Species Composition and Trichothecene Genotype Profiling of Fusarium Field Isolates Recovered from Wheat in Poland

 , , ,
, , ,
Abstract
:1. Introduction
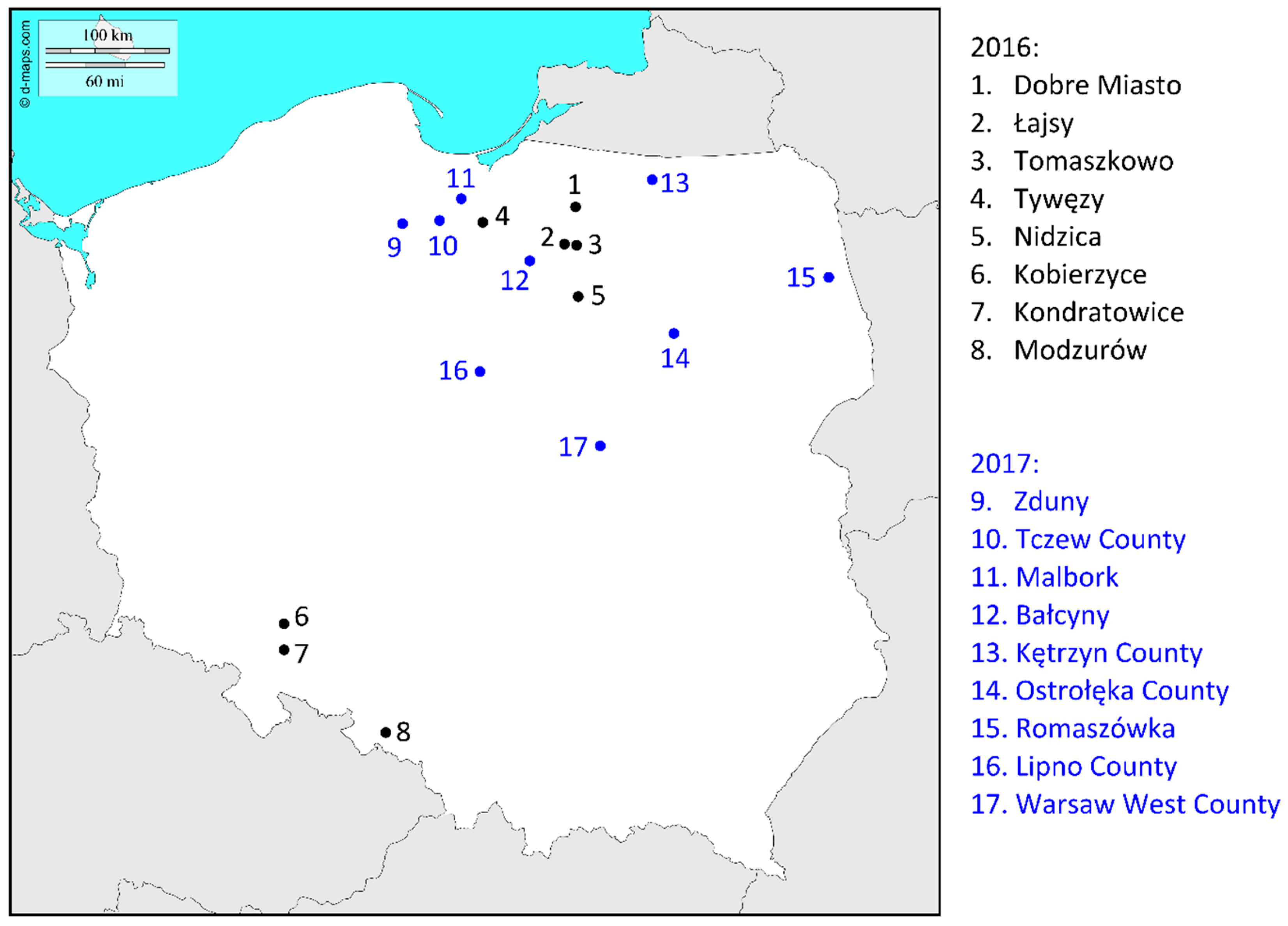
2. Results
2.1. Determination of Fungal Species and Trichothecene Genotypes
2.2. Comparative Analysis of the Tri Core Cluster
3. Discussion
4. Materials and Methods
4.1. Fusarium-Damaged Kernels (FDKs)
4.2. Preparation of Cell Lysates for qPCR
4.3. Determination of Fungal Species and Tri Genotypes
4.4. DNA Sequencing, Assembly, and Annotation of Tri Core Clusters
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of wheat diseases—A field perspective. Mol. Plant Pathol. 2018, 19, 1523–1536. [Google Scholar] [CrossRef] [PubMed]
- McMullen, M.; Jones, R.; Gallenberg, D. Scab of Wheat and Barley: A Re-emerging Disease of Devastating Impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar] [CrossRef]
- Doohan, F.M.; Brennan, J.M.; Cooke, B.M. Influence of climatic factors on Fusarium species pathogenic to cereals. Eur. J. Plant Pathol. 2003, 109, 755–768. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Baenziger, P.S.; Hernandez Nopsa, J.; Bockus, W.; Hallen-Adams, H. Management of Fusarium head blight of wheat and barley. Crop Prot. 2015, 73, 100–107. [Google Scholar] [CrossRef]
- Desjardins, A.E. Fusarium Mycotoxins Chemistry, Genetics and Biology; American Phytopathological Society Press: St. Paul, MN, USA, 2006. [Google Scholar]
- Basler, R. Diversity of Fusarium species isolated from UK forage maize and the population structure of F. graminearum from maize and wheat. PeerJ 2016, 4, e2143. [Google Scholar] [CrossRef] [PubMed]
- Klix, M.B.; Verreet, J.A.; Beyer, M. Comparison of the declining triazole sensitivity of Gibberella zeae and increased sensitivity achieved by advances in triazole fungicide development. Crop Prot. 2007, 26, 683–690. [Google Scholar] [CrossRef]
- Yin, Y.; Liu, X.; Li, B.; Ma, Z. Characterization of sterol demethylation inhibitor-resistant isolates of Fusarium asiaticum and F. graminearum collected from wheat in China. Phytopathology 2009, 99, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Becher, R.; Hettwer, U.; Karlovsky, P.; Deising, H.B.; Wirsel, S.G.R. Adaptation of Fusarium graminearum to tebuconazole yielded descendants diverging for levels of fitness, fungicide resistance, virulence, and mycotoxin production. Phytopathology 2010, 100, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Steiner, B.; Buerstmayr, M.; Michel, S.; Schweiger, W.; Lemmens, M.; Buerstmayr, H. Breeding strategies and advances in line selection for Fusarium head blight resistance in wheat. Trop. Plant Pathol. 2017, 42, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Ward, T.J.; Geiser, D.M.; Corby Kistler, H.; Aoki, T. Genealogical concordance between the mating type locus and seven other nuclear genes supports formal recognition of nine phylogenetically distinct species within the Fusarium graminearum clade. Fungal Genet. Biol. 2004, 41, 600–623. [Google Scholar] [CrossRef] [PubMed]
- Starkey, D.E.; Ward, T.J.; Aoki, T.; Gale, L.R.; Kistler, H.C.; Geiser, D.M.; Suga, H.; Tóth, B.; Varga, J.; O’Donnell, K. Global molecular surveillance reveals novel Fusarium head blight species and trichothecene toxin diversity. Fungal Genet. Biol. 2007, 44, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Sarver, B.A.; Ward, T.J.; Gale, L.R.; Broz, K.; Kistler, H.C.; Aoki, T. Novel Fusarium head blight pathogens from Nepal and Louisiana revealed by multilocus genealogical concordance. Fungal Genet. Biol. 2011, 48, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Van der Lee, T.; Waalwijk, C.; Chen, W.; Xu, J.; Xu, J.; Zhang, Y.; Feng, J. Population analysis of the Fusarium graminearum species complex from wheat in China show a shift to more aggressive isolates. PLoS ONE 2012, 7, e31722. [Google Scholar] [CrossRef] [PubMed]
