Unilateral Botulinum Neurotoxin-A Injection into the Striatum of C57BL/6 Mice Leads to a Different Motor Behavior Compared with Rats

 ,
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
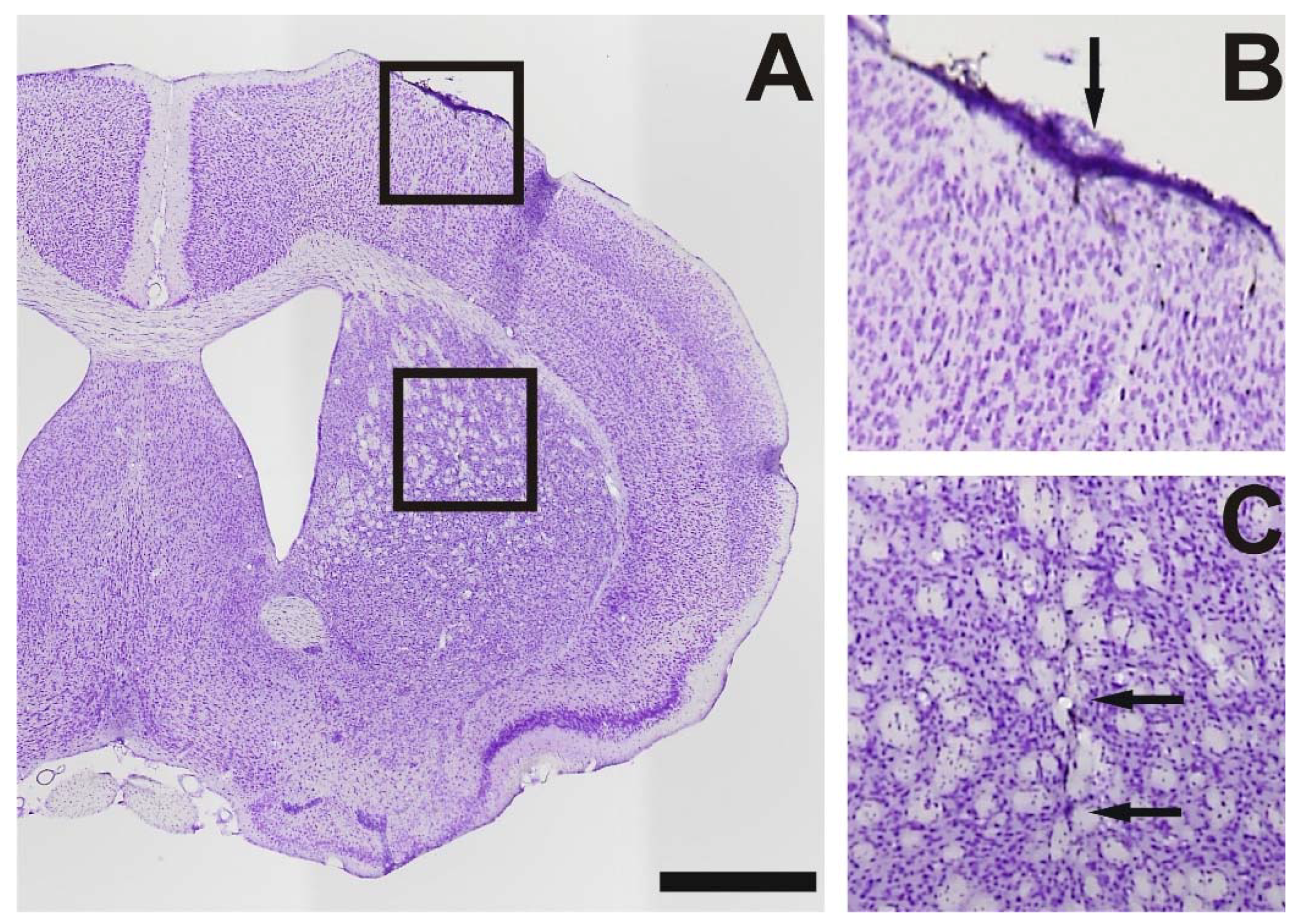{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
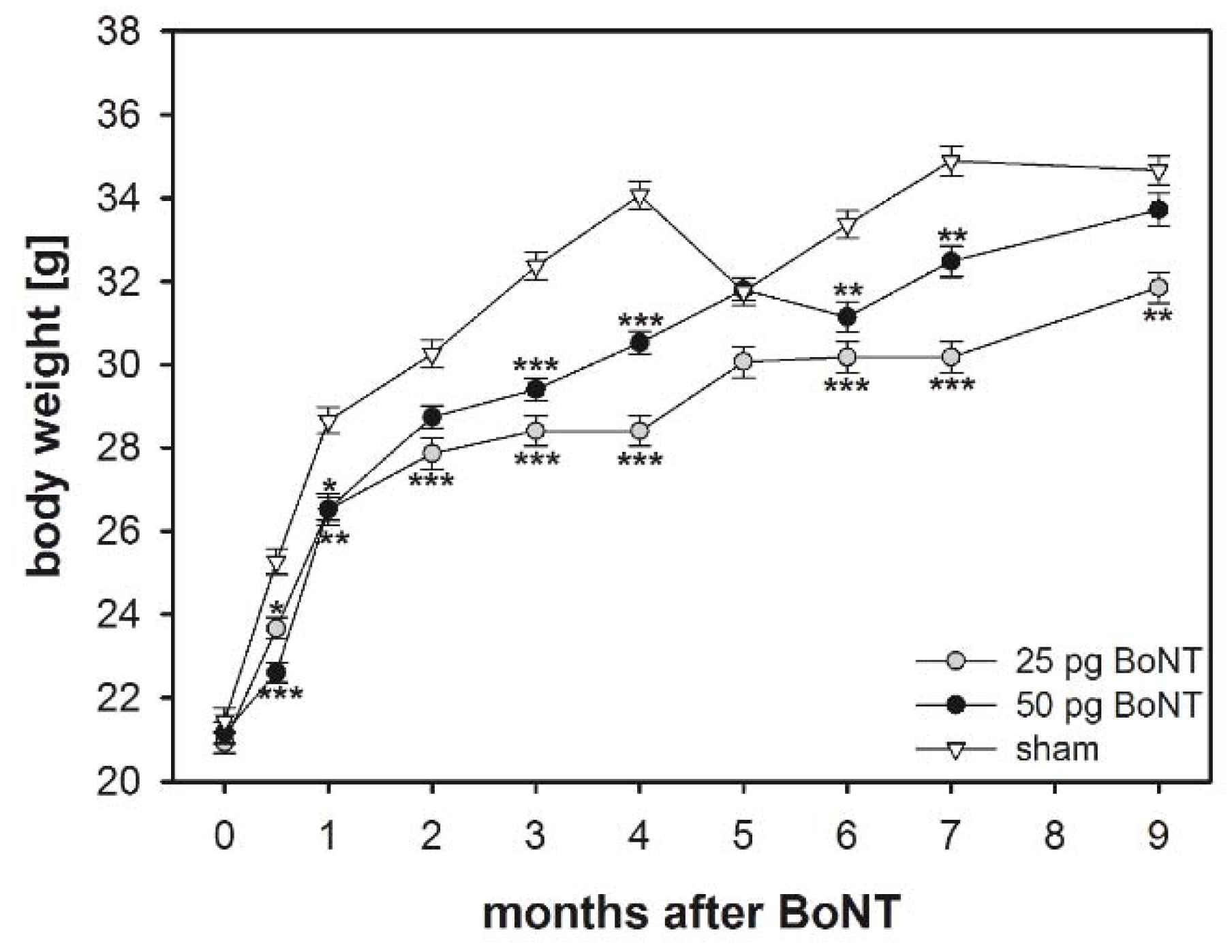
2.1. Body Weight
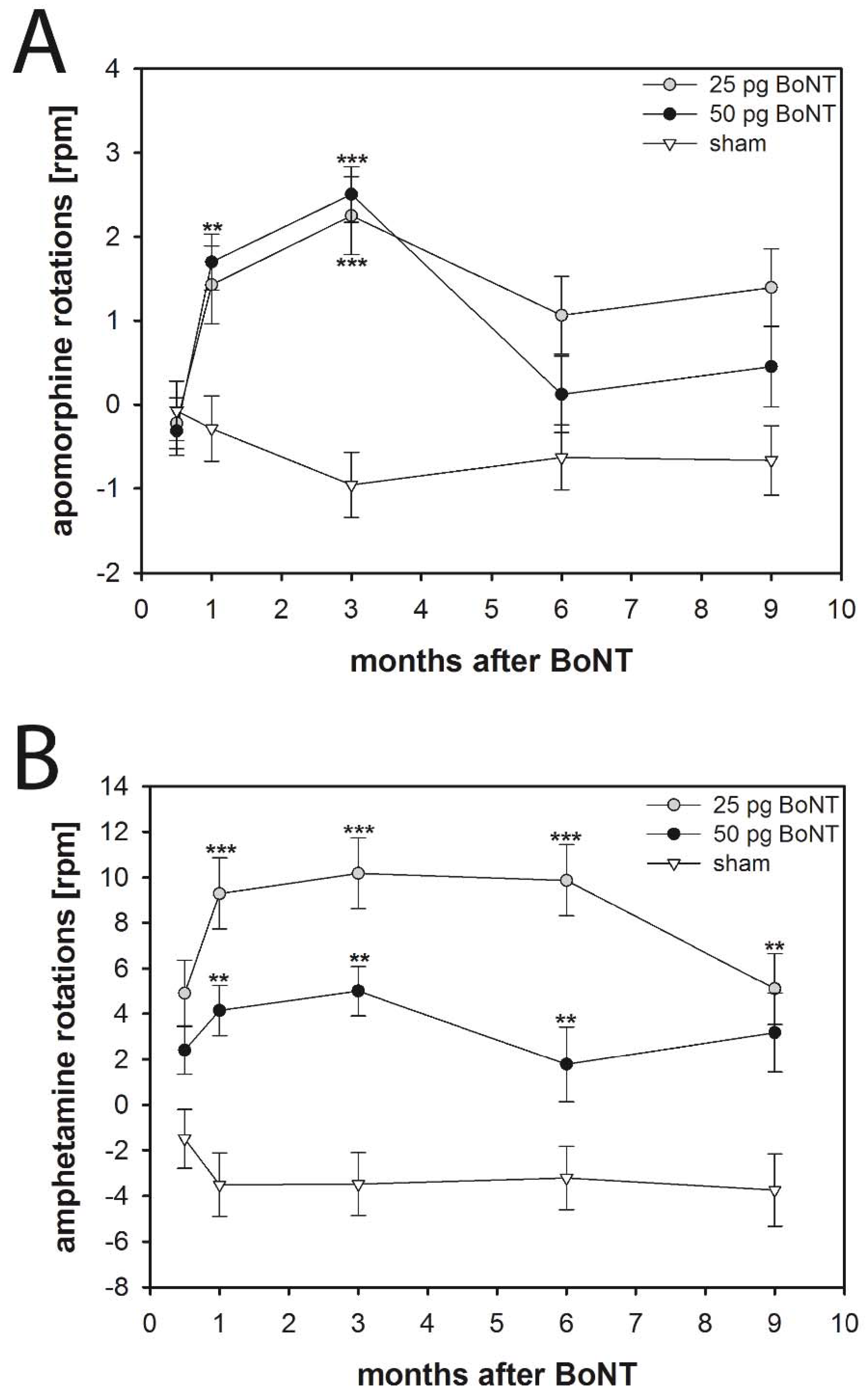
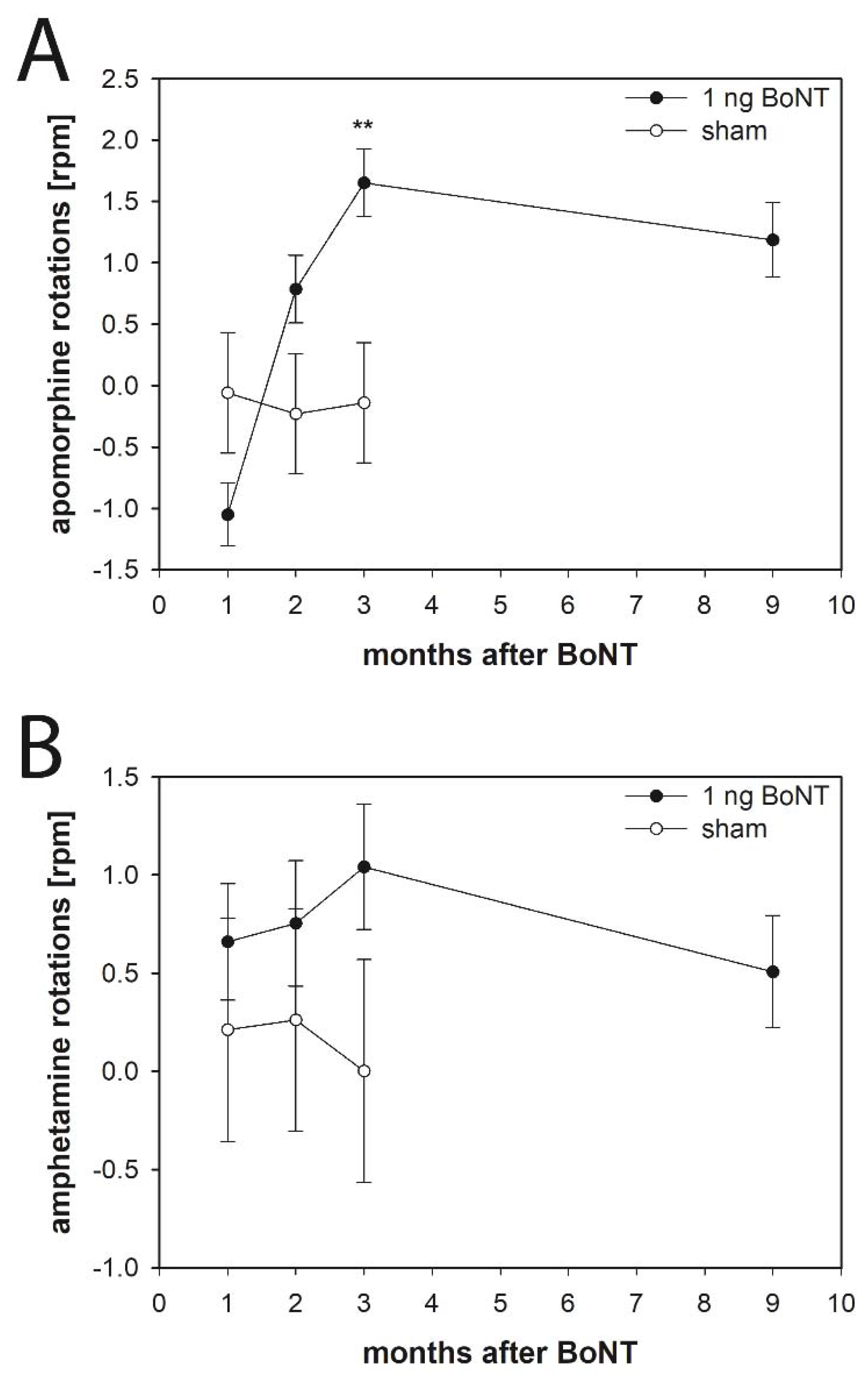
2.2. Apomorphine-Induced Rotations
2.2.1. Mice
2.2.2. Rats
2.3. Amphetamine-Induced Rotations
2.3.1. Mice
2.3.2. Rats
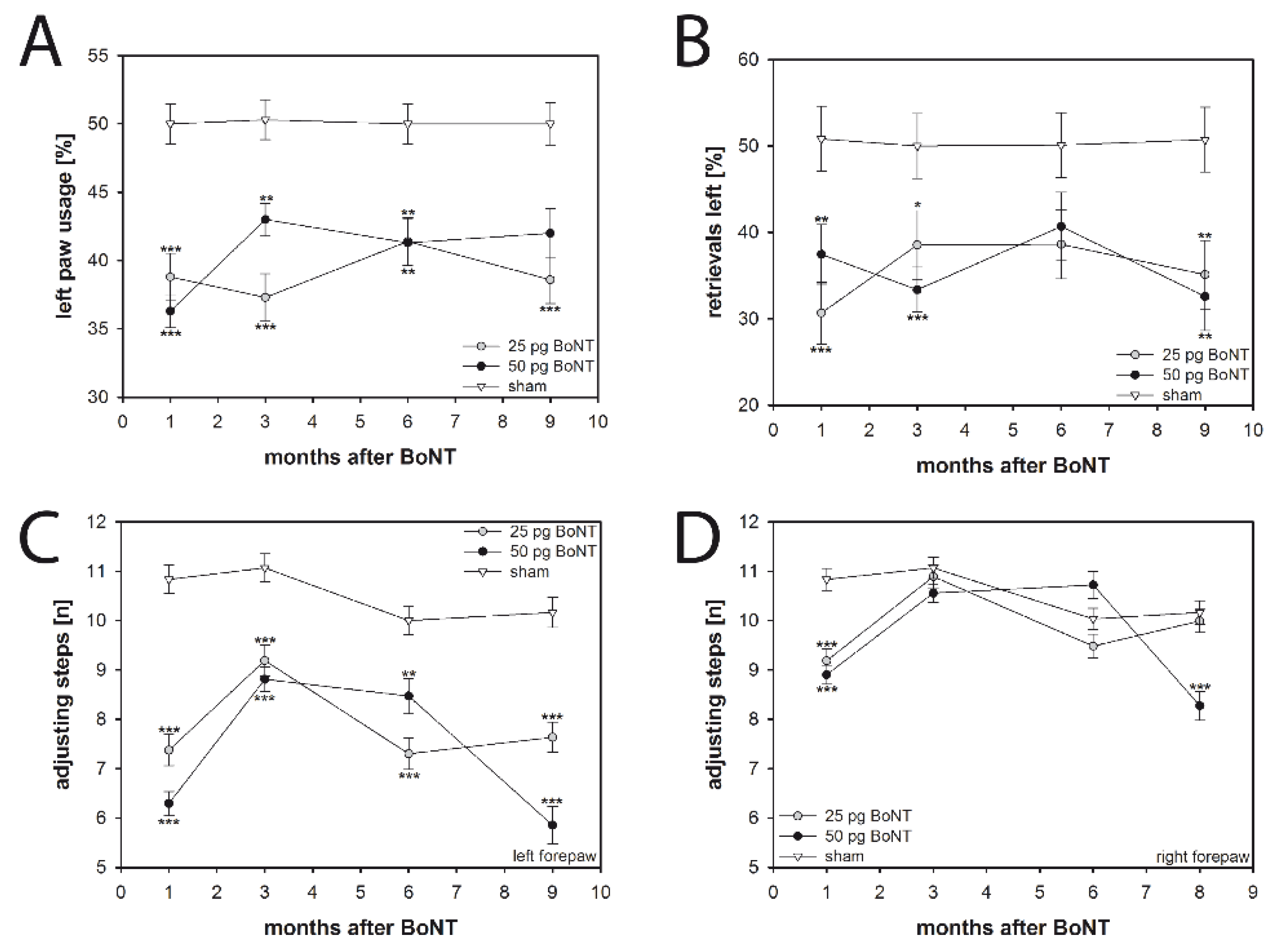
2.4. Cylinder Test
2.5. Corridor Task
2.6. Stepping Test
2.7. Hindlimb Clasping
3. Discussion
3.1. Basal Ganglia Circuitry after BoNT-A Injection
3.2. Body Weight
3.3. Spontaneous Motor Tests
3.3.1. Spontaneous Forelimb Use
3.3.2. Sensorimotor Integration
3.3.3. Forelimb Adjusting Steps
3.3.4. Hindlimb Clasping
3.4. Drug-Induced Rotation Tests
3.4.1. Apomorphine-Induced Rotations
3.4.2. Amphetamine-Induced Rotations
3.4.3. Receptors and Connectomics of the CPu
4. Conclusions and Future Perspectives
5. Materials and Methods
5.1. Animals
5.2. BoNT-A application
5.3. Body Weight
5.4. Behavioral Testing
5.4.1. Drug-Induced Rotation Tests (Apomorphine, Amphetamine)
5.4.2. Spontaneous Motor Tests
Cylinder Test
Corridor Task
Stepping Test
Hindlimb Clasping
5.5. Receptor Autoradiography and Histology
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 6-OHDA | 6-hydroxydopamine |
| BoNT-A | botulinum neurotoxin-A |
| ChAT | choline acetyltransferase |
| CPu | caudate-putamen |
| D1 | dopamine D1 receptor |
| D2 | dopamine D2 receptor |
| GABA | α-amino butyric acid |
| hemi-PD | hemiparkinsonian |
| ir | immunoreactive |
| nic | acetylcholine nicotinic α4β2 receptor |
| PD | Parkinson’s disease |
| SV2C | synaptic vesicle glycoprotein C |
| TH | tyrosine hydroxylase |
References
- Siderowf, A.; Stern, M. Update on Parkinson Disease. Ann. Intern. Med. 2003, 138, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.J.; West, A.B.; Dawson, V.L.; Dawson, T.M. Molecular Pathophysiology of Parkinson’s Disease. Annu. Rev. Neurosci. 2005, 28, 57–87. [Google Scholar] [CrossRef] [PubMed]
- Morin, N.; Jourdain, V.A.; Di Paolo, T. Modeling dyskinesia in animal models of Parkinson disease. Exp. Neurol. 2014, 256, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Albin, R.L.; Young, A.B.; Penney, J.B. The functional anatomy of basal ganglia disorders. Trends Neurosci. 1989, 12, 366–375. [Google Scholar] [CrossRef]
- DeLong, M.R.; Wichmann, T. Circuits and circuit disorders of the basal ganglia. Arch. Neurol. 2007, 64, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Marsden, C.D. The mysterious motor function of the basal ganglia: The Robert Wartenberg Lecture. Neurology 1982, 32, 514–539. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.A.; Rodriguez-Oroz, M.C.; Rodriguez, M.; DeLong, M.R.; Olanow, C.W. Pathophysiology of levodopa-induced dyskinesias in Parkinson’s disease: Problems with the current model. Ann. Neurol. 2000, 47, S22–S32. [Google Scholar] [PubMed]
- Coffield, J.A.; Yan, X. Neuritogenic actions of botulinum neurotoxin A on cultured motor neurons. J. Pharmacol. Exp. Ther. 2009, 330, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Pisani, A.; Bernardi, G.; Ding, J.; Surmeier, D.J. Re-emergence of striatal cholinergic interneurons in movement disorders. Trends Neurosci. 2007, 30, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Guzman, J.N.; Tkatch, T.; Chen, S.; Goldberg, J.A.; Ebert, P.J.; Levitt, P.; Wilson, C.J.; Hamm, H.E.; Surmeier, D.J. RGS4-dependent attenuation of M4 autoreceptor function in striatal cholinergic interneurons following dopamine depletion. Nat. Neurosci. 2006, 9, 832–842. [Google Scholar] [CrossRef] [PubMed]
- Oldenburg, I.A.; Ding, J.B. Cholinergic modulation of synaptic integration and dendritic excitability in the striatum. Curr. Opin. Neurobiol. 2011, 21, 425–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ztaou, S.; Maurice, N.; Camon, J.; Guiraudie-Capraz, G.; Kerkerian-Le Goff, L.; Beurrier, C.; Liberge, M.; Amalric, M. Involvement of Striatal Cholinergic Interneurons and M1 and M4 Muscarinic Receptors in Motor Symptoms of Parkinson’s Disease. J. Neurosci. 2016, 36, 9161–9172. [Google Scholar] [CrossRef] [PubMed]
