The Antioxidant Gallic Acid Inhibits Aflatoxin Formation in Aspergillus flavus by Modulating Transcription Factors FarB and CreA



Abstract
:1. Introduction
2. Results
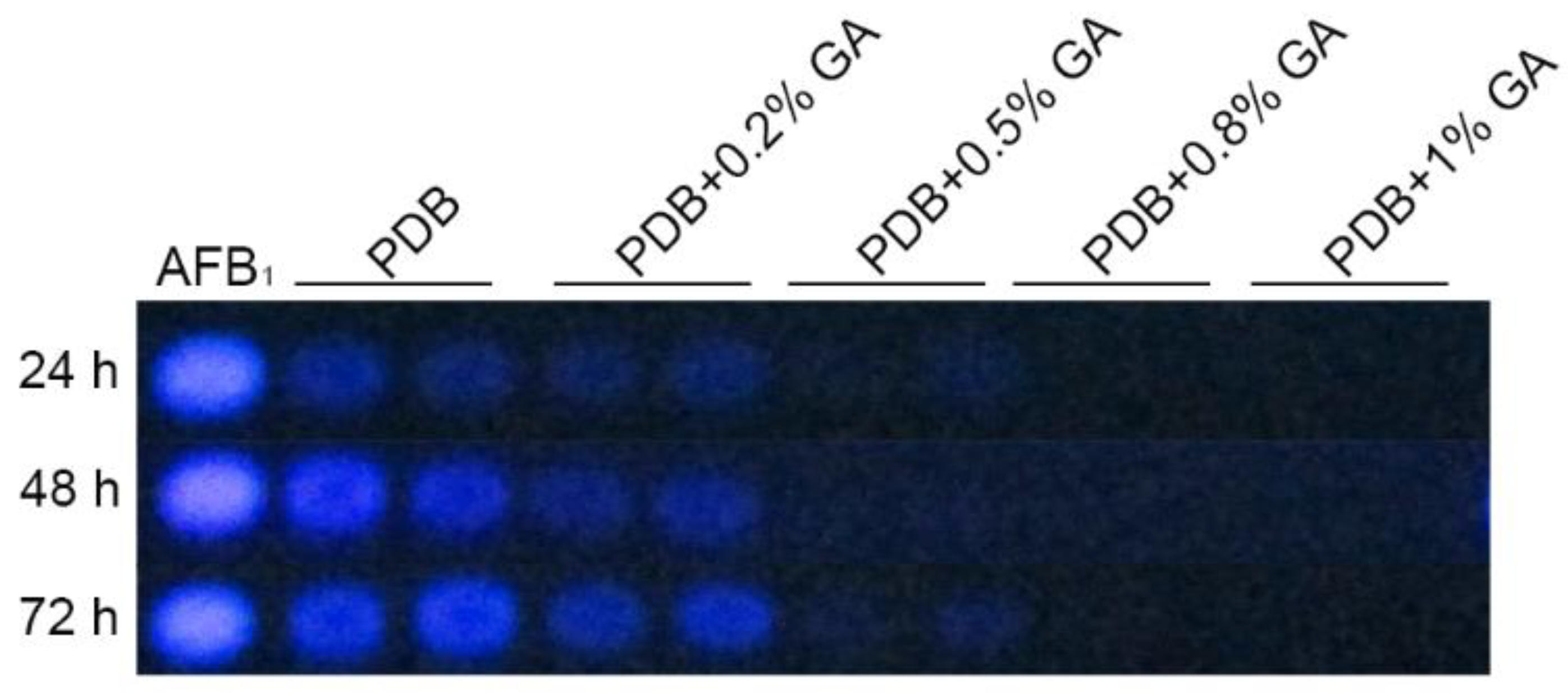
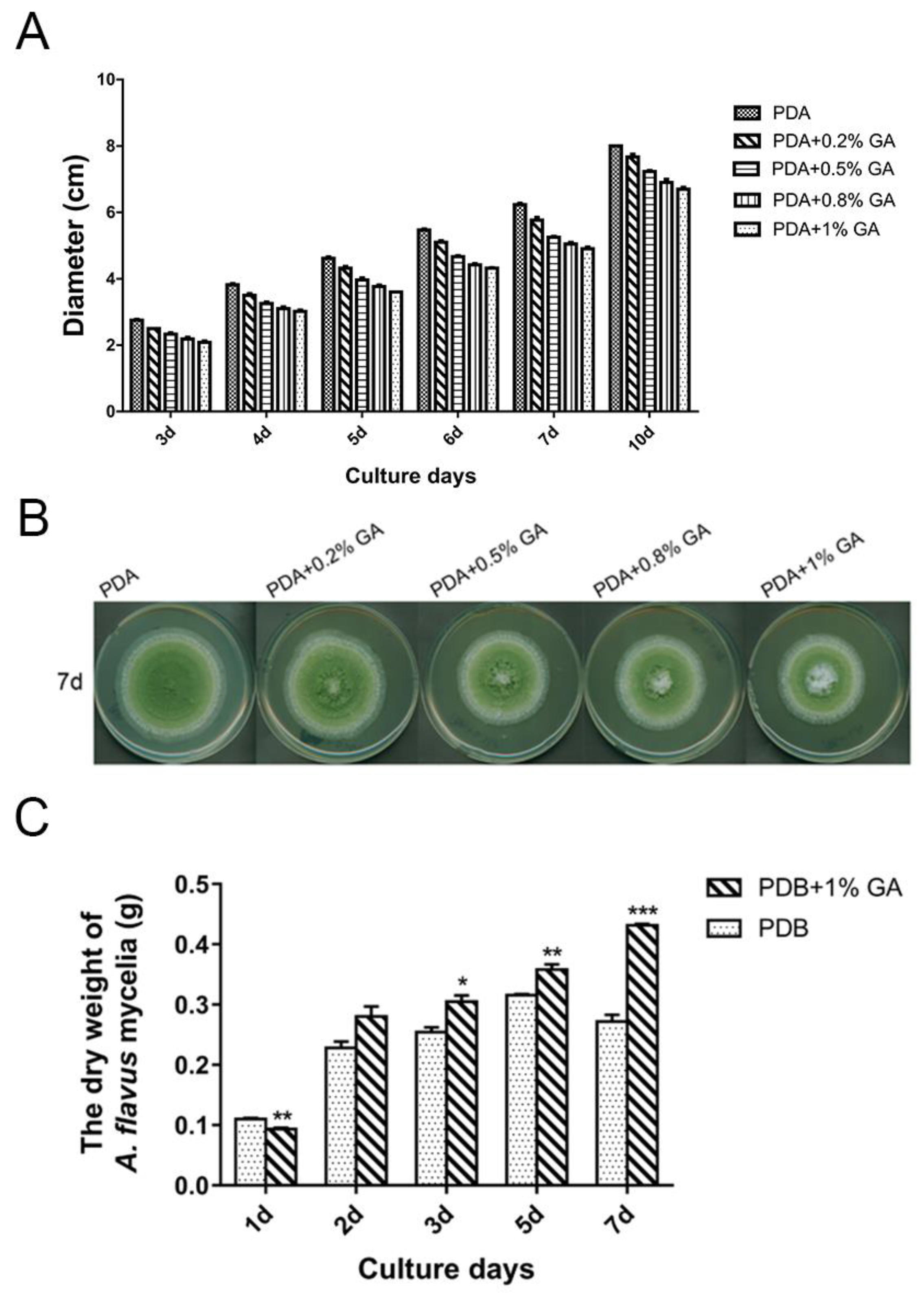
2.1. Effect of GA on Aflatoxin Biosynthesis and Growth in A. flavus
2.2. Transcriptomic Profiles of A. flavus Responding to GA
2.3. Effect of GA on the Expression of Genes in A. flavus Secondary Metabolism Clusters
2.4. Expression Changes of the Developmental Regulatory Genes and the Primary Metabolic Genes in A. flavus in Response to GA
2.5. Effect of GA on Oxidative Stress Genes and the Antioxidant Enzymatic Activities
3. Discussion
4. Materials and Methods
4.1. Fungal Strain and Culture Conditions
4.2. Aflatoxin Analysis and Measuring the A. flavus Colony Diameters and Mycelia Weights
4.3. RNA Extraction and cDNA Library Preparation for RNA-Seq Analysis
4.4. Data Analysis and Normalizing the Expression Levels of Genes
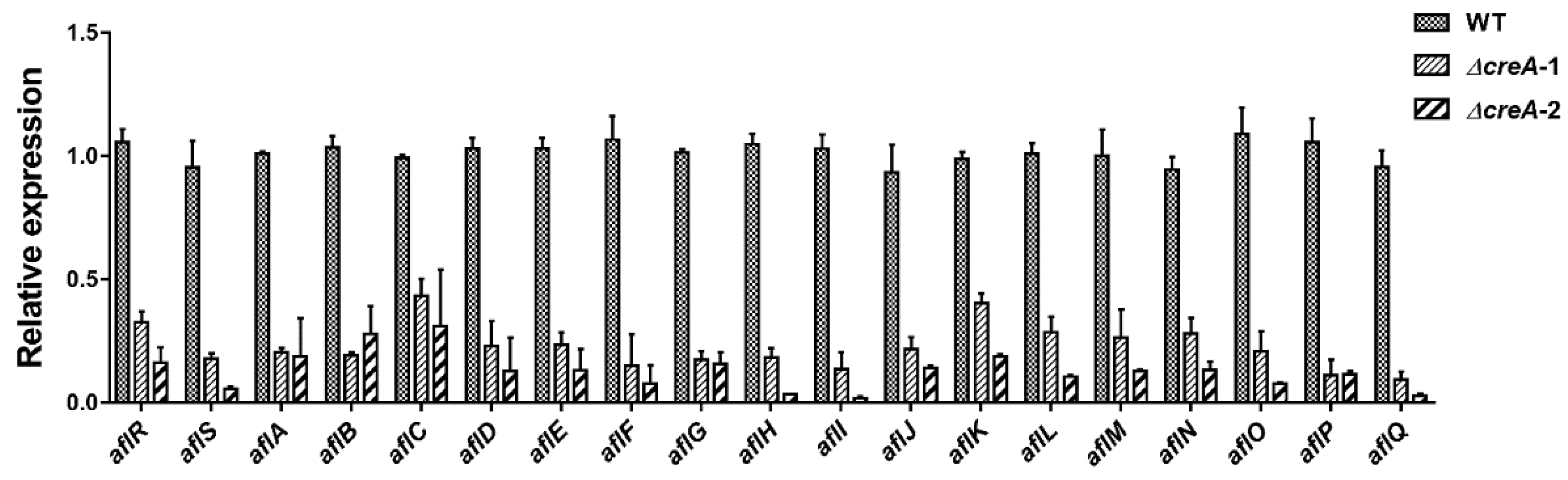
4.5. creA Mutant Construction and qRT-PCR Analysis
4.6. Antioxidant Enzymatic Activities and the Lipid Oxidation Level Measurement
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Nesbitt, B.F.; O’Kelly, J.; Sargeant, K.; Sheridan, A. Aspergillus flavus and turkey X disease. Toxic metabolites of Aspergillus flavus. Nature 1962, 195, 1062–1063. [Google Scholar] [CrossRef] [PubMed]
- Maggio-Hall, L.A.; Wilson, R.A.; Keller, N.P. Fundamental contribution of β-oxidation to polyketide mycotoxin production in planta. Mol. Plant Microbe Interact. 2005, 18, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; Punelli, M.; Smith, C.A.; Zjalic, S.; Scarpari, M.; Scala, V.; Cardinali, G.; Aspite, N.; Pinzari, F.; Payne, G.A.; et al. How peroxisomes affect aflatoxin biosynthesis in Aspergillus flavus. PLoS ONE 2012, 7, e48097. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Liang, Y.; Zhang, J.; Liu, C.M. Aspergillus flavus grown in peptone as the carbon source exhibits spore density- and peptone concentration-dependent aflatoxin biosynthesis. BMC Microbiol. 2012, 12, 106. [Google Scholar] [CrossRef] [PubMed]
- Kiser, R.C.; Niehaus, W.G. Purification and kinetic characterization of mannitol-1-phosphate dehydrogenase from Aspergillus niger. Arch. Biochem. Biophys. 1981, 211, 613–621. [Google Scholar] [CrossRef]