- Ward, T.J.; Clear, R.M.; Rooney, A.P.; O’Donnell, K.; Gaba, D.; Patrick, S.; Starkey, D.E.; Gilbert, J.; Geiser, D.M.; Nowicki, T.W. An adaptive evolutionary shift in Fusarium head blight pathogen populations is driving the rapid spread of more toxigenic Fusarium graminearum in North America. Fungal Genet. Biol. 2008, 45, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Scoz, L.B.; Astolfi, P.; Reartes, D.S.; Schmale III, D.G.; Moraes, M.G.; Del Ponte, E.M. Trichothecene mycotoxin genotypes of Fusarium graminearum sensu stricto and Fusarium meridionale in wheat from southern Brazil. Plant Pathol. 2009, 58, 344–351. [Google Scholar] [CrossRef]
- Reynoso, M.M.; Ramirez, M.L.; Torres, A.M.; Chulze, S.N. Trichothecene genotypes and chemotypes in Fusarium graminearum strains isolated from wheat in Argentina. Int. J. Food Microbiol. 2011, 145, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Castañares, E.; Albuquerque, D.R.; Dinolfo, M.I.; Pinto, V.F.; Patriarca, A.; Stenglein, S.A. Trichothecene genotypes and production profiles of Fusarium graminearum isolates obtained from barley cultivated in Argentina. Int. J. Food Microbiol. 2014, 179, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Suga, H.; Karugia, G.W.; Ward, T.J.; Gale, L.R.; Tomimura, K.; Nakajima, T.; Miyasaka, A.; Koizumi, S.; Kageyama, K.; Hyakumachi, M. Molecular characterization of the Fusarium graminearum species complex in Japan. Phytopathology 2008, 98, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; van der Lee, T.; Yang, X.; Yu, D.; Waalwijk, C. Fusarium populations on Chinese barley show a dramatic gradient in mycotoxin profiles. Phytopathology 2008, 98, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, Z.; van der Lee, T.; Chen, W.Q.; Xu, J.; Xu, J.S.; Yang, L.; Yu, D.; Waalwijk, C.; Feng, J. Population genetic analyses of Fusarium asiaticum populations from barley suggest a recent shift favoring 3ADON producers in southern China. Phytopathology 2010, 100, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Van der Lee, T.; Zhang, H.; van Diepeningen, A.; Waalwijk, C. Biogeography of Fusarium graminearum species complex and chemotypes: A review. Food Addit. Contam. Part A 2015, 32, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q.L. Fusarium culmorum: Causal agent of foot and root rot and head blight on wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Wakuliński, W.; Chełkowski, J. Fusarium species transmitted with seeds of wheat, rye, barley, oats and triticale. Hodowla Roślin Aklimatyzacja Nasiennictwo 1993, 37, 131–136. [Google Scholar]
- Chełkowski, J. Distribution of Fusarium species and their mycotoxins in cereal grains. In Mycotoxins in Agriculture and Food Safety; Sinha, K.K., Bhatnagar, D., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1998; pp. 45–64. [Google Scholar]
- Tomczak, M.; Wiśniewska, H.; Stcępień, Ł.; Kostecki, M.; Chełkowski, J.; Goliński, P. Deoxynivalenol, nivalenol and moniliformin in wheat samples with head blight (scab) symptoms in Poland (1998–2000). Eur. J. Plant Pathol. 2002, 108, 625–630. [Google Scholar] [CrossRef]