- Clarke, C.E. Medical Management of Parkinson’s Disease. J. Neurol. Neurosurg. Psychiatry 2002, 72, i22–i27. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, H.H. Updates in the medical management of Parkinson disease. Clevel. Clin. J. Med. 2012, 79, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Connolly, B.S.; Lang, A.E. Pharmacological treatment of Parkinson disease: A review. JAMA 2014, 311, 1670–1683. [Google Scholar] [CrossRef] [PubMed]
- Antipova, V.; Hawlitschka, A.; Mix, E.; Schmitt, O.; Dräger, D.; Benecke, R.; Wree, A. Behavioral and structural effects of unilateral intrastriatal injections of botulinum neurotoxin a in the rat model of Parkinson’s disease. J. Neurosci. Res. 2013, 91, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Hawlitschka, A.; Antipova, V.; Schmitt, O.; Witt, M.; Benecke, R.; Mix, E.; Wree, A. Intracerebrally applied botulinum neurotoxin in experimental neuroscience. Curr. Pharm. Biotechnol. 2013, 14, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Holzmann, C.; Dräger, D.; Mix, E.; Hawlitschka, A.; Antipova, V.; Benecke, R.; Wree, A. Effects of intrastriatal botulinum neurotoxin A on the behavior of Wistar rats. Behav. Brain Res. 2012, 234, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Wree, A.; Mix, E.; Hawlitschka, A.; Antipova, V.; Witt, M.; Schmitt, O.; Benecke, R. Intrastriatal botulinum toxin abolishes pathologic rotational behaviour and induces axonal varicosities in the 6-OHDA rat model of Parkinson’s disease. Neurobiol. Dis. 2011, 41, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Mehlan, J.; Brosig, H.; Schmitt, O.; Mix, E.; Wree, A.; Hawlitschka, A. Intrastriatal injection of botulinum neurotoxin-A is not cytotoxic in rat brain—A histological and stereological analysis. Brain Res. 2016, 1630, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Antipova, V.A.; Holzmann, C.; Schmitt, O.; Wree, A.; Hawlitschka, A. Botulinum Neurotoxin A Injected Ipsilaterally or Contralaterally into the Striatum in the Rat 6-OHDA Model of Unilateral Parkinson’s Disease Differently Affects Behavior. Front. Behav. Neurosci. 2017, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Wedekind, F.; Oskamp, A.; Lang, M.; Hawlitschka, A.; Zilles, K.; Wree, A.; Bauer, A. Intrastriatal administration of botulinum neurotoxin A normalizes striatal D2R binding and reduces striatal D1R binding in male hemiparkinsonian rats. J. Neurosci. Res. 2018, 96, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Mann, T.; Zilles, K.; Dikow, H.; Hellfritsch, A.; Cremer, M.; Piel, M.; Rösch, F.; Hawlitschka, A.; Schmitt, O.; Wree, A. Dopamine, Noradrenaline and Serotonin Receptor Densities in the Striatum of Hemiparkinsonian Rats following Botulinum Neurotoxin-A Injection. Neuroscience 2018, 374, 187–204. [Google Scholar] [CrossRef] [PubMed]
- Hawlitschka, A.; Holzmann, C.; Witt, S.; Spiewok, J.; Neumann, A.M.; Schmitt, O.; Wree, A.; Antipova, V. Intrastriatally injected botulinum neurotoxin-A differently effects cholinergic and dopaminergic fibers in C57BL/6 mice. Brain Res. 2017, 1676, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Corchs, F.; Nutt, D.J.; Hince, D.A.; Davies, S.J.; Bernik, M.; Hood, S.D. Evidence for serotonin function as a neurochemical difference between fear and anxiety disorders in humans? J. Psychopharmacol. 2015, 29, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Ellenbroek, B.; Youn, J. Rodent models in neuroscience research: Is it a rat race? Dis. Model. Mech. 2016, 9, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, S.; Zador, A.M. Mice and rats achieve similar levels of performance in an adaptive decision-making task. Front. Syst. Neurosci. 2014, 8, 173. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Westenberger, A.; Hollingworth, P.; Harold, D.; Jones, L.; Owen, M.J.; Williams, J.; Marques, S.C.F.; Oliveira, C.R.; Pereira, C.M.F.; et al. Genetics of Parkinson’s Disease. Int. J. Geriatr. Psychiatry 2012, 26, a008888. [Google Scholar] [CrossRef] [PubMed]
