1. Introduction
Staphylococcus aureus continues to be one of the most prevalent pathogens causing intramammary infections (IMI) in dairy cows. It’s a worldwide pathogen recognized as a cause of subclinical infections, resulting in increased somatic cell count (SCC), but may also cause clinical mastitis. Staphylococcal mastitis is a major problem in dairy industry, affecting animal health and causing economic losses of up to €300 per cow per year, due to the reduced milk quality and production [
1,
2]. The main reservoir of
S. aureus seems to be the infected quarter, and transmission usually occurs from cow to cow during milking.
Successful infection depends on virulence factors produced by
S. aureus. A wide spectrum of secreted and cell surface-associated virulence factors can be expressed to promote adhesion to the host extracellular matrix components, damage host cells, and fight the immune system [
3]. At least 25 different toxins (such as enterotoxins SEA to SEQ, toxic shock syndrome toxin-1 TSST-1, exfoliative toxins Eta, Etb), 15 microbial surface components recognizing adhesive matrix molecules, which are important for adhesion to tissues (such as clumping factor A
clfA, intercellular adhesion genes
icaA and
icaD), 20 immune evasion molecules (such as protein A, coagulase, haemolysins and leucocidins, factors associated with suppressing innate immunity) and several other
S. aureus virulence factors are known. Some virulence factors are expressed by genes that are located on mobile genetic elements called pathogenicity islands (i.e., TSST and some enterotoxins) or lysogenic bacteriophages (i.e., Panton-Valentine Leucocidin, PVL) and others such as the staphylococcal complement inhibitor,
scn, the chemotaxis inhibitory protein,
chp, and staphylokinase,
sak, are integrated in the bacterial chromosome [
4]. Furthermore,
S. aureus can also acquire the staphylococcal cassette chromosome SCC
mec, giving rise to methicillin-resistant
S. aureus (MRSA) [
5]. In fact, the expression of the
mecA or
mecC gene in
S. aureus confers resistance to most of β-lactams, drugs which are frequently used for treatment of mastitis [
6].
The determination of the origin of the S. aureus isolates involved in the aetiology of bovine mastitis is highly relevant from the epidemiological point of view. In such a context, the precise characterization of this pathogen provides monitoring of the bacterial strains dissemination among animal populations.
Over the past two decades, a wide range of phenotyping and genotyping methods have been used or developed for
S. aureus including, but not limited to, ribotyping, RAPD-typing, PFGE, MLST, spa-typing, RS-PCR, coagulase gene RFLP, MLVA, micro-arrays and whole genome comparisons [
7,
8,
9,
10,
11]. Many molecular epidemiological studies have been based on the use of selected targets in the genome, giving rise to banding patterns based on restriction- or primer binding sites, or to allelic profiles for housekeeping or virulence genes [
12]. Such studies continue to be useful diagnostic tools when the aim is to understand pathogen sources and transmission mechanisms. Moreover, among the genotyping methods, the RS-PCR, based on amplifying the 16S-23S rRNA intergenic spacer region by PCR (ribosomal spacer-PCR), showed to be accurate, rapid and inexpensive with a discriminatory power like the other more-recognized genotyping methods [
13].
The aim of this study was to genotype by RS-PCR and compare the molecular-epidemiologic profiles of a large world collection of S. aureus isolates to deepen the knowledge on the circulating genetic lineages among the cow population with mastitis. The isolates were investigated for three genes related to host adhesion and invasion (clfA, clumping factor; cna, collagen-binding protein; fmtB, cell wall-associated protein), 22 genes that have the potential to interfere with host defence mechanisms (tsst, toxic shock syndrome toxin-1; scn, staphylococcal complement inhibitor; chp, chemotaxis inhibitory protein; sak, staphylokinase; enterotoxins from sea to sel; exfoliative toxins eta, etb and leucocidins lukE, lukE-lukD, lukM, lukSF-PV), and the gene encoding the acquisition of methicillin resistance (mecA).