- Stępień, Ł.; Popiel, D.; Koczyk, G.; Chełkowski, J. Wheat-infecting Fusarium species in Poland-their chemotypes and frequencies revealed by PCR assay. J. Appl. Genet. 2008, 49, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewska, H.; Stępień, Ł.; Waśkiewicz, A.; Beszterda, M.; Góral, T.; Belter, J. Toxigenic Fusarium species infecting wheat heads in Poland. Cent. Eur. J. Biol. 2014, 9, 163–172. [Google Scholar] [CrossRef]
- Kuzdraliński, A.; Nowak, M.; Szczerba, H.; Dudziak, K.; Muszyńska, M.; Leśniowska-Nowak, J. The composition of Fusarium species in wheat husks and grains in south-eastern Poland. J. Integr. Agric. 2017, 16, 1530–1536. [Google Scholar] [CrossRef]
- Waalwijk, C.; Kastelein, P.; De Vries, I.; Kerényi, Z.; Van Der Lee, T.; Hesselink, T.; Köhl, J.; Kema, G. Major changes in Fusarium spp. in wheat in the Netherlands. Eur. J. Plant Pathol. 2003, 109, 743–754. [Google Scholar] [CrossRef]
- Giraud, F.; Pasquali, M.; Jarroudi, M.; Vrancken, C.; Brochot, C.; Cocco, E.; Hoffmann, L.; Delfosse, P.; Bohn, T. Fusarium head blight and associated mycotoxin occurrence on winter wheat in Luxembourg in 2007/2008. Food Addit. Contam. Part A 2010, 27, 825–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chełkowski, J.; Gromadzka, K.; Stępień, Ł.; Lenc, L.; Kostecki, M.; Berthiller, F. Fusarium species, zearalenone and deoxynivalenol content in preharvest scabby wheat heads from Poland. World Mycotoxin J. 2012, 5, 133–141. [Google Scholar] [CrossRef]
- Ward, T.J.; Bielawski, J.P.; Kistler, H.C.; Sullivan, E.; O’Donnell, K. Ancestral polymorphism and adaptive evolution in the trichothecene mycotoxin gene cluster of phytopathogenic Fusarium. Proc. Natl. Acad. Sci. USA 2002, 99, 9278–9283. [Google Scholar] [CrossRef] [PubMed]
- Pasquali, M.; Giraud, F.; Brochot, C.; Cocco, E.; Hoffmann, L.; Bohn, T. Genetic Fusarium chemotyping as a useful tool for predicting nivalenol contamination in winter wheat. Int. J. Food Microbiol. 2010, 137, 246–253. [Google Scholar] [CrossRef] [PubMed]
- De Kuppler, A.M.; Steiner, U.; Sulyok, M.; Krska, R.; Oerke, E.C. Genotyping and phenotyping of Fusarium graminearum isolates from Germany related to their mycotoxin biosynthesis. Int. J. Food. Microbiol. 2011, 151, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Puri, K.D.; Zhong, S. The 3ADON population of Fusarium graminearum found in North Dakota is more aggressive and produces a higher level of DON than the prevalent 15ADON population in spring wheat. Phytopathology 2010, 100, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Malihipour, A.; Gilbert, J.; Piercey-Normore, M.; Cloutier, S. Molecular phylogenetic analysis, trichothecene chemotype patterns, and variation in aggressiveness of Fusarium isolates causing head blight in wheat. Plant Dis. 2012, 96, 1016–1025. [Google Scholar] [CrossRef]
- Amarasinghe, C.C.; Tamburic-Ilincic, L.; Gilbert, J.; Brûlé-Babel, A.L.; Fernando, W.G.D. Evaluation of different fungicides for control of Fusarium head blight in wheat inoculated with 3ADON and 15ADON chemotypes of Fusarium graminearum in Canada. Can. J. Plant Pathol. 2013, 35, 200–208. [Google Scholar] [CrossRef]
- Gilbert, J.; Clear, R.M.; Ward, T.J.; Gaba, D.; Tekauz, A.; Turkington, T.K.; Woods, S.M.; Nowicki, T.; O’Donnell, K. Relative aggressiveness and production of 3- or 15-acetyl deoxynivalenol and deoxynivalenol by Fusarium graminearum in spring wheat. Can. J. Plant Pathol. 2010, 32, 146–152. [Google Scholar] [CrossRef]
- Von der Ohe, C.; Gauthier, V.; Tamburic-Ilincic, L.; Brule-Babel, A.; Fernando, W.G.D.; Clear, R.; Ward, T.J.; Miedaner, T. A comparison of aggressiveness and deoxynivalenol production between Canadian Fusarium graminearum isolates with 3-acetyl and 15-acetyldeoxynivalenol chemotypes in field-grown spring wheat. Eur. J. Plant Pathol. 2010, 127, 407–417. [Google Scholar] [CrossRef]