- Lazarov, O.; Hollands, C. Hippocampal neurogenesis: Learning to remember. Prog. Neurobiol. 2016, 138–140, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Lindström, M.; Korkeala, H. Laboratory diagnostics of botulism. Clin. Microbiol. Rev. 2006, 19, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Overgaard, A.; Tena-Sempere, M.; Franceschini, I.; Desroziers, E.; Simonneaux, V.; Mikkelsen, J.D. Comparative analysis of kisspeptin-immunoreactivity reveals genuine differences in the hypothalamic Kiss1 systems between rats and mice. Peptides 2013, 45, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Sesardic, D.; Das, R.G. Alternatives to the LD50 assay for botulinum toxin potency testing: Strategies and progress towards refinement, reduction and replacement. AATEX 2008, 14, 581–585. [Google Scholar]
- Wheeler, C.; Inami, G.; Mohle-Boetani, J.; Vugia, D. Sensitivity of mouse bioassay in clinical wound botulism. Clin. Infect. Dis. 2009, 48, 1669–1673. [Google Scholar] [CrossRef] [PubMed]
- Antonucci, F.; Rossi, C.; Gianfranceschi, L.; Rossetto, O.; Caleo, M. Long-distance retrograde effects of botulinum neurotoxin A. J. Neurosci. 2008, 28, 3689–3696. [Google Scholar] [CrossRef] [PubMed]
- De Leonibus, E.; Costantini, V.J.A.; Massaro, A.; Mandolesi, G.; Vanni, V.; Luvisetto, S.; Pavone, F.; Oliverio, A.; Mele, A. Cognitive and neural determinants of response strategy in the dual-solution plus-maze task. Learn. Mem. 2011, 18, 241–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caleo, M.; Restani, L.; Gianfranceschi, L.; Costantin, L.; Rossi, C.; Rossetto, O.; Montecucco, C.; Maffei, L. Transient Synaptic Silencing of Developing Striate Cortex Has Persistent Effects on Visual Function and Plasticity. J. Neurosci. 2007, 27, 4530–4540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiavo, G.; Montecucco, C. Tetanus and Botulism Neurotoxins: Isolation and Assay. Methods Enzymol. 1995, 248, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Montecucco, C.; Schiavo, G. Structure and Function of Tetanus and Botulinum Neurotoxins. Q. Rev. Biophys. 1995, 28, 423–472. [Google Scholar] [CrossRef] [PubMed]
- Cremer, J.N.N.; Amunts, K.; Schleicher, A.; Palomero-Gallagher, N.; Piel, M.; Rösch, F.; Zilles, K. Changes in the expression of neurotransmitter receptors in Parkin and DJ-1 knockout mice—A quantitative multireceptor study. Neuroscience 2015, 311, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Cremer, J.N.; Amunts, K.; Graw, J.; Piel, M.; Rösch, F.; Zilles, K. Neurotransmitter receptor density changes in Pitx3ak mice—A model relevant to parkinson’s disease. Neuroscience 2015, 285, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Orth, M.; Tabrizi, S.J. Models of Parkinson’s disease. Mov. Disord. 2003, 18, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Le, W.; Jankovic, J. Animal models of Parkinson’s disease. Park. Dis. Diagn. Mot. Symptoms Non-Mot. Features 2013, 115, 115–135. [Google Scholar] [CrossRef]
- Wong, P.C.; Cai, H.; Borchelt, D.R.; Price, D.L. Genetically engineered mouse models of neurodegenerative diseases. Nat. Neurosci. 2002, 5, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Magen, I.; Chesselet, M.-F. Genetic mouse models of Parkinson’s disease. Recent Adv. Park. Dis. Clin. Res. 2010, 184, 53–87. [Google Scholar] [CrossRef]
- Blandini, F.; Armentero, M.-T. Animal models of Parkinson’s disease. FEBS J. 2012, 279, 1156–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, M.; Wang, Z.; Ding, J.; An, X.; Ingham, C.A.; Shering, A.F.; Wokosin, D.; Ilijic, E.; Sun, Z.; Sampson, A.R.; et al. Selective elimination of glutamatergic synapses on striatopallidal neurons in Parkinson disease models. Nat. Neurosci. 2006, 9, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.Á.; Marin, C.; Rodriguez-Oroz, C.; Blesa, J.; Benitez-Temiño, B.; Mena-Segovia, J.; Rodríguez, M.; Olanow, C.W. The basal ganglia in Parkinson’s disease: Current concepts and unexplained observations. Ann. Neurol. 2008, 64, S30–S46. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.Á.; Rodríguez-Oroz, M.C.; Benitez-Temino, B.; Blesa, F.J.; Guridi, J.; Marin, C.; Rodriguez, M. Functional organization of the basal ganglia: Therapeutic implications for Parkinson’s disease. Mov. Disord. 2008, 23, S548–S559. [Google Scholar] [CrossRef] [PubMed]
- Bordia, T.; Zhang, D.; Perez, X.A.; Quik, M. Striatal cholinergic interneurons and D2 receptor-expressing GABAergic medium spiny neurons regulate tardive dyskinesia. Exp. Neurol. 2016, 286, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Hurley, M.J.; Jenner, P. What has been learnt from study of dopamine receptors in Parkinson’s disease? Pharmacol. Ther. 2006, 111, 715–728. [Google Scholar] [CrossRef] [PubMed]