- Niehaus, W.G.; Dilts, R.P. Purification and characterization of mannitol dehydrogenase from Aspergillus parasiticus. J. Bacteriol. 1982, 151, 243–250. [Google Scholar] [PubMed]
- Jayashree, T.; Subramanyam, C. Oxidative stress as a prerequisite for aflatoxin production by Aspergillus parasiticus. Free Radic. Biol. Med. 2000, 29, 981–985. [Google Scholar] [CrossRef]
- Reverberi, M.; Zjalic, S.; Ricelli, A.; Fabbri, A.A.; Fanelli, C. Oxidant/antioxidant balance in Aspergillus parasiticus affects aflatoxin biosynthesis. Mycotoxin Res. 2006, 22, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Narasaiah, K.V.; Sashidhar, R.B.; Subramanyam, C. Biochemical analysis of oxidative stress in the production of aflatoxin and its precursor intermediates. Mycopathologia 2006, 162, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.Y.; Roze, L.V.; Wee, J.; Linz, J.E. Evidence that a transcription factor regulatory network coordinates oxidative stress response and secondary metabolism in aspergilli. Microbiologyopen 2013, 2, 144–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanelli, C.; Fabbri, A.A.; Finotti, E.; Passi, S. Stimulation of aflatoxin biosynthesis by lipophilic epoxides. J. Gen. Microbiol. 1983, 129, 1721–1723. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Q.; Jiang, H.F.; Zhou, Y.Q.; Lei, Y.; Wang, S.Y.; Liao, B.S. Ethylene inhibited aflatoxin biosynthesis is due to oxidative stress alleviation and related to glutathione redox state changes in Aspergillus flavus. Int. J. Food Microbiol. 2009, 130, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; Fabbri, A.A.; Zjalic, S.; Ricelli, A.; Punelli, F.; Fanelli, C. Antioxidant enzymes stimulation in Aspergillus parasiticus by Lentinula edodes inhibits aflatoxin production. Appl. Microbiol. Biotechnol. 2005, 69, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Caceres, I.; El Khoury, R.; Bailly, S.; Oswald, I.P.; Puel, O.; Bailly, J.D. Piperine inhibits aflatoxin B1 production in Aspergillus flavus by modulating fungal oxidative stress response. Fungal Genet. Biol. 2017, 107, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, N.; Molyneux, R.J. Phytochemical inhibition of aflatoxigenicity in Aspergillus flavus by constituents of walnut (Juglans regia). J. Agric. Food Chem. 2004, 52, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Affeldt, K.J.; Keller, N.P. Characterization of the far transcription factor family in Aspergillus flavus. G3 2016, 6, 3269–3281. [Google Scholar] [CrossRef] [PubMed]