- Quarta, A.; Mita, G.; Haidukowski, M.; Santino, A.; Mulè, G.; Visconti, A. Assessment of trichothecene chemotypes of Fusarium culmorum occurring in Europe. Food Addit. Contam. 2005, 22, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Kulik, T. Development of TaqMan Assays for 3ADON, 15ADON and NIV Fusarium Genotypes Based on Tri12 Gene. Cereal Res. Commun. 2011, 39, 201–215. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Jensen, J.D.; Rodríguez, A.; Jørgensen, L.N.; Justesen, A.F. TRI12 based quantitative real-time PCR assays reveal the distribution of trichothecene genotypes of F. graminearum and F. culmorum isolates in Danish small grain cereals. Int. J. Food. Microbiol. 2012, 157, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Balmas, V.; Scherm, B.; Marcello, A.; Beyer, M.; Hoffmann, L.; Migheli, Q.; Pasquali, M. Fusarium species and chemotypes associated with Fusarium head blight and Fusarium root rot on wheat in Sardinia. Plant Pathol. 2015, 64, 972–979. [Google Scholar] [CrossRef]
- Burlakoti, R.R.; Neate, S.M.; Adhikari, T.B.; Gyawali, S.; Salas, B.; Steffenson, B.J.; Schwarz, P.B. Trichothecene profiling and population genetic analysis of Gibberella zeae from barley in North Dakota and Minnesota. Phytopathology 2011, 101, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Beyer, M.; Pogoda, F.; Pallez, M.; Lazic, J.; Hoffmann, L.; Pasquali, M. Evidence for a reversible drought induced shift in the species composition of mycotoxin producing Fusarium head blight pathogens isolated from symptomatic wheat heads. Int. J. Food Microbiol. 2014, 182, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Waalwijk, C.; de Vries, I.M.; Köhl, J.; Xu, X.; van der Lee, T.A.J.; Kema, G.H.J. Development of quantitative detection methods for Fusarium in cereals and their application. In Mycotoxins: Detection Methods, Management, Public Health and Agricultural Trade; Leslie, J., Bandyopadhyay, R., Visconti, A., Eds.; CABI Publishing: Wallingford, UK, 2008; pp. 197–207. [Google Scholar]
- Boutigny, A.L.; Ward, T.J.; Ballois, N.; Iancu, G.; Ioos, R. Diversity of the Fusarium graminearum species complex on French cereals. Eur. J. Plant Pathol. 2014, 138, 133–148. [Google Scholar] [CrossRef]
- Talas, F.; Parzies, H.K.; Miedaner, T. Diversity in genetic structure and chemotype composition of Fusarium graminearum sensu stricto populations causing wheat head blight in individual fields in Germany. Eur. J. Plant Pathol. 2011, 1, 39–48. [Google Scholar] [CrossRef]
- Prodi, A.; Purahong, W.; Tonti, S.; Salomoni, D.; Nipoti, P.; Covarelli, L.; Pisi, A. Difference in chemotype composition of Fusarium graminearum populations isolated from durum wheat in adjacent areas separated by the Apennines in Northern-Central Italy. Plant Pathol. J. 2011, 27, 354–359. [Google Scholar] [CrossRef]
- Prodi, A.; Tonti, S.; Nipoti, P.; Pancaldi, D.; Pisi, A. Identification of deoxynivalenol and nivalenol producing chemotypes of Fusarium graminearum isolates from durum wheat in a restricted area of northern Italy. J. Plant Pathol. 2009, 91, 727–731. [Google Scholar] [CrossRef]
- Somma, S.; Petruzzella, A.L.; Logrieco, A.F.; Meca, G.; Cacciola, O.S.; Moretti, A. Phylogenetic analyses of Fusarium graminearum strains from cereals in Italy, and characterisation of their molecular and chemical chemotypes. Crop Past. Sci. 2014, 65, 52–60. [Google Scholar] [CrossRef]