- Mamaligas, A.A.; Cai, Y.; Ford, C.P. Nicotinic and opioid receptor regulation of striatal dopamine D2-receptor mediated transmission. Sci. Rep. 2016, 6, 37834. [Google Scholar] [CrossRef] [PubMed]
- Perreault, M.L.; Hasbi, A.; O’Dowd, B.F.; George, S.R. The dopamine D1–D2 receptor heteromer in striatal medium spiny neurons: Evidence for a third distinct neuronal pathway in basal ganglia. Front. Neuroanat. 2011, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Rico, A.J.; Dopeso-Reyes, I.G.; Martínez-Pinilla, E.; Sucunza, D.; Pignataro, D.; Roda, E.; Marín-Ramos, D.; Labandeira-García, J.L.; George, S.R.; Franco, R.; et al. Neurochemical evidence supporting dopamine D1–D2 receptor heteromers in the striatum of the long-tailed macaque: Changes following dopaminergic manipulation. Brain Struct. Funct. 2017, 222, 1767–1784. [Google Scholar] [CrossRef] [PubMed]
- Ariano, M.A.; Stromski, C.J.; Smyk-Randall, E.M.; Sibley, D.R. D2 dopamine receptor localization on striatonigral neurons. Neurosci. Lett. 1992, 144, 215–220. [Google Scholar] [CrossRef]
- Brock, J.W.; Farooqui, S.; Ross, K.; Prasad, C. Localization of dopamine D2 receptor protein in rat brain using polyclonal antibody. Brain Res. 1992, 578, 244–250. [Google Scholar] [CrossRef]
- Yung, K.K.L.; Bolam, J.P.; Smith, A.D.; Hersch, S.M.; Ciliax, B.J.; Levey, A.I. Immunocytochemical localization of D1 and D2 dopamine receptors in the basal ganglia of the rat: Light and electron microscopy. Neuroscience 1995, 65, 709–730. [Google Scholar] [CrossRef]
- Li, N.; Chen, T.-W.; Guo, Z.V.; Gerfen, C.R.; Svoboda, K. A motor cortex circuit for motor planning and movement. Nature 2015, 519, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Kirik, D.; Rosenblad, C.; Bjorklund, A. Preservation of a functional nigrostriatal dopamine pathway by GDNF in the intrastriatal 6-OHDA lesion model depends on the site of administration of the trophic factor. Eur. J. Neurosci. 2000, 12, 3871–3882. [Google Scholar] [CrossRef] [PubMed]
- Dowd, E.; Monville, C.; Torres, E.M.; Dunnett, S.B. The Corridor Task: A simple test of lateralised response selection sensitive to unilateral dopamine deafferentation and graft-derived dopamine replacement in the striatum. Brain Res. Bull. 2005, 68, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, D.F.; Moloney, T.C.; Dowd, E. Further validation of the corridor task for assessing deficit and recovery in the hemi-Parkinsonian rat: Restoration of bilateral food retrieval by dopamine receptor agonism. Behav. Brain Res. 2006, 169, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Grealish, S.; Mattsson, B.; Draxler, P.; Björklund, A. Characterisation of behavioural and neurodegenerative changes induced by intranigral 6-hydroxydopamine lesions in a mouse model of Parkinson’s disease. Eur. J. Neurosci. 2010, 31, 2266–2278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starkey, M.L.; Barritt, A.W.; Yip, P.K.; Davies, M.; Hamers, F.P.T.; McMahon, S.B.; Bradbury, E.J. Assessing behavioural function following a pyramidotomy lesion of the corticospinal tract in adult mice. Exp. Neurol. 2005, 195, 524–539. [Google Scholar] [CrossRef] [PubMed]
- Welniarz, Q.; Dusart, I.; Gallea, C.; Roze, E. One hand clapping: Lateralization of motor control. Front. Neuroanat. 2015, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Blume, S.R.; Cass, D.K.; Tseng, K.Y. Stepping test in mice: A reliable approach in determining forelimb akinesia in MPTP-induced Parkinsonism. Exp. Neurol. 2009, 219, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Heuer, A.; Smith, G.A.; Lelos, M.J.; Lane, E.L.; Dunnett, S.B. Unilateral nigrostriatal 6-hydroxydopamine lesions in mice I: Motor impairments identify extent of dopamine depletion at three different lesion sites. Behav. Brain Res. 2012, 228, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Baskin, Y.K.; Dietrich, W.D.; Green, E.J. Two effective behavioral tasks for evaluating sensorimotor dysfunction following traumatic brain injury in mice. J. Neurosci. Methods 2003, 129, 87–93. [Google Scholar] [CrossRef]
- Cohen, N.R.; Taylor, J.S.; Scott, L.B.; Guillery, R.W.; Soriano, P.; Furley, A.J. Errors in corticospinal axon guidance in mice lacking the neural cell adhesion molecule L1. Curr. Biol. 1998, 8, 26–33. [Google Scholar] [CrossRef]
