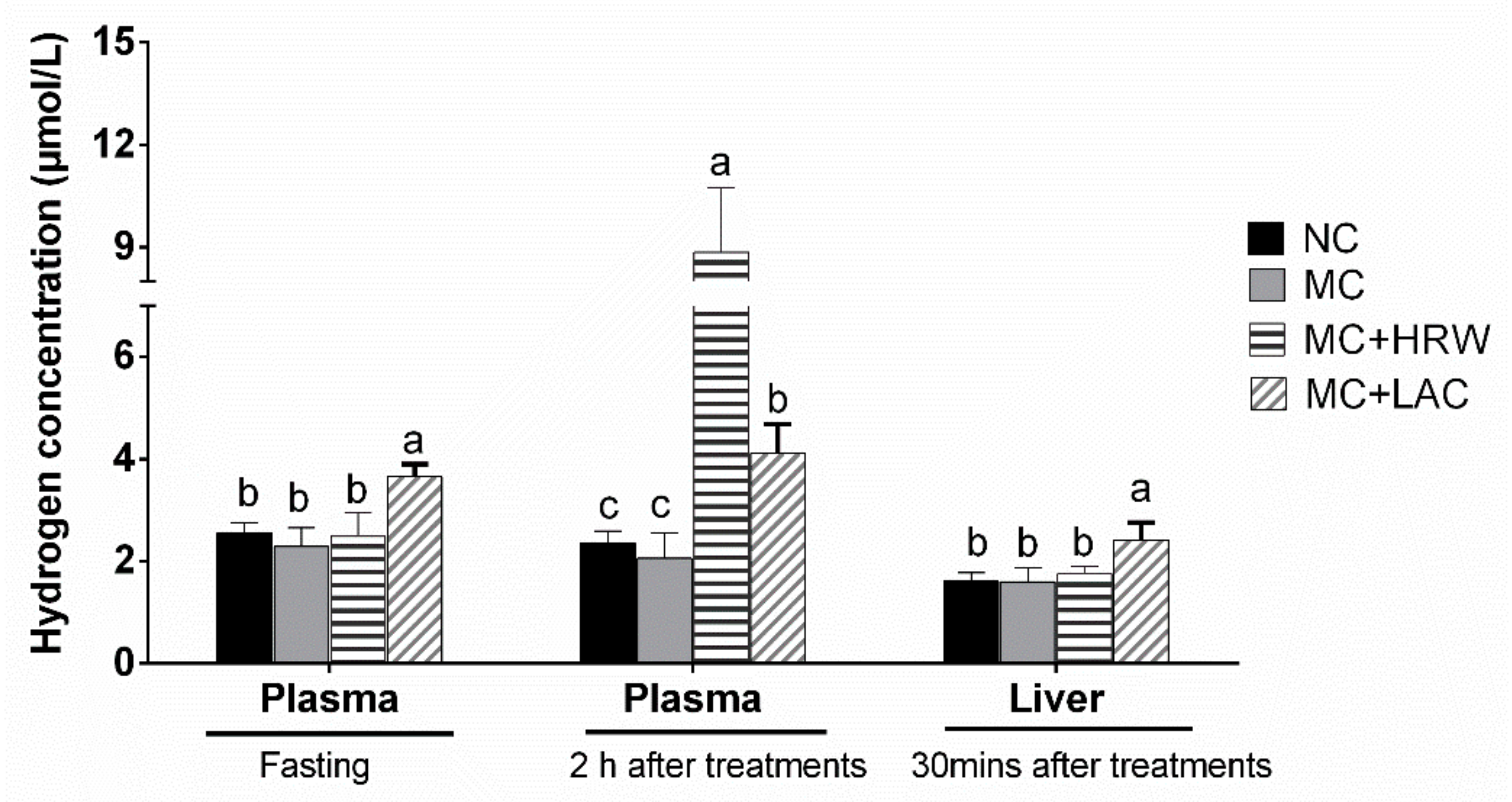
The piglets from the same little were randomly assigned to receive 1 of 4 treatments (total 6 pigs/treatment): uncontaminated basal diet (negative control (NC)), mycotoxin-contaminated (MC) diet, MC diet + lactulose (MC + LAC) and MC diet + hydrogen-rich water (MC + HRW). After a 6-day adaption period, each treatment fed their respective diet for 25 days. Piglets received two times/day (10:00 and 14:00, respectively) and 10 mL/kg BW/time for their respective treatments. Piglets in NC and MC groups were orally administrated with hydrogen-free water (HFW), MC + HRW group received hydrogen-rich water (HRW), and MC + LAC group received 500 mg/kg BW of LAC (dissolved in 10 mL of HFW). The hydrogen-rich water (Beijing Hydrovita Biotechnology Company, Beijing, China) was kept in 300 mL aluminum pouches and administered to piglets within 15 min after opened. The H2 concentration was 0.6 mM as measured by an H2 sensor (Unisense, Aarhus, Denmark). Each animal’s daily feed intake and weekly body weight were recorded. Amounts of HFW, LAC and HRW were dependent on the body weight and updated weekly.
On Day 21, plasma samples at fasting and two hours after different administrations were collected to detect the hydrogen levels. One piglet was removed from the MC, MC + LAC and MC + HRW groups fed with Fusarium mycotoxins contaminated maize due to the poor health condition. Therefore, five independent replicates from each group were used in this study. At the end of the experiment, 30 min after administration of different treatments, piglets were euthanized by an intramuscular injection of sodium pentobarbital (40 mg/kg BW). The serum and liver were sampled.
5.5. Serum Hormones and Antioxidant Assay
Serum hormone levels of ghrelin, PYY, and CKK were measured by commercial ELISA kit according to their instructions (Fangcheng Beijing Technology Co. Ltd., Beijing, China). Liver samples were homogenized as previously described [
51]. Oxidant and antioxidant parameters in the serum and liver supernatants were analyzed using assay kit according to the manufacturer’s instructions (Nanjing Jiancheng, Nanjing, China).
5.6. Hydrogen Gas Measurement in Plasma and Liver Samples
Hydrogen levels in serum samples were analyzed using a hydrogen sensor (Unisense, Aarhus, Denmark). Samples of liver were prepared as previously described [
52]. Briefly, piglets were euthanized with sodium pentobarbital and placed in supine position. An incision was made in the abdomen. Hydrogen microelectrode (diameter, 50 μm) was inserted into the liver at a depth of 500 μm.
5.7. Statistical Analysis
All statistical analyses were performed using the SPSS software (Version 18.0, SPSS Inc., Chicago, IL, USA, 2009). The differences among treatments were evaluated using one-way ANOVA followed by Tukey–Kramer test. Data were considered to be statistically significant if p < 0.05.



{kind=link}
{kind=link}