- Yli-Mattila, T.; Gagkaeva, T.; Ward, T.J.; Aoki, T.; Kistler, H.C.; O’Donnell, K. A novel Asian clade within the Fusarium graminearum species complex includes a newly discovered cereal head blight pathogen from the Russian Far East. Mycologia 2009, 101, 841–852. [Google Scholar] [CrossRef] [PubMed]
- Llorens, A.; Hinojo, M.J.; Mateo, R.; González-Jaén, M.T.; Valle-Algarra, F.M.; Logrieco, A.; Jiménez, M. Characterization of Fusarium spp. isolates by PCR-RFLP analysis of the intergenic spacer region of the rRNA gene (rDNA). Int. J. Food Microbiol. 2006, 106, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Isebaert, S.; De Sager, S.; Devreese, R.; Verhoeven, R.; Maene, P.; Heremans, B.; Haesaert, G. Mycotoxin-producing Fusarium species occurring in winter wheat in Belgium (Flanders) during 2002–2005. J. Phytopathology 2009, 157, 108–116. [Google Scholar] [CrossRef]
- Champeil, A.; Doré, T.; Fourbet, J.F. Fusarium head blight: Epidemiological origin of the effects of cultural practices on head blight attacks and the production of mycotoxins by Fusarium in wheat grains. Plant Sci. 2004, 166, 1389–1415. [Google Scholar] [CrossRef]
- Wang, Y.Z. Epidemiology and management of wheat scab in China. In Fusarium Head Scab: Global Status and Future Prospects; Dubin, H.J., Gilchrist, L., Reeves, J., McNab, A., Eds.; CIMMYT: EI Batan, Mexico, 1997; pp. 97–105. [Google Scholar]
- Manstretta, V.; Rossi, V. Effects of weather variables on ascospore discharge from Fusarium graminearum perithecia. PLoS ONE 2015, 10, e0138860. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Sun, H.Y.; Li, W.; Xia, Y.L.; Deng, Y.Y.; Zhang, A.X.; Chen, H.G. Fitness of three chemotypes of Fusarium graminearum species complex in major winter wheat-producing areas of China. PLoS ONE 2017, 12, e0174040. [Google Scholar] [CrossRef] [PubMed]
- Pancaldi, D.; Tonti, S.; Prodi, A.; Salomoni, D.; Dal Pra’, M.; Nipoti, P.; Alberti, I.; Pisi, A. Survey of the main causal agents of Fusarium head blight of durum wheat around Bologna, northern Italy. Phytopathol. Mediterr. 2010, 49, 258–266. [Google Scholar] [CrossRef]
- Parikka, P.; Hakala, K.; Tiilikkala, K. Expected shifts in Fusarium species’ composition on cereal grain in Northern Europe due to climatic change. Food Addit. Contam. Part A 2012, 29, 1543–1555. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.; Haber, S. Overview of some recent research developments in Fusarium head blight of wheat. Can. J. Plant Pathol. 2013, 35, 149–174. [Google Scholar] [CrossRef]
- Owczarek, M.; Filipiak, J. Contemporary changes of thermal conditions in Poland, 1951–2015. Bull. Geogr. Phys. Geogr. Ser. 2016, 10, 31–50. [Google Scholar] [CrossRef]
- World Meteorological Organization. WMO Statement on the state of the global climate in 2017; WMO-No. 1212; World Meteorological Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Kelly, A.C.; Ward, T.J. Population genomics of Fusarium graminearum reveals signatures of divergent evolution within a major cereal pathogen. PLoS ONE 2018, 13, e0194616. [Google Scholar] [CrossRef] [PubMed]
- Pasquali, M.; Beyer, M.; Logrieco, A.; Audenaert, K.; Balmas, V.; Basler, R.; Boutigny, A.-L.; Chrpová, J.; Czembor, E.; Gagkaeva, T.; et al. A European Database of Fusarium graminearum and F. culmorum Trichothecene Genotypes. Front. Microbiol. 2016, 7, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gale, L.R.; Harrison, S.A.; Ward, T.J.; O’Donnell, K.; Milus, E.A.; Gale, S.W.; Kistler, H.C. Nivalenol-type populations of Fusarium graminearum and F. asiaticum are prevalent on wheat in southern Louisiana. Phytopathology 2011, 101, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Gomes, L.B.; Ward, T.J.; Badiale-Furlong, E.; Del Ponte, E.M. Species composition, toxigenic potential and pathogenicity of Fusarium graminearum species complex isolates from southern Brazilian rice. Plant Pathol. 2015, 64, 980–987. [Google Scholar] [CrossRef]
