The PHD Transcription Factor Rum1 Regulates Morphogenesis and Aflatoxin Biosynthesis in Aspergillus flavus

Abstract
:1. Introduction
2. Results
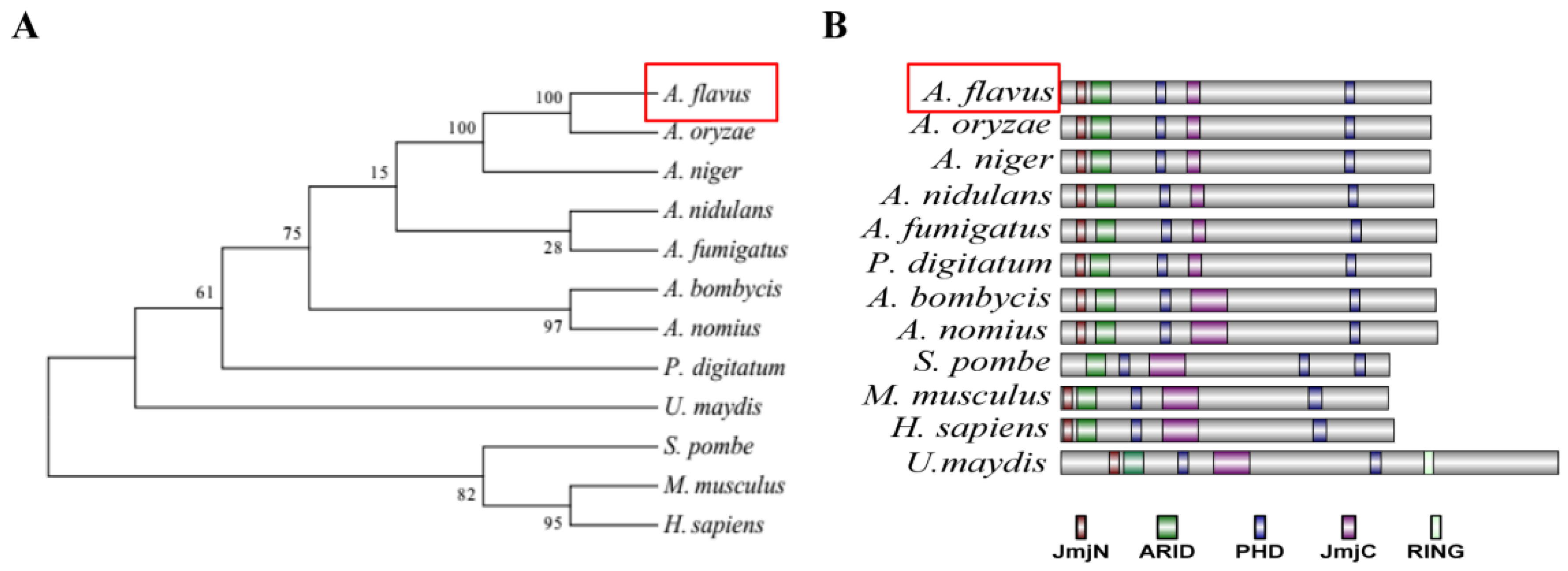
2.1. Characterization of the PHD Transcription Factor Rum1 in A. flavus
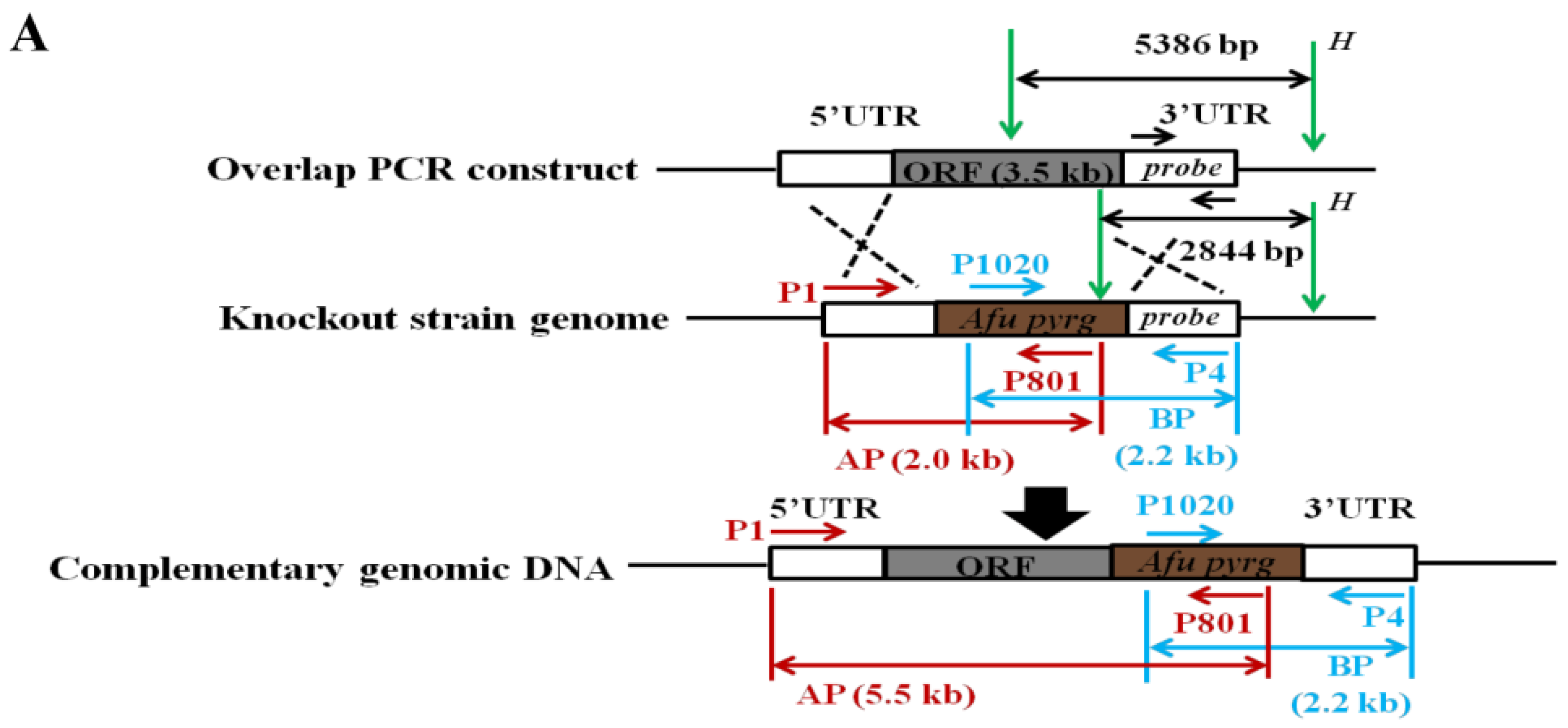
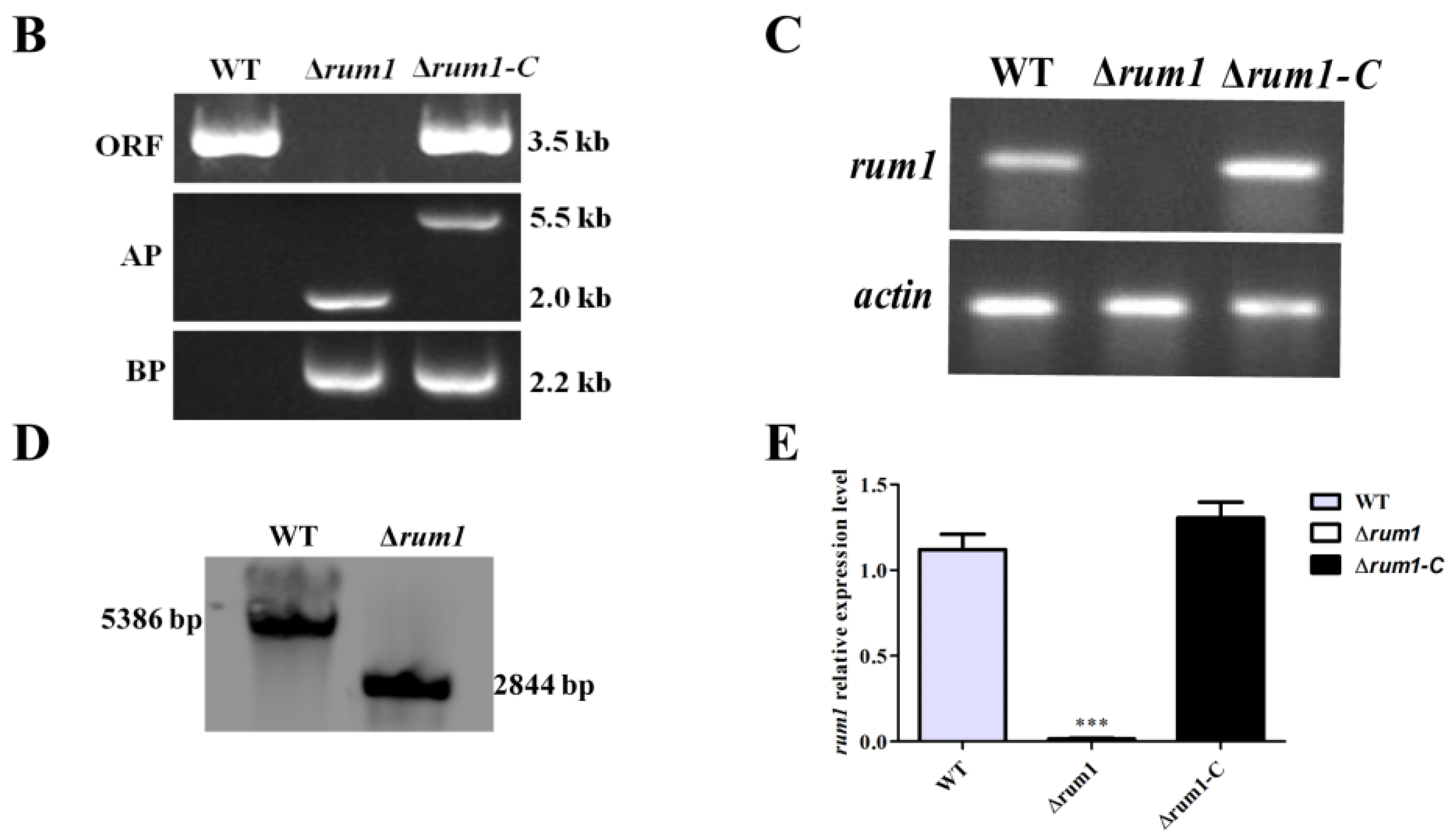
2.2. Construction of the Deleted Mutant (Δrum1) and Complemented Mutant (Δrum1-C)
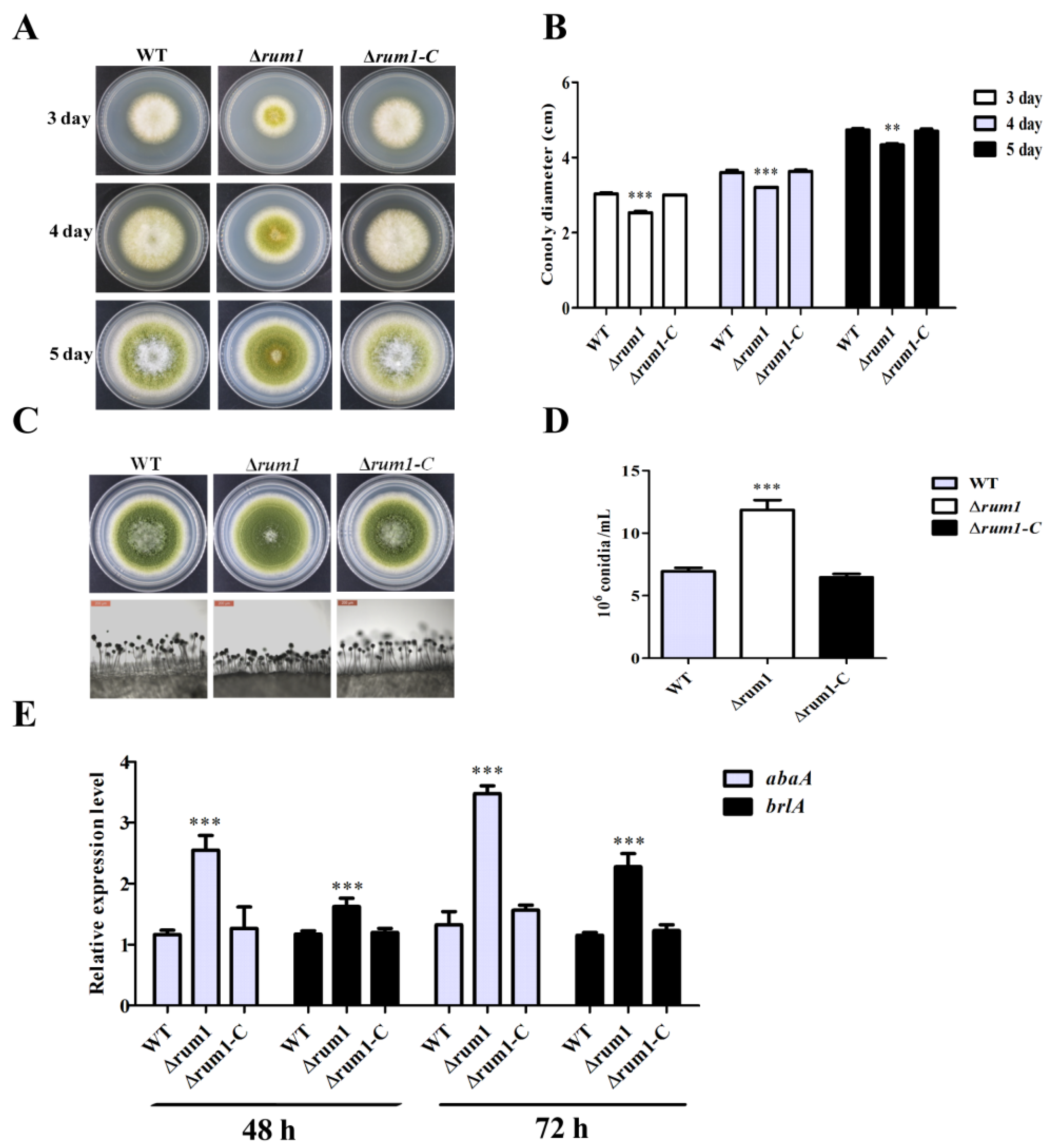
2.3. Rum1 Is Involved in Mycelium Growth and Conidiation
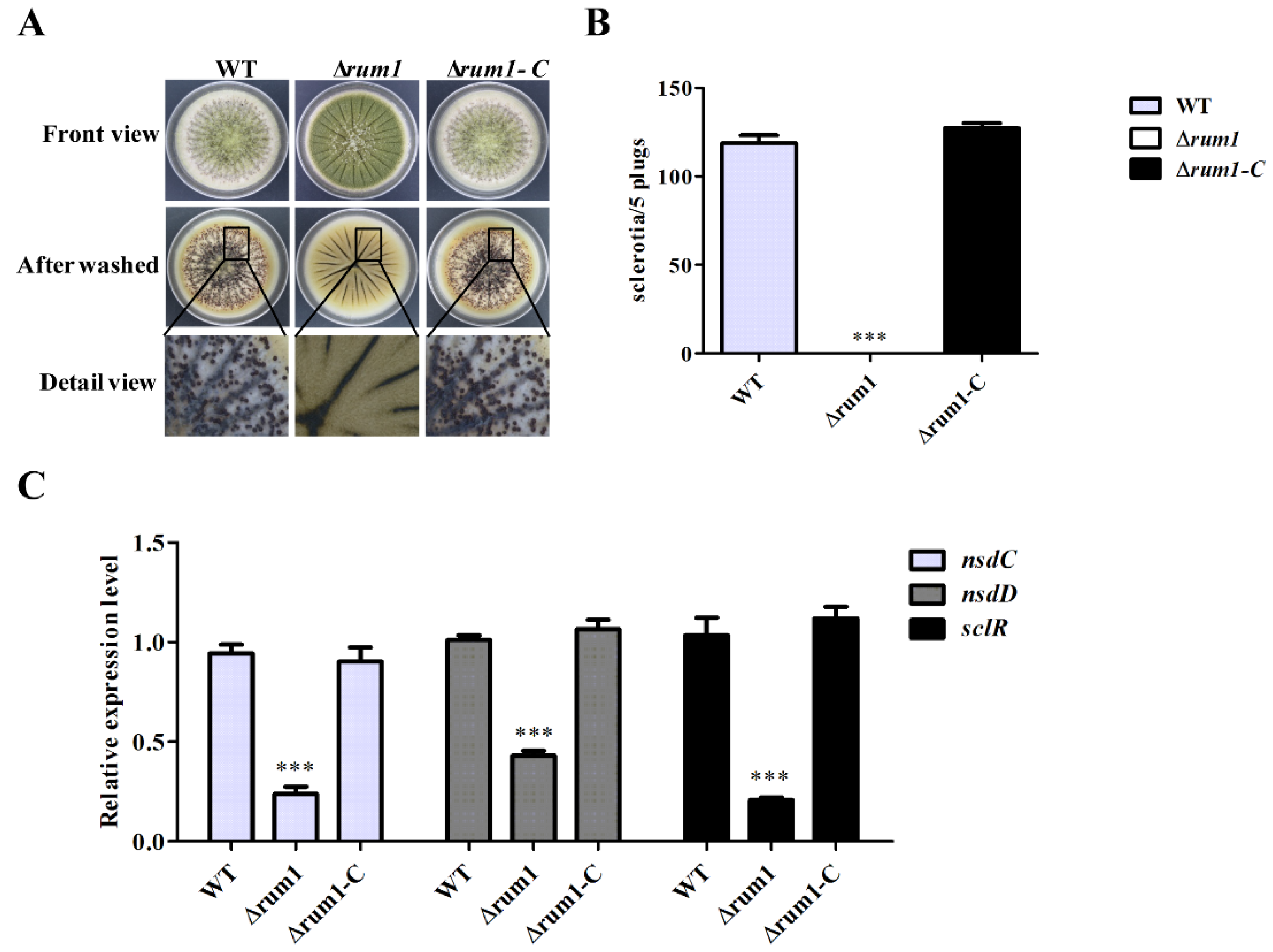
2.4. Rum1 Is Essential for Sclerotial Generation
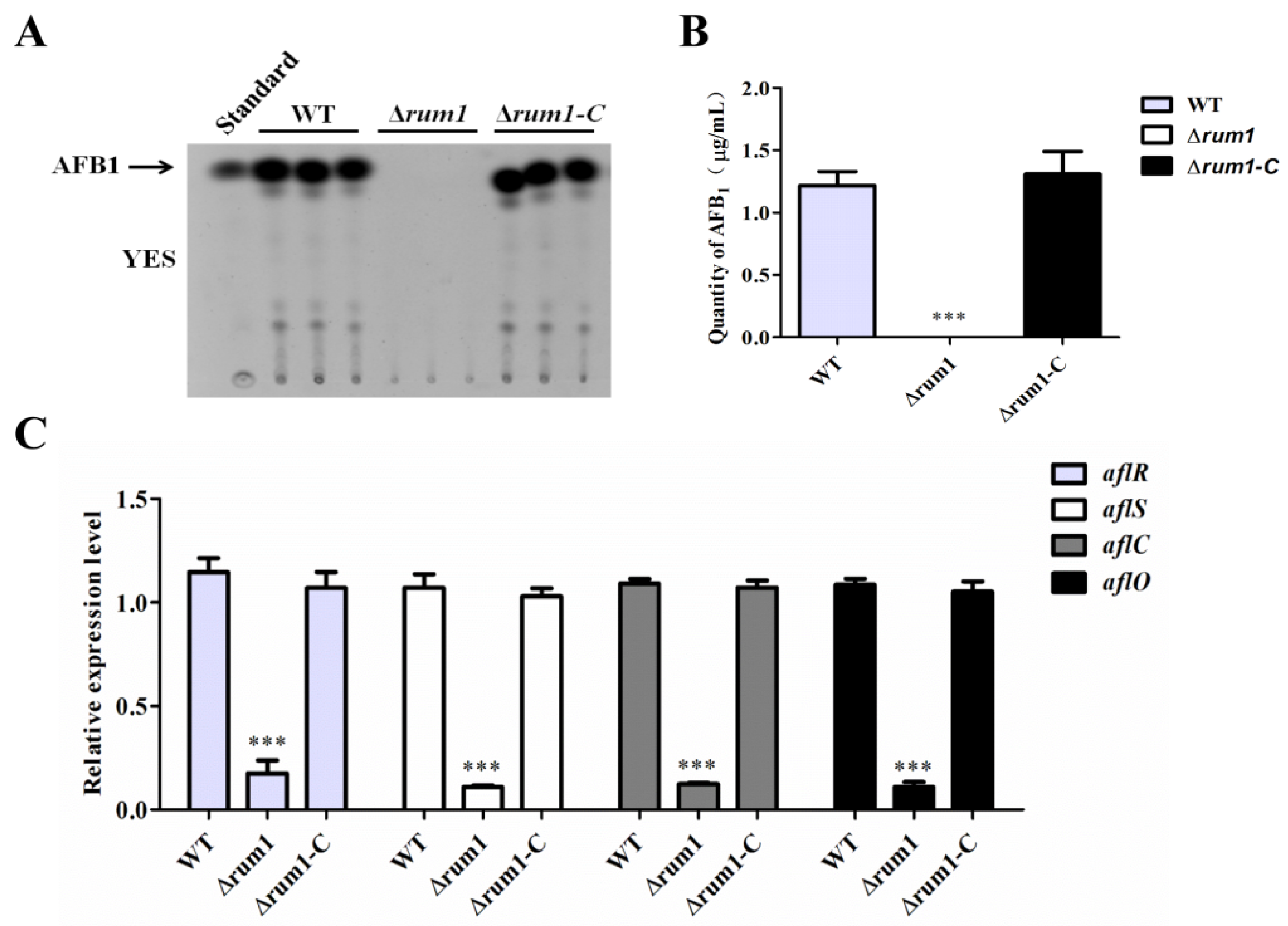
2.5. Rum1 Positively Regulates Aflatoxin Biosynthesis
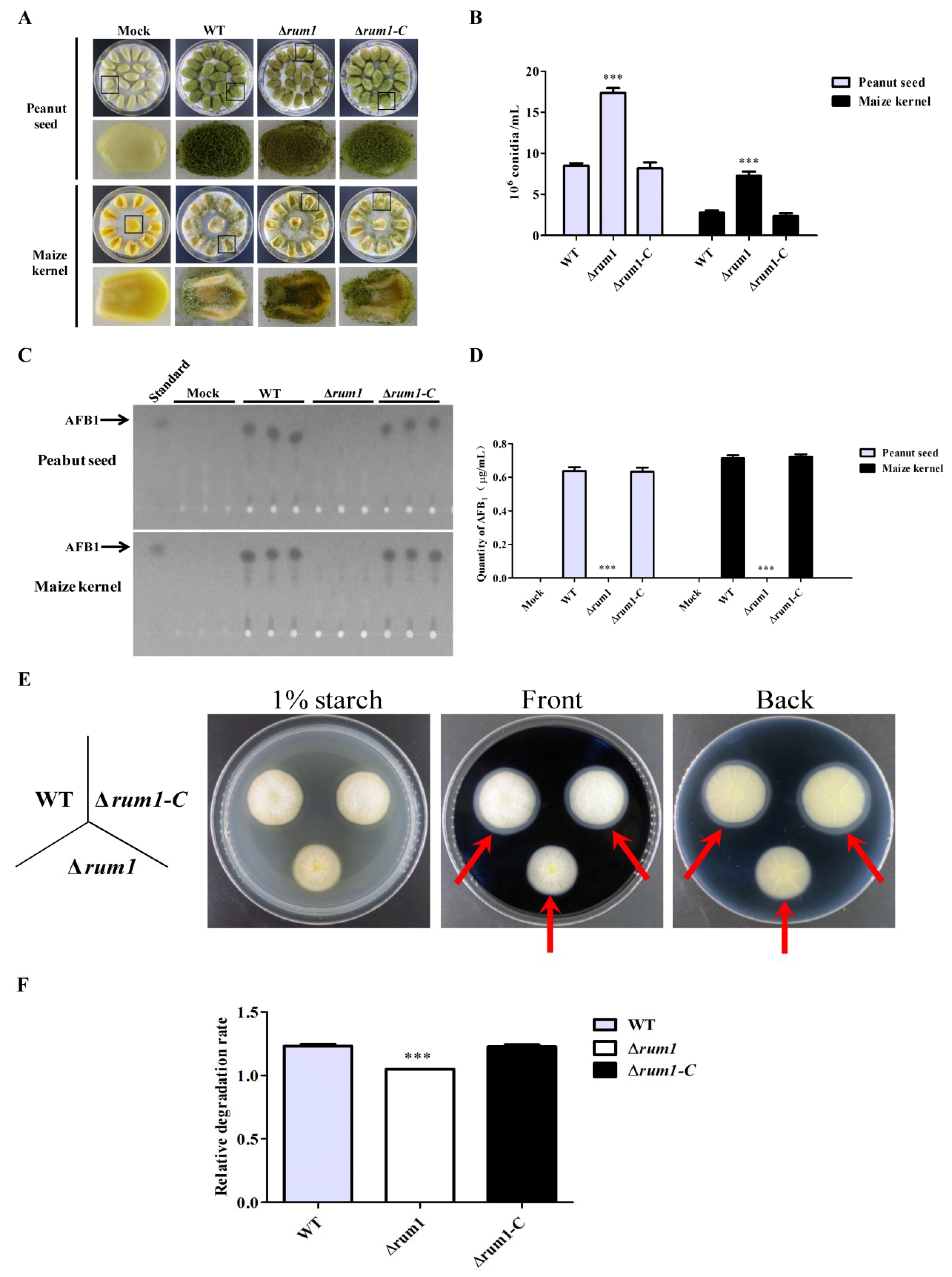
2.6. Rum1 is Involved in the Colonization of A. flavus to Crop Kernels
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fungal Strains and Primers
5.2. Phylogenetic Analysis
5.3. Mutant Strains Construction
5.4. Real-Time Quantitative Reverse Transcription PCR
5.5. Morphological Analysis
5.6. Aflatoxin Analysis
5.7. Stress Assays
5.8. Crop Kernels Colonization Assays
5.9. The Analysis on the Activity of Amylase
5.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hedayati, M.T.; Pasqualotto, A.C.; Warn, P.A.; Bowyer, P.; Denning, D.W. Aspergillus flavus: Human pathogen, allergen and mycotoxin producer. Microbiology 2007, 153, 1677–1692. [Google Scholar] [CrossRef] [PubMed]
- Xing, F.; Ding, N.; Liu, X.; Selvaraj, J.N.; Wang, L.; Zhou, L.; Zhao, Y.; Wang, Y.; Liu, Y. Variation in fungal microbiome (mycobiome) and aflatoxins during simulated storage of in-shell peanuts and peanut kernels. Sci. Rep. 2016, 6, 25930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohd Redzwan, S.; Rosita, J.; Mohd Sokhini, A.M.; Nurul Aqilah, A.R.; Wang, J.S.; Kang, M.S.; Zuraini, A. Detection of serum AFB1-lysine adduct in Malaysia and its association with liver