- Fasoyin, O.E.; Wang, B.; Qiu, M.; Han, X.; Chung, K.R.; Wang, S. Carbon catabolite repression gene creA regulates morphology, aflatoxin biosynthesis and virulence in Aspergillus flavus. Fungal Genet. Biol. 2018, 115, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Amare, M.G.; Keller, N.P. Molecular mechanisms of Aspergillus flavus secondary metabolism and development. Fungal Genet. Biol. 2014, 66, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Georgianna, D.R.; Fedorova, N.D.; Burroughs, J.L.; Dolezal, A.L.; Bok, J.W.; Horowitz-Brown, S.; Woloshuk, C.P.; Yu, J.; Keller, N.P.; Payne, G.A. Beyond aflatoxin: Four distinct expression patterns and functional roles associated with Aspergillus flavus secondary metabolism gene clusters. Mol. Plant Pathol. 2010, 11, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chang, P.K.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered pathway genes in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science. 2008, 320, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Amaike, S.; Keller, N.P. Distinct roles for VeA and LaeA in development and pathogenesis of Aspergillus flavus. Eukaryot. Cell 2009, 8, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Harris-Coward, P.Y.; Ehrlich, K.C.; Mack, B.M.; Kale, S.P.; Larey, C.; Calvo, A.M. NsdC and NsdD affect Aspergillus flavus morphogenesis and aflatoxin production. Eukaryot. Cell 2012, 11, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Strijbis, K.; van den Burg, J.; Visser, W.F.; van den Berg, M.; Distel, B. Alternative splicing directs dual localization of Candida albicans 6-phosphogluconate dehydrogenase to cytosol and peroxisomes. FEMS Yeast Res. 2012, 12, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, A.; Buchanan, R.L. Regulation of aflatoxin biosynthesis: Induction of aflatoxin production by various carbohydrates. J. Food Sci. 1981, 46, 633–635. [Google Scholar] [CrossRef]
- Maggio-Hall, L.A.; Keller, N.P. Mitochondrial β-oxidation in Aspergillus nidulans. Mol. Microbiol. 2004, 54, 1173–1185. [Google Scholar] [CrossRef] [PubMed]