- Steward, O.; Zheng, B.; Ho, C.; Anderson, K.; Tessier-Lavigne, M. The Dorsolateral Corticospinal Tract in Mice: An Alternative Route for Corticospinal Input to Caudal Segments following Dorsal Column Lesions. J. Comp. Neurol. 2004, 472, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Glajch, K.E.; Fleming, S.M.; Surmeier, D.J.; Osten, P. Sensorimotor assessment of the unilateral 6-hydroxydopamine mouse model of Parkinson’s disease. Behav. Brain Res. 2012, 230, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Boix, J.; Padel, T.; Paul, G. A partial lesion model of Parkinson’s disease in mice—Characterization of a 6-OHDA-induced medial forebrain bundle lesion. Behav. Brain Res. 2015, 284, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Fernagut, P.O.; Diguet, E.; Bioulac, B.; Tison, F. MPTP potentiates 3-nitropropionic acid-induced striatal damage in mice: Reference to striatonigral degeneration. Exp. Neurol. 2004, 185, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Guyenet, S.J.; Furrer, S.A.; Damian, V.M.; Baughan, T.D.; La Spada, A.R.; Garden, G.A. A simple composite phenotype scoring system for evaluating mouse models of cerebellar ataxia. J. Vis. Exp. 2010, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.; Koyama, A.; Masliah, E.; Mucke, L. Tau reduction does not prevent motor deficits in two mouse models of Parkinson’s disease. PLoS ONE 2011, 6, e29257. [Google Scholar] [CrossRef] [PubMed]
- Lieu, C.A.; Chinta, S.J.; Rane, A.; Andersen, J.K. Age-Related Behavioral Phenotype of an Astrocytic Monoamine Oxidase-B Transgenic Mouse Model of Parkinson’s Disease. PLoS ONE 2013, 8, e54200. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.D.; Weiss, B. Reversal of supersensitive apomorphine-induced rotational behavior in mice by continuous exposure to apomorphine. J. Pharmacol. Exp. Ther. 1986, 238, 242–247. [Google Scholar] [PubMed]
- Ford, C.P. The role of D2-autoreceptors in regulating dopamine neuron activity and transmission. Neuroscience 2014, 282, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha, C.; Wietzikoski, E.C.; Ferro, M.M.; Martinez, G.R.; Vital, M.A.B.F.; Hipólide, D.; Tufik, S.; Canteras, N.S. Hemiparkinsonian rats rotate toward the side with the weaker dopaminergic neurotransmission. Behav. Brain Res. 2008, 189, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, A.E.; Volz, T.J.; Riddle, E.L.; Gibb, J.W.; Hanson, G.R. New insights into the mechanism of action of amphetamines. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 681–698. [Google Scholar] [CrossRef] [PubMed]
- Del Arco, A.; González-Mora, J.L.; Armas, V.R.; Mora, F. Amphetamine increases the extracellular concentration of glutamate in striatum of the awake rat: Involvement of high affinity transporter mechanisms. Neuropharmacology 1999, 38, 943–954. [Google Scholar] [CrossRef]
- Del Arco, A.; Castañeda, T.R.; Mora, F. Amphetamine releases GABA in striatum of the freely moving rat: Involvement of calcium and high affinity transporter mechanisms. Neuropharmacology 1998, 37, 199–205. [Google Scholar] [CrossRef]
- Hernandez, L.; Lee, F.; Hoebel, B.G. Simultaneous microdialysis and amphetamine infusion in the nucleus accumbens and striatum of freely moving rats: Increase in extracellular dopamine and serotonin. Brain Res. Bull. 1987, 19, 623–628. [Google Scholar] [CrossRef]
- Mandel, R.J.; Leanza, G.; Nilsson, O.G.; Rosengren, E. Amphetamine induces excess release of striatal acetylcholine in vivo that is independent of nigrostriatal dopamine. Brain Res. 1994, 653, 57–65. [Google Scholar] [CrossRef]
- Mora, F.; Porras, A. Effects of amphetamine on the release of excitatory amino acid neurotransmitters in the basal ganglia of the conscious rat. Can. J. Physiol. Pharmacol. 1993, 71, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Miele, M.; Mura, M.A.; Enrico, P.; Esposito, G.; Serra, P.A.; Migheli, R.; Zangani, D.; Miele, E.; Desole, M.S. On the mechanism of d-amphetamine-induced changes in glutamate, ascorbic acid and uric acid release in the striatum of freely moving rats. Br. J. Pharmacol. 2000, 129, 582–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulzer, D.; Sonders, M.S.; Poulsen, N.W.; Galli, A. Mechanisms of neurotransmitter release by amphetamines: A review. Prog. Neurobiol. 2005, 75, 406–433. [Google Scholar] [CrossRef] [PubMed]