and kidney functions. Int. J. Hyg. Environ. Health 2014, 217, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Yu, J. Current Understanding on Aflatoxin Biosynthesis and Future Perspective in Reducing Aflatoxin Contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misihairabgwi, J.M.; Ezekiel, C.N.; Sulyok, M.; Shephard, G.S.; Krska, R. Mycotoxin contamination of foods in Southern Africa: A 10-year review (2007–2016). Crit. Rev. Food Sci. Nutr. 2017, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Ann. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Ehrlich, K.C. Aflatoxigenicity in Aspergillus: Molecular genetics, phylogenetic relationships and evolutionary implications. Mycopathologia 2006, 162, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Kale, S.P.; Milde, L.; Trapp, M.K.; Frisvad, J.C.; Keller, N.P.; Bok, J.W. Requirement of LaeA for secondary metabolism and sclerotial production in Aspergillus flavus. Fungal Genet. Biol. 2008, 45, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, K.C.; Mack, B.M.; Wei, Q.; Li, P.; Roze, L.V.; Dazzo, F.; Cary, J.W.; Bhatnagar, D.; Linz, J.E. Association with AflR in endosomes reveals new functions for AflJ in aflatoxin biosynthesis. Toxins 2012, 4, 1582–1600. [Google Scholar] [CrossRef] [PubMed]
- Masanga, J.O.; Matheka, J.M.; Omer, R.A.; Ommeh, S.C.; Monda, E.O.; Alakonya, A.E. Downregulation of transcription factor aflR in Aspergillus flavus confers reduction to aflatoxin accumulation in transgenic maize with alteration of host plant architecture. Plant Cell Rep. 2015, 34, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Lohmar, J.M.; Satterlee, T.; Cary, J.W.; Calvo, A.M. The Master Transcription Factor mtfA Governs Aflatoxin Production, Morphological Development and Pathogenicity in the Fungus Aspergillus flavus. Toxins 2016, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Fountain, J.C.; Bajaj, P.; Pandey, M.; Nayak, S.N.; Yang, L.; Kumar, V.; Jayale, A.S.; Chitikineni, A.; Zhuang, W.; Scully, B.T.; et al. Oxidative stress and carbon metabolism influence Aspergillus flavus transcriptome composition and secondary metabolite production. Sci. Rep. 2016, 6, 38747. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Han, X.; Bai, Y.; Lin, Z.; Qiu, M.; Nie, X.; Wang, S.; Zhang, F.; Zhuang, Z.; Yuan, J.; et al. Effects of nitrogen metabolism on growth and aflatoxin biosynthesis in Aspergillus flavus. J. Hazard Mater. 2017, 324, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, K.C.; Chang, P.K.; Yu, J.; Cary, J.W.; Bhatnagar, D. Control of Aflatoxin Biosynthesis in Aspergilli. In Aflatoxins—Biochemistry and Molecular Biology; In Tech: New Orleans, LA, USA, 2011. [Google Scholar]
- Schindler, U.; Beckmann, H.; Cashmore, A.R. HAT3.1, a novel Arabidopsis homeodomain protein containing a conserved cysteine-rich region. Plant J. 1993, 4, 137–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aasland, R.; Gibson, T.J.; Stewart, A.F. The PHD finger: Implications for chromatin-mediated transcriptional regulation. Trends Biochem. Sci. 1995, 20, 56–59. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, J.; Wang, Y.; Zhao, Y.; Jiang, H.; Cheng, B. Systematic Analysis of the Maize PHD-Finger Gene Family Reveals a Subfamily Involved in Abiotic Stress Response. Int. J. Mol. Sci. 2015, 16, 23517–23544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, T.V.; Kaur, J.; Agashe, B.; Sundaresan, V.; Siddiqi, I. The duet gene is necessary for chromosome organization and progression during male meiosis in arabidopsis and encodes a PHD finger protein. Development 2003, 130, 5975–5987. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gao, Q.; Li, P.; Zhao, Q.; Zhang, J.; Li, J.; Koseki, H.; Wong, J. UHRF1 targets DNMT1 for DNA methylation through cooperative binding of hemi-methylated DNA and methylated H3K9. Nat. Commun. 