- Dyer, R.B.; Plattner, R.D.; Kendra, D.F.; Brown, D.W. Fusarium graminearum TRI14 is required for high virulence and DON production on wheat but not for DON synthesis in vitro. J. Agric. Food Chem. 2005, 53, 9281–9287. [Google Scholar] [CrossRef] [PubMed]
- Chamary, J.; Hurst, L.D. Evidence for selection on synonymous mutations affecting stability of mRNA secondary structure in mammals. Genome Biol. 2005, 6, R75. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, J.B.; Kudla, G. Synonymous but not the same: The causes and consequences of codon bias. Nat. Rev. Genet. 2011, 12, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, J.M.; Domingo-Calap, P.; Sanjuán, R. The fitness effects of synonymous mutations in DNA and RNA viruses. Mol. Biol. Evol. 2012, 29, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Agashe, D.; Martinez-Gomez, N.C.; Drummond, D.A.; Marx, C.J. Good codons, bad transcript: Large reductions in gene expression and fitness arising from synonymous mutations in a key enzyme. Mol. Biol. Evol. 2013, 30, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.F.; Hinz, A.; Kassen, R. Adaptive synonymous mutations in an experimentally evolved Pseudomonas fluorescens population. Nat. Commun. 2014, 5, 4076. [Google Scholar] [CrossRef] [PubMed]
- Agashe, D.; Sane, M.; Phalnikar, K.; Diwan, G.D.; Habibullah, A.; Martinez-Gomez, N.C.; Sahasrabuddhe, V.; Polachek, W.; Wang, J.; Chubiz, L.M.; et al. Large effect beneficial synonymous mutations mediate rapid and parallel adaptation in a bacterium. Mol. Biol. Evol. 2016, 33, 1542–1553. [Google Scholar] [CrossRef] [PubMed]
- Bullerman, L.B. Mycotoxins Classifications. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Cambridge, UK, 2003; pp. 4080–4089. [Google Scholar]
- Smith, D.; Onions, A.H.S. The Preservation and Maintenance of Living Fungi; IMI Technical Handbook No.2; International Mycological Institute, CAB International: Oxford, UK, 1994. [Google Scholar]
- Bilska, K.; Kulik, T.; Ostrowska-Kołodziejczak, A.; Buśko, M.; Pasquali, M.; Beyer, M.; Baturo-Cieśniewska, A.; Juda, M.; Załuski, D.; Treder, K.; et al. Development of a highly sensitive FcMito qPCR assay for the quantification of the toxigenic fungal plant pathogen Fusarium culmorum. Toxins 2018, 10, 211. [Google Scholar] [CrossRef] [PubMed]
- Kulik, T.; Ostrowska, A.; Buśko, M.; Pasquali, M.; Beyer, M.; Stenglein, S.; Załuski, D.; Sawicki, J.; Treder, K.; Perkowski, J. Development of an FgMito assay: A highly sensitive mitochondrial based qPCR assay for quantification of Fusarium graminearum sensu stricto. Int. J. Food Microbiol. 2015, 210, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Waalwijk, C.; van der Heide, R.; de Vries, I.; van der Lee, T.; Schoen, C.; Corainville, G.C.; Häuser-Hahn, I.; Kastelein, P.; Köhl, J.; Lonnet, P.; et al. Quantitative detection of Fusarium species in wheat using TaqMan. Eur. J. Plant Pathol. 2004, 110, 481–494. [Google Scholar] [CrossRef]
- Kulik, T.; Buśko, M.; Bilska, K.; Ostrowska-Kołodziejczak, A.; van Diepeningen, A.D.; Perkowski, J.; Stenglein, S. Depicting the discrepancy between tri genotype and chemotype on the basis of strain CBS 139514 from a field population of F. graminearum sensu stricto from Argentina. Toxins 2016, 8, 330. [Google Scholar] [CrossRef] [PubMed]