- Butcher, S.P.; Fairbrother, I.S.; Kelly, J.S.; Arbuthnott, G.W. Amphetamine-Induced Dopamine Release in the Rat Striatum: An In Vivo Microdialysis Study. J. Neurochem. 1988, 50, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Kroken, A.R.; Blum, F.C.; Zuverink, M.; Barbieri, J.T. Entry of Botulinum neurotoxin subtypes A1 and A2 into neurons. Infect. Immun. 2016, 85, IAI.00795-16. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Yeh, F.; Tepp, W.H.; Dean, C.; Johnson, E.A.; Janz, R.; Chapman, E.R. SV2 Is the Protein Receptor for Botulinum Neurotoxin A. Science 2006, 312, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Dardou, D.; Dassesse, D.; Cuvelier, L.; Deprez, T.; De Ryck, M.; Schiffmann, S.N. Distribution of SV2C mRNA and protein expression in the mouse brain with a particular emphasis on the basal ganglia system. Brain Res. 2011, 1367, 130–145. [Google Scholar] [CrossRef] [PubMed]
- Dardou, D.; Monlezun, S.; Foerch, P.; Courade, J.P.; Cuvelier, L.; De Ryck, M.; Schiffmann, S.N. A role for SV2C in basal ganglia functions. Brain Res. 2013, 1507, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Janz, R.; Südhof, T.C. SV2C is a synaptic vesicle protein with an unusually restricted localization: Anatomy of a synaptic vesicle protein family. Neuroscience 1999, 94, 1279–1290. [Google Scholar] [CrossRef]
- Bajjalieh, S.M.; Peterson, K.; Linial, M.; Scheller, R.H. Brain contains two forms of synaptic vesicle protein 2. Proc. Natl. Acad. Sci. USA 1993, 90, 2150–2154. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, N.; Saito, A.; Karasawa, T.; Harita, Y.; Suzuki, K.; Koike, H.; Han, G.D.; Shimizu, F.; Kawachi, H. Synaptic vesicle protein 2B is expressed in podocyte, and its expression is altered in proteinuric glomeruli. J. Am. Soc. Nephrol. 2006, 17, 2748–2759. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.M.; Janz, R.; Belizaire, R.; Frishman, L.J.; Sherry, D.M. Differential distribution and developmental expression of synaptic vesicle protein 2 isoforms in the mouse retina. J. Comp. Neurol. 2003, 460, 106–122. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Yamamoto, A.; Yatsushiro, S.; Yamada, H.; Futai, M.; Yamaguchi, A.; Moriyama, Y. Synaptic vesicle protein SV2B, but not SV2A, is predominantly expressed and associated with microvesicles in rat pinealocytes. J. Neurochem. 1998, 71, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Ashton, A.C.; Dolly, J.O. Characterization of the Inhibitory Action of Botulinum Neurotoxin Type A on the Release of Several Transmitters from Rat Cerebrocortical Synaptosomes. J. Neurochem. 1988, 50, 1808–1816. [Google Scholar] [CrossRef] [PubMed]
- Bigalke, H.; Heller, I.; Bizzini, B.; Habermann, E. Tetanus toxin and botulinum A toxin inhibit release and uptake of various transmitters, as studied with particulate preparations from rat brain and spinal cord. Naunyn. Schmiedebergs. Arch. Pharmacol. 1981, 316, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Bozzi, Y.; Costantin, L.; Antonucci, F.; Caleo, M. Action of botulinum neurotoxins in the central nervous system: Antiepileptic effects. Neurotox. Res. 2006, 9, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Dunn, A.R.; Stout, K.A.; Ozawa, M.; Lohr, K.M.; Hoffman, C.A.; Bernstein, A.I.; Li, Y.; Wang, M.; Sgobio, C.; Sastry, N.; et al. Synaptic vesicle glycoprotein 2C (SV2C) modulates dopamine release and is disrupted in Parkinson disease. Proc. Natl. Acad. Sci. USA 2017, 114, E2253–E2262. [Google Scholar] [CrossRef] [PubMed]
- Mahrhold, S.; Rummel, A.; Bigalke, H.; Davletov, B.; Binz, T. The synaptic vesicle protein 2C mediates the uptake of botulinum neurotoxin A into phrenic nerves. FEBS Lett. 2006, 580, 2011–2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, L.B.; First, E.R.; MacCallum, R.D.; Gupta, A. Pharmacologic characterization of botulinum toxin for basic science and medicine. Toxicon 1997, 35, 1373–1412. [Google Scholar] [CrossRef]
- Mann, T.; Zilles, K.; Klawitter, F.; Cremer, M.; Hawlitschka, A.; Palomero-Gallagher, N.; Schmitt, O.; Wree, A. Acetylcholine neurotransmitter receptor densities in the striatum of hemiparkinsonian rats following Botulinum neurotoxin-A injection. Front. Neuroanat. 2018, in press. [Google Scholar]
- Mann, T.; Zilles, K.; Frederike, V.; Höhmann, K.; Hellfritsch, A.; Van Bonn, S.; Cremer, M.; Schmitt, O.; Hawlitschka, A.; Wree, A. Glutamate, GABA and adenosine neurotransmitter receptor densities in the striatum of hemiparkinsonian rats following Botulinum neurotoxin-A injection. 2018; in press. [Google Scholar]