2013, 4, 1563. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.; Alvarez-Venegas, R.; Yilmaz, M.; Le, O.; Hou, G.; Sadder, M.; Al-Abdallat, A.; Xia, Y.; Lu, G.; Ladunga, I. The highly similar arabidopsis homologs of trithorax ATX1 and ATX2 encode proteins with divergent biochemical functions. Plant Cell 2008, 20, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Merkel, D.J.; Wells, S.B.; Hilburn, B.C.; Elazzouzi, F.; Pérez-Alvarado, G.C.; Lee, B.M. The C-terminal region of cytoplasmic polyadenylation element binding protein is a ZZ domain with potential for protein–protein interactions. J. Mol. Biol. 2013, 425, 2015–2026. [Google Scholar] [CrossRef] [PubMed]
- Quadbeck-Seeger, C.; Wanner, G.; Huber, S.; Kahmann, R.; Kamper, J. A protein with similarity to the human retinoblastoma binding protein 2 acts specifically as a repressor for genes regulated by the b mating type locus in Ustilago maydis. Mol. Microbiol. 2000, 38, 154–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, S.J.; Stelzer, C.P.; Welch, D.B.; Logsdon, J.M., Jr. Comparative transcriptome analysis of obligately asexual and cyclically sexual rotifers reveals genes with putative functions in sexual reproduction, dormancy, and asexual egg production. BMC Genom. 2013, 14, 412. [Google Scholar] [CrossRef] [PubMed]
- Yabe, K.; Nakajima, H. Enzyme reactions and genes in aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 2004, 64, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Park, J.W.; Chun, Y.S. Jumonji histone demethylases as emerging therapeutic targets. Pharmacol. Res. 2016, 105, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Shmakova, A.; Batie, M.; Druker, J.; Rocha, S. Chromatin and oxygen sensing in the context of JmjC histone demethylases. Biochem. J. 2014, 462, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slama, P. Two-domain analysis of JmjN-JmjC and