3. Discussion
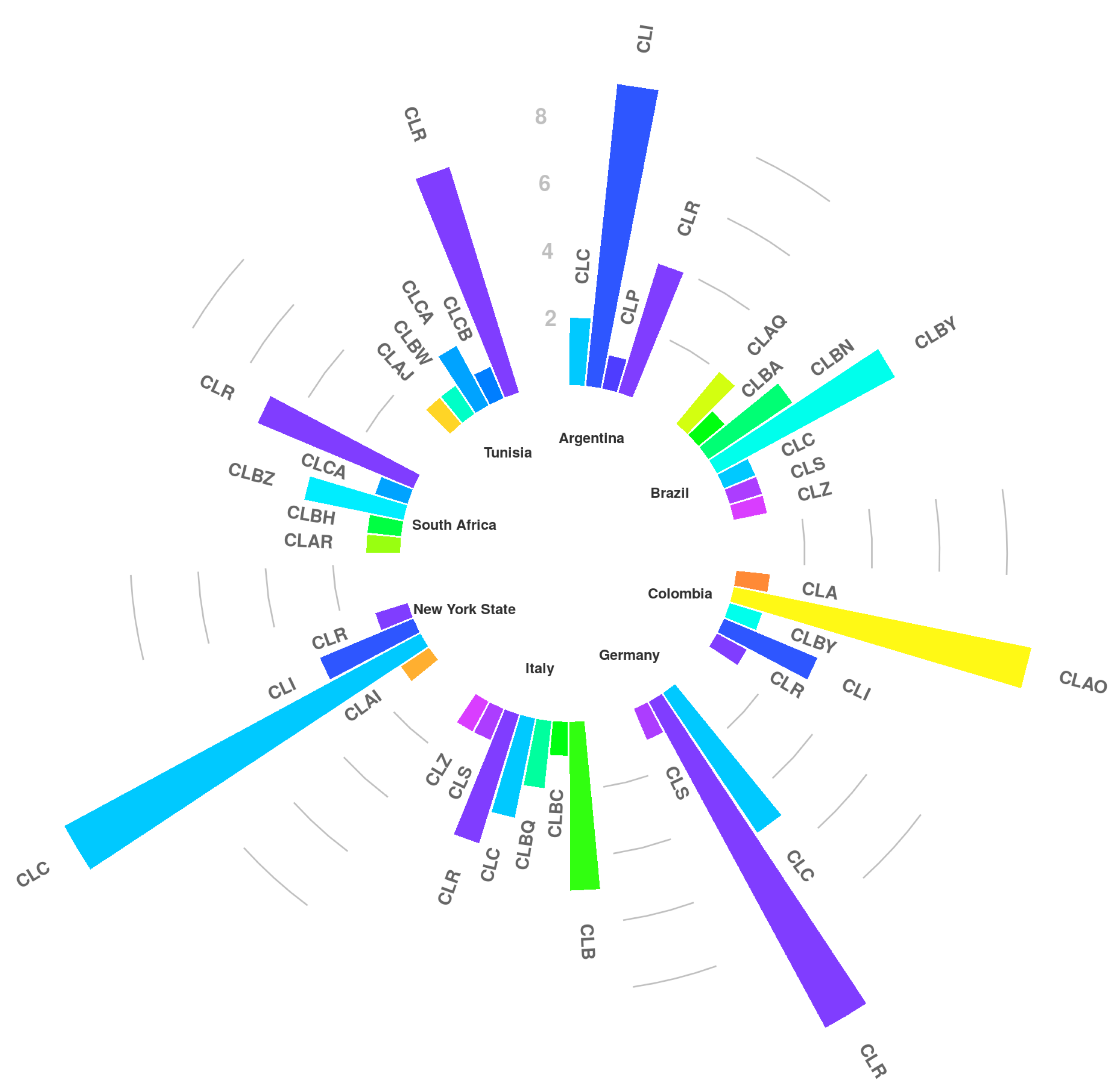
Pathogenic factors of S. aureus enable this bacterium to attach, colonize, invade and infect the host tissue. In this study, S. aureus isolates, collected from eight different countries, were investigated using RS-PCR genotyping and PCR analysis for the carriage of different virulence factors to examine the epidemiology of this microorganism.