- Midgley, M. Carnitine acetyltransferase is absent from acuJ mutants of Aspergillus nidulans. FEMS Microbiol. Lett. 1993, 108, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, G.J.; Visser, J. Carbon repression in Aspergilli. FEMS Microbiol. Lett. 1997, 151, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Cubero, B.; Scazzocchio, C. Two different, adjacent and divergent zinc finger binding sites are necessary for CREA-mediated carbon catabolite repression in the proline gene cluster of Aspergillus nidulans. EMBO J. 1994, 13, 407–415. [Google Scholar] [PubMed]
- Balázs, A.; Pócsi, I.; Hamari, Z.; Leiter, É.; Emri, T.; Miskei, M.; Oláh, J.; Tóth, V.; Hegedűs, N.; Prade, R.A.; et al. AtfA bZIP-type transcription factor regulates oxidative and osmotic stress responses in Aspergillus nidulans. Mol. Genet. Genomics 2010, 283, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Arima, T.; Iwashita, K.; Yamada, O.; Gomi, K.; Akita, O. Aspergillus oryzae atfB encodes a transcription factor required for stress tolerance in conidia. Fungal Genet. Biol. 2008, 45, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Toone, W.M.; Morgan, B.A.; Jones, N. Redox control of AP-1-like factors in yeast and beyond. Oncogene 2001, 20, 2336–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.K.; Scharfenstein, L.L.; Luo, M.; Mahoney, N.; Molyneux, R.J.; Yu, J.; Brown, R.L.; Campbell, B.C. Loss of msnA, a putative stress regulatory gene, in Aspergillus parasiticus and Aspergillus flavus increased production of conidia, aflatoxins and kojic acid. Toxins 2011, 3, 82–104. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.H.; Scott, J.B.; Bhaheetharan, J.; Sharpee, W.C.; Milde, L.; Wilson, R.A.; Keller, N.P. Oxygenase coordination is required for morphological transition and the host–fungus interaction of Aspergillus flavus. Mol. Plant Microbe Interact. 2009, 22, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Affeldt, K.J.; Carrig, J.; Amare, M.; Keller, N.P. Global survey of canonical Aspergillus flavus G protein-coupled receptors. mBio 2014, 5, e01501-14. [Google Scholar] [CrossRef] [PubMed]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. RSC Adv. 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- Yabe, K.; Nakajima, H. Enzyme reactions and genes in aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 2004, 64, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, R.L.; Lewis, D.F. Regulation of aflatoxin biosynthesis: Effect of glucose on activities of various glycolytic enzymes. Appl. Environ. Microbiol. 1984, 48, 306–310. [Google Scholar] [PubMed]