PHD-JmjC lysine demethylases: Detecting an inter-domain evolutionary stress. Proteins 2018, 86, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Quan, Z.; Oliver, S.G.; Zhang, N. JmjN interacts with JmjC to ensure selective proteolysis of Gis1 by the proteasome. Microbiology 2011, 157, 2694–2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, F.T.; Madsen, C.K.; Nordland, A.M.; Grasser, M.; Merkle, T.; Grasser, K.D. A novel family of plant DNA-binding proteins containing both HMG-box and AT-rich interaction domains. Biochemistry 2008, 47, 13207–13214. [Google Scholar] [CrossRef] [PubMed]
- Wilsker, D.; Patsialou, A.; Dallas, P.B.; Moran, E. ARID proteins: A diverse family of DNA binding proteins implicated in the control of cell growth, differentiation, and development. Cell Growth Differ. 2002, 13, 95–106. [Google Scholar] [PubMed]
- Wang, X.; Nagl, N.G.; Wilsker, D.; Van Scoy, M.; Pacchione, S.; Yaciuk, P.; Dallas, P.B.; Moran, E. Two related ARID family proteins are alternative subunits of human SWI/SNF complexes. Biochem. J. 2004, 383, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, R.; Zhou, M.M. The PHD finger: A versatile epigenome reader. Trends Biochem. Sci. 2011, 36, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Yu, J.H. Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 2012, 15, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Yamada, O.; Lee, B.R.; Gomi, K.; Iimura, Y. Cloning and functional analysis of the Aspergillus oryzae conidiation regulator gene brlA by its disruption and misscheduled expression. J. Biosci. Bioeng. 1999, 87, 424–429. [Google Scholar] [CrossRef]
- Adams, T.H.; Boylan, M.T.; Timberlake, W.E. BrlA is necessary and sufficient to direct conidiophore development in Aspergillus nidulans. Cell 1988, 54, 353–362. [Google Scholar] [CrossRef]
- Han, S.; Adams, T.H. Complex control of the developmental regulatory locus brlA in Aspergillus nidulans. Mol. Genet. Genom. 2001, 266, 260–270. [Google Scholar]