The samples were obtained from collections of the collaborators, allowing a first overview about the presence of the various staphylococcal subtypes among countries. Three new genotypes were observed for South Africa whereas new variants were found in Argentina, Brazil, Colombia, Italy and New York State. As previously described [
14], GTB was observed only in Europe (Italy) while CLR and CLC clusters were observed throughout America, Europe and Africa; particularly CLR, which forms a large cluster containing 13 variants, was detected in each country involved, except for Brazil. It is quite well demonstrated [
14] that
S. aureus CLC and CLR clusters are “dairy cattle specific” whose spreading process must have been started a long time ago, with the spreading of breeding cattle from Europe to the other countries. On the contrary, GTB derives from a more recent bovine adaptation due to a new human-to-cow host jump [
13]. Certainly, further studies will be necessary to explain the different geographic distribution especially for the minor genotypes.
As previously described [
15],
S. aureus isolates harbouring genes coding for clumping factor (
clfA), a cell wall-associated protein (
fmtB), and collagen-binding protein (
cna) have a greater capability to adhere to extracellular matrix proteins, essential for colonization and the establishment of infections. Our results indicated that, except for the American isolates with a lower presence of
fmtB and
clfA genes, in the other seven countries these genes were widely present in the circulating isolates particularly in Brazilian and Tunisian ones. The presence of these genes, necessary for host invasion, could improve the persistence of the microorganism in the host, ensuring the probability of survival in the population.
And more, according to previous studies [
11,
13,
15], except for Brazil, Germany and USA, the remaining countries showed isolates encoding at least 2 virulence factors out of staphylococcal complement inhibitor (
scn), chemotaxis inhibitory protein of
S. aureus (
chp) and staphylokinase (
sak). These virulence factors show activity prevalently against the human innate immune system but their presence among isolates recovered in herds with high prevalence of
S. aureus mastitis suggests their involvement also in bovine mammary gland immune response [
16], and should be further studied, especially in Colombia and Tunisia where this gene cluster is quite common [
17]. In a previous study [
4], human strains were grouped in 7 immune evasion cluster (IEC) types, depending on the presence of 2 out of the 3 genes, in association or not with
sea or
sep. Unlike Colombian, Italian, South African strains and Tunisian isolates, the Argentinian ones carried only one gene,
sak, showing a clear distance from human strains. Among the isolates from the other countries, uniquely the Tunisian strains testing positive for IEC, did not harbor
sea.
Superantigens, especially enterotoxins, have been suggested to play a role in the development of mastitis, for instance by creating an attractive environment for colonization [
18] since they are more often identified in
S. aureus isolated from cows with mastitis than in isolates from healthy cows or from the environment [
19]. As a result, enterotoxins support the pathogenesis of
S. aureus compromising mammary gland immune response and susceptibility to antibiotics resulting in the onset of many diseases [
20]. In this study,
sea and
sei were the main enterotoxin genes present in all countries except for Tunisia (prevalence between 50% and 90%). While
seh gene had a frequency higher than 90% in Argentinian, Brazilian, Colombian and South African isolates,
sej and
sel genes were carried only by Italian and Tunisian isolates, respectively. Among the 120 isolates analyzed, only 17 (14%) were not enterotoxigenic (1 from Argentina, 1 from Colombia, 2 isolates from Germany, 3 from Italy, 1 from New York State, 1 from South Africa, and 8 from Tunisia). The remaining 103 isolates (86%) harboured a combination of at least 2 up to 5 enterotoxins with the linkages between
sea,
sed,
seg and
seh confirming their predominance in cows, as previously described [
21,
22,
23,
24]. The absence of the enterotoxin genes
seb and
see in our isolates was in accordance with previous results [
15,
22,
25,
26].