- Oh, S.W.; Harris, J.A.; Ng, L.; Winslow, B.; Cain, N.; Mihalas, S.; Wang, Q.; Lau, C.; Kuan, L.; Henry, A.M.; et al. A mesoscale connectome of the mouse brain. Nature 2014, 508, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerfen, C.R. Basal Ganglia. In The Rat Nervous System; Toga, A.W., Ed.; Elsevier Academic: Amsterdam, The Netherlands, 1994; pp. 217–227. ISBN 9780123970251. [Google Scholar]
- Klockgether, T. Medikamentöse behandlung der idiopathischen Parkinson-krankheit. Nervenarzt 2003, 74, S12–S21. [Google Scholar] [CrossRef] [PubMed]
- Horstink, M.; Tolosa, E.; Bonuccelli, U.; Deuschl, G.; Friedman, A.; Kanovsky, P.; Larsen, J.P.; Lees, A.; Oertel, W.; Poewe, W.; et al. Review of the therapeutic management of Parkinson’s disease. Report of a joint task force of the European Federation of Neurological Societies and the Movement Disorder Society-European Section. Part I: Early (uncomplicated) Parkinson’s disease. Eur. J. Neurol. 2006, 13, 1170–1185. [Google Scholar] [CrossRef] [PubMed]
- Whitney, C.M. Medications for Parkinson’s disease. Neurologist 2007, 13, 387–388. [Google Scholar] [CrossRef] [PubMed]
- Orsini, M.; Leite, M.A.A.; Chung, T.M.; Bocca, W.; de Souza, J.A.; de Souza, O.G.; Moreira, R.P.; Bastos, V.H.; Teixeira, S.; Oliveira, A.B.; et al. Botulinum neurotoxin type A in neurology: Update. Neurol. Int. 2015, 7, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Chen, S. Clinical uses of botulinum neurotoxins: Current indications, limitations and future developments. Toxins 2012, 4, 913–939. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, J. An update on new and unique uses of botulinum toxin in movement disorders. Toxicon 2017, 147, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, J. Botulinum toxin: State of the art. Mov. Disord. 2017, 32, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Bezard, E.; Yue, Z.; Kirik, D.; Spillantini, M.G. Animal models of Parkinson’s disease: Limits and relevance to neuroprotection studies. Mov. Disord. 2013, 28, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Yuan, L. Genetic variants and animal models in SNCA and Parkinson disease. Ageing Res. Rev. 2014, 15, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Pickrell, A.M.; Pinto, M.; Moraes, C.T. Mouse models of Parkinson’s disease associated with mitochondrial dysfunction. Mol. Cell. Neurosci. 2013, 55, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Gasior, M.; Tang, R.; Rogawski, M.A. Long-lasting attenuation of amygdala-kindled seizures after convection-enhanced delivery of botulinum neurotoxins a and B into the amygdala in rats. J. Pharmacol. Exp. Ther. 2013, 346, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Antonucci, F.; Bozzi, Y.; Caleo, M. Intrahippocampal infusion of botulinum neurotoxin E (BoNT/E) reduces spontaneous recurrent seizures in a mouse model of mesial temporal lobe epilepsy. Epilepsia 2009, 50, 963–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates; Academic Press: Amsterdam, The Netherlands, 2001; ISBN 0125476361. [Google Scholar]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 5th ed.; Elsevier Academic Press: Amsterdam, The Netherlands; London, UK, 2005; ISBN 0120885727. [Google Scholar]
- Pirazzini, M.; Rossetto, O.; Eleopra, R.; Montecucco, C. Botulinum Neurotoxins: Biology, Pharmacology, and Toxicology. Pharmacol. Rev. 2017, 69, 200–235. [Google Scholar] [CrossRef] [PubMed]
- Frevert, J. Content of botulinum neurotoxin in botox®vistabel®, dysport®azzalure®, and xeomin®bocouture. Drugs R D 2010, 10, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Scaglione, F. Conversion ratio between botox®, dysport®, and xeomin® in clinical practice. Toxins 2016, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Bensadoun, J.C.; Deglon, N.; Tseng, J.L.; Ridet, J.L.; Zurn, A.D.; Aebischer, P. Lentiviral vectors as a gene delivery system in the mouse midbrain: Cellular and behavioral improvements in a 6-OHDA model of Parkinson’s disease using GDNF. Exp. Neurol. 2000, 164, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Iancu, R.; Mohapel, P.; Brundin, P.; Paul, G. Behavioral characterization of a unilateral 6-OHDA-lesion model of Parkinson’s disease in mice. Behav. Brain Res. 2005, 162, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Da Conceição, F.S.L.; Ngo-Abdalla, S.; Houzel, J.-C.; Rehen, S.K. Murine model for Parkinson’s disease: From 6-OH dopamine lesion to behavioral test. J. Vis. Exp. 2010, 35, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Brundin, P.; Isacson, O.; Gage, F.H.; Prochiantz, A.; Björklund, A. The rotating 6-hydroxydopamine-lesioned mouse as a model for assessing functional effects of neuronal grafting. Brain Res. 1986, 366, 346–349. [Google Scholar] [CrossRef]