- Davis, N.D.; Diener, U.L. Growth and aflatoxin production by Aspergillus parasiticus from various carbon sources. Appl. Microbiol. 1968, 16, 158–159. [Google Scholar] [PubMed]
- Ries, L.N.A.; Beattie, S.R.; Espeso, E.A.; Cramer, R.A.; Goldman, G.H. Diverse regulation of the CreA carbon catabolite repressor in Aspergillus nidulans. Genetics 2016, 203, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Payne, G.A.; Widstrom, N.W. Aflatoxin in maize. Crit. Rev. Plant Sci. 1992, 10, 423–440. [Google Scholar] [CrossRef]
- Pfannenstiel, B.T.; Zhao, X.; Wortman, J.; Wiemann, P.; Throckmorton, K.; Spraker, J.E.; Soukup, A.A.; Luo, X.; Lindner, D.L.; Lim, F.Y.; et al. Revitalization of a forward genetic screen identifies three new regulators of fungal secondary metabolism in the genus Aspergillus. mBio 2017, 8, e01246-17. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Q.; Zhao, X.X.; Wang, C.C.; Xie, Y.; Li, G.H.; He, Z.M. 5-Azacytidine inhibits aflatoxin biosynthesis in Aspergillus flavus. Ann. Microbiol. 2013, 63, 763–769. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]
- Lim, F.Y.; Sanchez, J.F.; Wang, C.C.C.; Keller, N.P. Toward awakening cryptic secondary metabolite gene clusters in filamentous fungi. Methods Enzymol. 2012, 517, 303–324. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, E.; Nayak, T.; Oakley, C.E.; Edgerton, H.; Xiong, Y.; Taheri-Talesh, N.; Osmani, S.A.; Oakley, B.R. Fusion PCR and gene targeting in Aspergillus nidulans. Nat. Protoc. 2007, 1, 3111–3120. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Spraker, J.E.; Bok, J.W.; Velk, T.; He, Z.M.; Keller, N.P. A cellular fusion cascade regulated by LaeA is required for sclerotial development in Aspergillus flavus. Front. Microbiol. 2017, 8, 1925. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | P02_P | P08_P |
|---|---|---|
| Up | 271 | 764 |
| Down | 264 | 1276 |
| Total | 535 | 2040 |
| P02-P a | P08-P b | |||||||
|---|---|---|---|---|---|---|---|---|
| Gene Name | Log2 Ratio c | p Value | Up/Down | Significant e | Log2 Ratio d | p Value | Up/Down | Significant e |