- Jin, F.J.; Takahashi, T.; Matsushima, K.; Hara, S.; Shinohara, Y.; Maruyama, J.; Kitamoto, K.; Koyama, Y. SclR, a basic helix-loop-helix transcription factor, regulates hyphal morphology and promotes sclerotial formation in Aspergillus oryzae. Eukaryot. Cell 2011, 10, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Kwon, N.J.; Choi, J.M.; Lee, I.S.; Jung, S.; Yu, J.H. NsdD is a key repressor of asexual development in Aspergillus nidulans. Genetics 2014, 197, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Han, Z.; Yin, Y.; Lohmar, J.M.; Shantappa, S.; Harris-Coward, P.Y.; Mack, B.; Ehrlich, K.C.; Wei, Q.; Arroyo-Manzanares, N.; et al. Transcriptome Analysis of Aspergillus flavus Reveals veA-Dependent Regulation of Secondary Metabolite Gene Clusters, Including the Novel Aflavarin Cluster. Eukaryot. Cell 2015, 14, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Butchko, R.A.; Fernandes, M.; Keller, N.P.; Leonard, T.J.; Adams, T.H. Conservation of structure and function of the aflatoxin regulatory gene aflR from Aspergillus nidulans and A. flavus. Curr. Genet. 1996, 29, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Liang, L.; Ran, F.; Liu, Y.; Li, Z.; Lan, H.; Gao, P.; Zhuang, Z.; Zhang, F.; Nie, X.; et al. The DmtA methyltransferase contributes to Aspergillus flavus conidiation, sclerotial production, aflatoxin biosynthesis and virulence. Sci. Rep. 2016, 6, 23259. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Liu, Y.; Liang, L.; Li, Z.; Qin, Q.; Nie, X.; Wang, S. The high-affinity phosphodiesterase PdeH regulates development and aflatoxin biosynthesis in Aspergillus flavus. Fungal Genet. Biol. 2017, 101, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K.; Scharfenstein, L.L.; Wei, Q.; Bhatnagar, D. Development and refinement of a high-efficiency gene-targeting system for Aspergillus