- Fu, A.; Zhou, Q.H.; Hui, E.K.W.; Lu, J.Z.; Boado, R.J.; Pardridge, W.M. Intravenous treatment of experimental Parkinson’s disease in the mouse with an IgG-GDNF fusion protein that penetrates the blood-brain barrier. Brain Res. 2010, 1352, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Ungerstedt, U.; Arbuthnott, G.W. Quantitative recording of rotational behavior in rats after 6-hydroxy-dopamine lesions of the nigrostriatal dopamine system. Brain Res. 1970, 24, 485–493. [Google Scholar] [CrossRef]
- Schallert, T.; Tillerson, J.L. Intervention Strategies for Degeneration of Dopamine Neurons in Parkinsonism: Optimizing Behavioral Assessment of Outcome. In Central Nervous System Diseases; Emerich, D.F., Dean, R.L., Sanberg, P.R., Eds.; Humana Press: Totowa, NJ, USA, 2000; pp. 131–151. ISBN 978-1-59259-691-1. [Google Scholar]
- Schackel, S.; Pauly, M.-C.; Piroth, T.; Nikkhah, G.; Döbrössy, M.D. Donor age dependent graft development and recovery in a rat model of Huntington’s disease: Histological and behavioral analysis. Behav. Brain Res. 2013, 256, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Chou, A.H.; Chen, C.Y.; Chen, S.Y.; Chen, W.J.; Chen, Y.L.; Weng, Y.S.; Wang, H.L. Polyglutamine-expanded ataxin-7 causes cerebellar dysfunction by inducing transcriptional dysregulation. Neurochem. Int. 2010, 56, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Zilles, K.; Qü, M.S.; Schröder, H.; Schleicher, A. Neurotransmitter receptors and cortical architecture. J. Hirnforsch. 1991, 32, 343–356. [Google Scholar] [PubMed]
- Zilles, K.; Werner, L.; Qü, M.; Schleicher, A.; Gross, G. Quantitative auto radiography of 11 different transmitter binding sites in the basal forebrain region of the rat-evidence of heterogeneity in distribution patterns. Neuroscience 1991, 42, 473–481. [Google Scholar] [CrossRef]
- Zilles, K.; Gross, G.; Schleicher, A.; Schildgen, S.; Bauer, A.; Bahro, M.; Schwendemann, G.; Zech, K.; Kolassa, N. Regional and laminar distributions of alpha 1-adrenoceptors and their subtypes in human and rat hippocampus. Neuroscience 1991, 40, 307–320. [Google Scholar] [CrossRef]
- Zilles, K.; Palomero-Gallagher, N.; Grefkes, C.; Scheperjans, F.; Boy, C.; Amunts, K.; Schleicher, A. Architectonics of the human cerebral cortex and transmitter receptor fingerprints: Reconciling functional neuroanatomy and neurochemistry. Eur. Neuropsychopharmacol. 2002, 12, 587–599. [Google Scholar] [CrossRef]
- Zilles, K.; Schleicher, A.; Palomero-Gallagher, N.; Amunts, K. Quantitative Analysis of Cyto- and Receptor Architecture of the Human Brain. In Brain Mapping: The Methods; Toga, A.W., Mazziotta, J.C., Eds.; Elsevier Academic Press: London, UK, 2002; Volume 58, pp. 573–602. ISBN 9780126930191. 0385-5600 (Print) 0385-5600 (Linking). [Google Scholar]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antipova, V.; Wree, A.; Holzmann, C.; Mann, T.; Palomero-Gallagher, N.; Zilles, K.; Schmitt, O.; Hawlitschka, A. Unilateral Botulinum Neurotoxin-A Injection into the Striatum of C57BL/6 Mice Leads to a Different Motor Behavior Compared with Rats. Toxins 2018, 10, 295. https://doi.org/10.3390/toxins10070295
Antipova V, Wree A, Holzmann C, Mann T, Palomero-Gallagher N, Zilles K, Schmitt O, Hawlitschka A. Unilateral Botulinum Neurotoxin-A Injection into the Striatum of C57BL/6 Mice Leads to a Different Motor Behavior Compared with Rats. Toxins. 2018; 10(7):295. https://doi.org/10.3390/toxins10070295
Chicago/Turabian StyleAntipova, Veronica, Andreas Wree, Carsten Holzmann, Teresa Mann, Nicola Palomero-Gallagher, Karl Zilles, Oliver Schmitt, and Alexander Hawlitschka. 2018. "Unilateral Botulinum Neurotoxin-A Injection into the Striatum of C57BL/6 Mice Leads to a Different Motor Behavior Compared with Rats" Toxins 10, no. 7: 295. https://doi.org/10.3390/toxins10070295