| veA (AFLA_066460) | −0.4470 | 0.6546 | down | no | −0.3915 | 2.64 × 10−1 | down | no |
| laeA (AFLA_033290) | −0.2987 | 0.8134 | down | no | −0.6969 | 1.71 × 10−2 | down | no |
| velB (AFLA_081490) | 0.9331 | 0.1439 | up | no | 0.0635 | 9.35 × 10−1 | up | no |
| brlA (AFLA_082850) | −2.5580 | 0.0010 | down | yes | −2.3343 | 9.88 × 10−6 | down | yes |
| NsdC (AFLA_131330) | 1.6888 | 0.0091 | up | yes | 1.1464 | 3.45 × 10−5 | up | yes |
| creA (AFLA_134680) | −0.3367 | 0.8277 | down | no | −1.1486 | 9.15 × 10−8 | down | yes |
| farB (AFLA_012010) | −0.7799 | 0.4354 | down | no | −1.5190 | 4.81 × 10−5 | down | yes |
| foxA (AFLA_041590) | −0.5139 | 0.6420 | down | no | −1.1589 | 1.71 × 10−5 | down | yes |
| AcuJ (AFLA_135240) | −0.8148 | 0.2928 | down | no | −1.2998 | 1.85 × 10−5 | down | yes |
| PexK (AFLA_036410) | −0.7519 | 0.3792 | down | no | −0.8524 | 2.72 × 10−4 | down | no |
| zwf1 (AFLA_086620) | 0.6412 | 0.4931 | up | no | 0.7590 | 6.7 × 10−3 | up | no |
| sol3 (AFLA_080390) | 0.2110 | 0.8837 | up | no | 1.0991 | 1.0 × 10−4 | up | yes |
| gnd1 (AFLA_036840) | 0.3718 | 0.7936 | up | no | 0.9072 | 1.3 × 10−3 | up | no |
| Gene Name | CreA Binding Motif (5′-SYGGRG-3′) a | Position Relative to Start Codon | Description |
|---|---|---|---|
| AFLA_139190 | GTGGAG | −361 | aflK/vbs/VERB synthase |
| AFLA_139200 | CTGGAG | −795 | aflQ/ordA/ord-1/oxidoreductase/cytochrome P450 monooxigenase |
| AFLA_139210 | No motif found | aflP/omtA/omt-1/O-methyltransferase A | |
| AFLA_139220 | GTGGGG | −1201 | aflO/omtB/dmtA/O-methyltransferase B |
| AFLA_139230 | CTGGGG/CCGGAG/GCGGAG | −355/−398/−301 | aflI/avfA/cytochrome P450 monooxygenase |
| AFLA_139250 | CTGGGG | −238 | aflL/verB/desaturase/P450 monooxygenase |
| AFLA_139260 | CTGGGG/GCGGAG | −365/−810 | aflG/avnA/ord-1/cytochrome P450 monooxygenase |
| AFLA_139280 | No motif found | aflN/verA/monooxygenase | |
| AFLA_139300 | No motif found | aflM/ver-1/dehydrogenase/ketoreductase | |
| AFLA_139310 | CTGGGG/GCGGAG | −482/−617 | aflE/norA/aad/adh-2/NOR reductase/dehydrogenase |
| AFLA_139320 | CCGGGG/CTGGGG | −387/−299 | aflJ/estA/esterase |
| AFLA_139330 | GTGGGG/GCGGAG | −488/−58 | aflH/adhA/short chain alcohol dehydrogenase |
| AFLA_139340 | GTGGAG/CCGGGG | −737/−762 | aflS/pathway regulator |
| AFLA_139360 | GTGGGG/CCGGAG/CTGGAG | −369/−895/−276 | aflR/apa-2/afl-2/transcription activator |