- Kulik, T.; Abarenkov, K.; Buśko, M.; Bilska, K.; van Diepeningen, A.D.; Ostrowska-Kołodziejczak, A.; Krawczyk, K.; Brankovics, B.; Stenglein, S.; Sawicki, J.; Perkowski, J. ToxGen: An improved reference database for the identification of type B-trichothecene genotypes in Fusarium. PeerJ 2017, 5, e2992. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Year | Localization | Number of Isolates | F. graminearum s.s. | F. culmorum | F. avenaceum | F. poae | |||
|---|---|---|---|---|---|---|---|---|---|
| Total | 15ADON | 3ADON | Total | 3ADON | |||||
| Wheat kernels | |||||||||
| 2016 | Dobre Miasto | 22 | 21 (95.45%) | 15 (71.43%) | 6 (28.57%) | 0 | 0 | 1 (4.55%) | 0 |
| Łajsy | 85 | 85 (100%) | 75 (88.24% | 10 (11.76%) | 0 | 0 | 0 | 0 | |
| Tomaszkowo | 57 | 54 (94.74%) | 45 (83.33%) | 9 (16.67%) | 1 (1.75%) | 1 (100%) | 2 (3.51%) | 0 | |
| Tywęzy | 5 | 5 (100%) | 5 (100) | 0 | 0 | 0 | 0 | 0 | |
| Nidzica | 20 | 17 (85%) | 15 (88.24%) | 2 (11.76%) | 0 | 0 | 3 (15%) | 0 | |
| Kobierzyce | 7 | 6 (85.71%) | 6 (100%) | 0 | 0 | 0 | 1 (14.29) | 0 | |
| Kondratowice | 11 | 10 (90.91%) | 10 (100%) | 0 | 0 | 0 | 0 | 1 (9.09%) | |
| Modzurów | 59 | 44 (74.58%) | 37 (84.09%) | 7 (15.91%) | 8 (13.56%) | 8 (100%) | 7 (11.86%) | 0 | |
| Total in 2016 | 266 | 242 (90.98%) | 208 (85.95%) | 34 (14.05%) | 9 (3.38%) | 9 (100%) | 14 (5.26%) | 1 (0.38%) | |
| 2017 | Zduny | 11 | 9 (81.82%) | 8 (88.89%) | 1 (11.11%) | 0 | 0 | 2 (18.18%) | 0 |
| Tczew County | 26 | 26 (100%) | 25 (96.15%) | 1 (3.85%) | 0 | 0 | 0 | 0 | |
| Malbork | 6 | 6 (100%) | 6 (100%) | 0 | 0 | 0 | 0 | 0 | |
| Bałcyny | 8 | 5 (62.50%) | 5 (100%) | 0 | 0 | 0 | 3 (37.50%) | 0 | |
| Kętrzyn County | 11 | 6 (54.55%) | 5 (83.33%) | 1 (16.67%) | 0 | 0 | 5 (45.45%) | 0 | |
| Ostrołęka County | 76 | 35 (46.05%) | 31 (88.57%) | 4 (11.43%) | 7 (9.21%) | 7 (100%) | 34 (44.74%) | 0 | |
| Romaszówka | 15 | 9 (60.00%) | 7 (77.78%) | 2 (22.22%) | 2 (13.33%) | 2 (100%) | 4 (26.67%) | 0 | |
| Lipno County | 33 | 32 (96.97%) | 32 (100%) | 0 | 0 | 0 | 1 (3.03%) | 0 | |
| Warsaw West County | 11 | 6 (54.55%) | 6 (100%) | 0 | 0 | 0 | 5 (45.45%) | 0 | |
| Total in 2017 | 197 | 134 (68.02%) | 125 (93.28%) | 9 (6.72%) | 9 (4.75) | 9 (100%) | 54 (27.41%) | 0 | |
| Total wheat kernels | 463 | 376 (81.21%) | 333 (88.56%) | 43 (11.44%) | 18 (3.89%) | 18 (100%) | 68 (14.69%) | 1 (0.22%) | |
| Barley kernels from Modzurów | 56 | 9 (16.07%) | 9 (100%) | 0 | 0 | 0 | 45 (80.36%) | 2 (3.57%) | |
| Strain | Origin | Host | Year of Isolation | Culture Collection |
|---|---|---|---|---|
| 16-21-z | Poland, Dobre Miasto | winter wheat | 2016 | 1 |
| 16-92-z | Poland, Modzurów | winter wheat | 2016 | 1 |
| 16-43-tp | Poland, Tomaszkowo | winter wheat | 2016 | 1 |
| 16-462-z | Poland, Modzurów | winter wheat | 2016 | 1 |
| 03132 | Poland, Lewin Brzeski | winter wheat | 2003 | 1 |
| 04286 | Poland, Bałcyny | winter wheat | 2004 | 1 |
| 04501 | Poland, Martąg | winter wheat | 2004 | 1 |
| CBS 138561 | Poland | wheat | 2010 | 4 |
| 37 | Germany | unknown | 1994 | 2 |
| 74b | Germany | unknown | 2004 | 2 |
| N2-1 | Germany, Uelzen | winter wheat | 2017 | 1 |
| N4-1 | Germany, Uelzen | winter wheat | 2017 | 1 |
| N4-2 | Germany, Uelzen | winter wheat | 2017 | 1 |
| N5-1 | Germany, Uelzen | winter wheat | 2017 | 1 |
| N6-1 | Germany, Uelzen | winter wheat | 2017 | 1 |
| N6-2 | Germany, Uelzen | winter wheat | 2017 | 1 |
| N7-1 | Germany, Uelzen | winter wheat | 2017 | 1 |
| N8-2 | Germany, Uelzen | winter wheat | 2017 | 1 |
| N10-1 | Germany, Uelzen | winter wheat | 2017 | 1 |
| N10-2 | Germany, Uelzen | winter wheat | 2017 | 1 |