Here, among all the isolates we did not find the presence of
etb exfoliative gene and only one isolate from South Africa was positive for
eta gene. These results agree with previous studies conducted in different countries [
27,
28,
29], showing that
S. aureus isolates from animals with mastitis were rarely positive for exfoliative toxins. On the contrary, in Europe, Kot and coworkers reported a 14.5% of
S. aureus harbouring the
eta gene from bovine mastitis [
30]. In our study, the presence of
tsst gene was more relevant, being carried by 37% of Argentinian, 23% of German, 16% of Tunisian and 6% of Italian isolates. All these isolates were also positive at least for a combination of
sec and
sel, or
sec,
seg, and
sei or
sec,
seg and
sej or
sec,
seg and
sel genes which are located on the same bovine staphylococcal pathogenicity island (SaPIbov), confirming a positive correlation between
sec, sei or
sej and
tsst, as previously reported [
31].
Panton-Valentine leucocidin, encoded by 2 co-transcribed genes located on a prophage, causes leukocyte destruction and tissue necrosis [
32]. The presence of PVL-encoding genes in
S. aureus is reported to be associated with increased disease severity [
33]. In the present study, the presence of PVL gene was lower than 20% in South Africa and New York State, higher than 80% in Argentina, Colombia and Brazil, while in Germany, Italy and Tunisia none of the
S. aureus isolates carried the gene. For European countries, previously published results were in accordance with this study [
34,
35]. Additionally, genes encoding the bicomponent leucotoxin
lukE-lukD were observed in all isolates, and, except for South Africa with only 2 isolates, most of the other isolates harboured
lukM, a gene encoding one operon like the one of PVL. The high rates of
lukE-lukD and
lukM found in this study agree with other reports [
34,
35,
36]. Additionally, only 2 isolates, one from Germany and one from Italy were positive for
mecA, confirming the low diffusion of MRSA among bovine mastitis isolates [
37,
38]; interestingly, they are both GTS, in accordance with previous results [
13].
4. Conclusions
Knowledge about the epidemiology of
S. aureus genotypes in dairy species and herds might help to formulate strategies for reducing the infection spreading and for focused treatments. In our work we found that CLR and CLC clusters and some virulence factors related to host invasion, such as
fmtB,
cna,
clfA or immune defense impairment such as leukocidin genes, were the most frequent ones. These genes combination could be related to the
S. aureus ability to colonize the host. Further,
fmtB gene has been shown to be related to the resistance of
S. aureus to β-lactam antibiotics [
10]. Therefore, due to the prevalence of these genes worldwide, it might be useful screening them in
S. aureus isolates to help predicting clinical outcomes and specially to identify harmful strains. Meanwhile, our work demonstrated also that each country had a specific genotypic pattern and in some countries the isolates harboured some virulence factors, such as PVL-encoding genes, with high prevalence, recommending a close surveillance of
S. aureus isolates in the animals of these countries to avoid the wide spreading of these genes. Finally, it is notable that most of the isolates worldwide were negative for
mecA, confirming the evidence of the low diffusion of MRSA among bovine mastitis isolates, as previously described [
37,
38].
In conclusion, this study confirms the wide variety of S. aureus genotypes found in dairy cattle worldwide and that genetic differences are related to geographical origin of the isolates, suggesting that considering the region of interest and the strain virulence might help to formulate strategies directed to reduce the infection spreading and to set up control measures according to pathogen and host features. Therefore, based the characterization of the circulating strain, the farmer would be able to decide to segregate positive cows applying hygienic milking procedures and a suitable milking order, or even to cull the infected animals.


 ,
, 


{kind=link}