| AFLA_139370 | CTGGAG/CTGGGG | −450/−819 | aflB/fas-1/fatty acid synthase beta subunit |
| AFLA_139380 | GTGGAG | −116 | aflA/fas-2/hexA/fatty acid synthase alpha subunit |
| AFLA_139390 | No motif found | aflD/nor-1/reductase | |
| AFLA_139410 | GTGGCG | −763 | aflC/pksA/pksL1/polyketide synthase |
| AFLA_139440 | No motif found | aflF/norB/dehydrogenase |
| P02-P a | P08-P b | |||||||
|---|---|---|---|---|---|---|---|---|
| Gene Name | Log2 Ratio c | p Value | Up/Down | Significant e | Log2 Ratio d | p Value | Up/Down | Significant e |
| ppoC (AFLA_030430) | −2.0268 | 0.0003 | down | yes | −1.4056 | 7.25 × 10−2 | down | no |
| gprC (AFLA_074150) | −1.4207 | 0.0703 | down | no | −1.5934 | 4.14 × 10−6 | down | yes |
| gprK (AFLA_009790) | −1.2764 | 0.3936 | down | no | −2.1266 | 9.73 × 10−4 | down | yes |
| gprM (AFLA_075000) | −2.0079 | 0.0069 | down | yes | −1.4338 | 3.74 × 10−2 | down | yes |
| glutathione S-transferase (AFLA_016400) | 0.1570 | 0.9701 | up | no | 1.1480 | 1.20 × 10−4 | up | yes |
| glutathione S-transferase (AFLA_023740) | 0.4667 | 0.6766 | up | no | 1.2355 | 2.13 × 10−3 | up | yes |
| glutathione S-transferase (AFLA_087240) | 0.8037 | 0.3272 | up | no | 2.0188 | 1.07 × 10−12 | up | yes |
| AhpC/TSA family thioredoxin peroxidase (AFLA_097940) | 1.1384 | 0.0655 | up | no | 1.2150 | 4.41 × 10−5 | up | yes |
| thioredoxin reductase Trr1/Trr2 (AFLA_051770) | 0.8950 | 0.2535 | up | no | 1.0196 | 3.79 × 10−2 | up | yes |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Zhi, Q.-Q.; Li, J.-Y.; Keller, N.P.; He, Z.-M. The Antioxidant Gallic Acid Inhibits Aflatoxin Formation in Aspergillus flavus by Modulating Transcription Factors FarB and CreA. Toxins 2018, 10, 270. https://doi.org/10.3390/toxins10070270
Zhao X, Zhi Q-Q, Li J-Y, Keller NP, He Z-M. The Antioxidant Gallic Acid Inhibits Aflatoxin Formation in Aspergillus flavus by Modulating Transcription Factors FarB and CreA. Toxins. 2018; 10(7):270. https://doi.org/10.3390/toxins10070270
Chicago/Turabian StyleZhao, Xixi, Qing-Qing Zhi, Jie-Ying Li, Nancy P. Keller, and Zhu-Mei He. 2018. "The Antioxidant Gallic Acid Inhibits Aflatoxin Formation in Aspergillus flavus by Modulating Transcription Factors FarB and CreA" Toxins 10, no. 7: 270. https://doi.org/10.3390/toxins10070270