| 237 | Luxembourg | winter wheat | 2007 | 2 |
| 321 | Luxembourg | winter wheat | 2007 | 2 |
| 630 | Luxembourg | winter wheat | 2007 | 2 |
| 09-03a | Luxembourg | winter wheat | 2009 | 2 |
| 09-04a | Luxembourg | winter wheat | 2009 | 2 |
| 09-5a | Luxembourg | winter wheat | 2009 | 2 |
| 09-13a | Luxembourg | winter wheat | 2009 | 2 |
| 09-21a | Luxembourg | winter wheat | 2009 | 2 |
| 09-53b | Luxembourg | winter wheat | 2009 | 2 |
| St-6 | Russia, Stavropol region | winter wheat | 2015 | 3 |
| St-9 | Russia, Stavropol region | winter wheat | 2014 | 3 |
| Kr 275-1 | Russia, Krasnodar region | winter wheat | 2014 | 3 |
| 433-2 | Russia, Kabardino-Balkaria | winter wheat | 2014 | 3 |
| CBS 134070 | USA, Illinois, Urbana | Miscanthus giganteus | unknown | 4 |
| GZ3639, CBS 110266 | USA, Kansas | wheat | unknown | 4 |
| CBS 139513 | Argentina, Tandil | barley | 2011 | 4 |
| CBS 139514 | Argentina, Tapalqué | barley | 2010 | 4 |
| 114-2 | Argentina, Loberia | barley | 2012 | 1 |
| 23-4 | Argentina, Lauquen | barley | 2011 | 1 |
| Primer/Probe Sequence | Reaction Reagents | Reaction Condition | References | |
|---|---|---|---|---|
| Species | ||||
| F. culmorum | F: TCGTTGACGGTGAGGGTTGT R: GACTCGAACACGTCAACCAACTT Probe: FAM-CGGTTATTATTTCGAAAAGT- MGB | 2 μL gDNA, 14.3 μL H2O, 6.7 μM of each primer, 1.7 μM of probe, 3.6 μL TaqMan Fast Advanced Master Mix (Applied Biosystems, Foster City, CA, USA). | 95 °C for 20 s, (95 °C for 1 s, 60 °C for 30 s) × 40 | [79] |
| F. graminearum s.s. | F: TGGCCTGAATGAAGGATTTCTAG R: CATCGTTGTTAACTTATTGGAGATG Probe: FAM-TTAAACACTCAAACACTACA- MGB | [80] | ||
| F. avenaceum | F: CCATCGCCGTGGCTTTC R: CAAGCCCACAGACACGTTGT Probe: FAM-ACGCAATTGACTATTGC-MGB | 2 μL gDNA, 10.8 μL H2O, 6.7 μM of each primer, 1.7 μM of probe, 7.2 μL TaqMan Fast Advanced Master Mix (Applied Biosystems, Foster City, CA, USA). | 95 °C for 20 s, (95 °C for 1 s, 60 °C for 50 s) × 40 | [81] |
| F. poae | F: AAATCGGCGTATAGGGTTGAGATA R: GCTCACACAGAGTAACCGAAACCT Probe: FAM-CAAAATCACCCAACCGACCCTTTC-TAMRA | 50 °C for 2 min, 95 °C for 10 min, (95 °C for 15 s, 60 °C for 60 s) × 40 | ||
| Tri genotype | ||||
| 3ADON | F: CATGCGGGACTTTGATCGAT R: TTTGTCCGCTTTCTTTCTATCATAAA Probe: FAM-CTCACCGATCATGTTC-MGB | 2 μL gDNA, 10.8 μL H2O, 6.7 μM of each primer, 1.7 μM of probe, 7.2 μL TaqMan Fast Advanced Master Mix (Applied Biosystems, Foster City, CA, USA). | 95 °C for 20 s, (95 °C for 1 s, 60 °C for 50 s) × 40 | [43] |
| 15ADON | F: TCCAATCATTGCCAGCCTCTA R: TGATGCGGAACATGGTCTGT Probe: FAM-ATGAGGGACTTTGACCAAT-MGB | |||
| NIV | F: TCGCCAGTCTCTGCATGAAG R: CCTTATCCGCTTTCTTTCTATCATAAA Probe: FAM-CTGATCATGTCCCGCATC-MGB |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilska, K.; Jurczak, S.; Kulik, T.; Ropelewska, E.; Olszewski, J.; Żelechowski, M.; Zapotoczny, P. Species Composition and Trichothecene Genotype Profiling of Fusarium Field Isolates Recovered from Wheat in Poland. Toxins 2018, 10, 325. https://doi.org/10.3390/toxins10080325
Bilska K, Jurczak S, Kulik T, Ropelewska E, Olszewski J, Żelechowski M, Zapotoczny P. Species Composition and Trichothecene Genotype Profiling of Fusarium Field Isolates Recovered from Wheat in Poland. Toxins. 2018; 10(8):325. https://doi.org/10.3390/toxins10080325
Chicago/Turabian StyleBilska, Katarzyna, Sebastian Jurczak, Tomasz Kulik, Ewa Ropelewska, Jacek Olszewski, Maciej Żelechowski, and Piotr Zapotoczny. 2018. "Species Composition and Trichothecene Genotype Profiling of Fusarium Field Isolates Recovered from Wheat in Poland" Toxins 10, no. 8: 325. https://doi.org/10.3390/toxins10080325