flavus. J. Microbiol. Methods 2010, 81, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Yu, S.; Qiu, M.; Wang, X.; Wang, Y.; Bai, Y.; Zhang, F.; Wang, S. Aspergillus flavus SUMO Contributes to Fungal Virulence and Toxin Attributes. J. Agric. Food Chem. 2016, 64, 6772–6782. [Google Scholar] [CrossRef] [PubMed]
- Lan, H.; Sun, R.; Fan, K.; Yang, K.; Zhang, F.; Nie, X.Y.; Wang, X.; Zhuang, Z.; Wang, S. The Aspergillus flavus Histone Acetyltransferase AflGcnE Regulates Morphogenesis, Aflatoxin Biosynthesis, and Pathogenicity. Front. Microbiol. 2016, 7, 1324. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, Y.; Li, X.; Fasoyin, O.E.; Hu, Y.; Liu, Y.; Yuan, J.; Zhuang, Z.; Wang, S. Histone Methyltransferase aflrmtA gene is involved in the morphogenesis, mycotoxin biosynthesis, and pathogenicity of Aspergillus flavus. Toxicon 2017, 127, 112–121. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype Description | Reference |
|---|---|---|
| A. flavus CA14 | ∆ku70, ∆pyrG | purchased from FGSC |
| wild-type (WT) | ∆ku70, ∆pyrG::AfpyrG | This study |
| ∆rum1 | ∆ku70, ∆rum1::AfpyrG | This study |
| ∆rum1-C | ∆ku70, ∆rum1::AfpyrG, rum1::AfpyrG | This study |
| Primer Name | Sequence (5′-3′) | Fragment |
|---|---|---|
| rum1-p1 | GGCACGAGCTATTAGTGATATTAGTCGAGTCCGA | 5′UTR of rum1 |
| rum1-p2 | CAAGTGAGCCGACCGATTGAGGGAAGTAGT | |
| rum1-p3 | TCCCTATCAACAAATTGGCGCTTCATGGGTTC | 3′UTR of rum1 |
| rum1-p4 | TGGATTCCTTCGGGGGCTAGTTTGCATC | |
| rum1-p5 | ACTACTTCCCTCAATCGGTCGGCTCACTT | A. fumigatus pyrG |
| GGCCTCAAACAATGCTCTTCACCC | ||
| rum1-p6 | GAACCCATGAAGCGCCAATTTGTTGATA | |
| GGGAGTCTGAGAGGAGGCACTGATGC | ||
| rum1-p7 | GACCTGTGAAGATGCTTGGTAGAGCTATTTCAG | Nesting primers |
| rum1-p8 | TATCTCATTGGACTGGACCCTGAGCGGGA | |
| rum1-p9 | CAACTCGACTGGCGGACAGCCT | A fragment from rum1 |
| rum1-p10 | TCATTTGCCGGAGAATATGTTCCAGTCCTTC | |
| P801 | CAGGAGTTCTCGGGTTGTCG | A. fumigatus pyrG |
| P1020 | CAGAGTATGCGGCAAGTCA | |
| rum-p3 + pyrg-F | CTTCATCGCGAGATAACACCCCCGATGG | 5′UTR of Δrum1-C |
| rum-p3 + pyrg-R | GGGTGAAGAGCATTGTTTGAGGCCCCATG | |
| pyrg-F | GCCTCAAACAATGCTCTTCACCC | A. fumigatus pyrG |
| pyrg-R | GTCTGAGAGGAGGCACTGATGC | |
| rum-p4 + pyrg-F | GCATCAGTGCCTCCTCTCAGACAGATTCTT | 3′UTR of Δrum1-C |
| GCCTTGCGCATTCATGACAAC |
| Gene | Forward Sequences (5′-3′) | Reverse Sequences (3′-5′) |
|---|---|---|
| rum1 | CTTGATGCATCTCTCTTT | CTTCCAGAGCCTCATTA |
| AGCTCTCCACGGTTC | GCATGTGTGTTCTCC | |
| brlA | GCCTCCAGCGTCAACCTTC | TCTCTTCAAATGCTCTTGCCTC |
| abaA | TCTTCGGTTGATGGATGATTTC | CCGTTGGGAGGCTGGGT |
| nsdC | GCCAGACTTGCCAATCAC | CATCCACCTTGCCCTTTA |
| nsdD | GGACTTGCGGGTCGTGCTA | AGAACGCTGGGTCTGGTGC |
| sclR | CAATGAGCCTATGGGAGTGG | ATCTTCGCCCGAGTGGTT |
| aflR | AAAGCACCCTGTCTTCCCTAAC | GAAGAGGTGGGTCAGTGTTTGTAG |
| aflS | CGAGTCGCTCAGGCGCTCAA | GCTCAGACTGACCGCCGCTC |
| aflC | GTGGTGGTTGCCAATGCG | CTGAAACAGTAGGACGGGAGC |
| aflO | GATTGGGATGTGGTCATGCGATT | GCCTGGGTCCGAAGAATGC |
| actin | ACGGTGTCGTCACAAACTGG | CGGTTGGACTTAGGGTTGATAG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Yang, G.; Zhang, D.; Liu, Y.; Li, Y.; Lin, G.; Guo, Z.; Wang, S.; Zhuang, Z. The PHD Transcription Factor Rum1 Regulates Morphogenesis and Aflatoxin Biosynthesis in Aspergillus flavus. Toxins 2018, 10, 301. https://doi.org/10.3390/toxins10070301
Hu Y, Yang G, Zhang D, Liu Y, Li Y, Lin G, Guo Z, Wang S, Zhuang Z. The PHD Transcription Factor Rum1 Regulates Morphogenesis and Aflatoxin Biosynthesis in Aspergillus flavus. Toxins. 2018; 10(7):301. https://doi.org/10.3390/toxins10070301
Chicago/Turabian StyleHu, Yule, Guang Yang, Danping Zhang, Yaju Liu, Yu Li, Guanglan Lin, Zhiqiang Guo, Shihua Wang, and Zhenhong Zhuang. 2018. "The PHD Transcription Factor Rum1 Regulates Morphogenesis and Aflatoxin Biosynthesis in Aspergillus flavus" Toxins 10, no. 7: 301. https://doi.org/10.3390/toxins10070301