Biodegradation of Ochratoxin A for Food and Feed Decontamination


Abstract
:
1. Introduction
1.1. Overview

 are 216, and 330 nm (ε 37,060 and 6050, respectively); and (b)
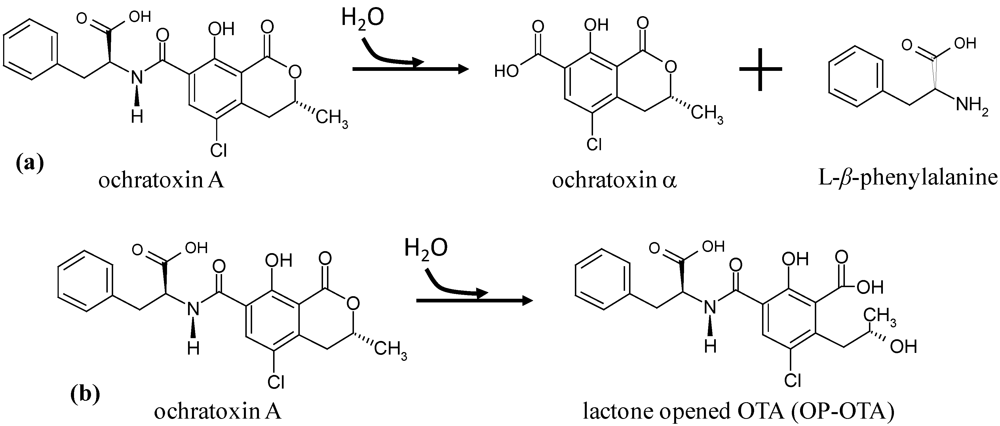
are 216, and 330 nm (ε 37,060 and 6050, respectively); and (b)  are 214 and 332 nm (ε 37,200 and 6330, respectively) [9]. The infrared spectrum in chloroform presents major peaks at 3380, 1723, 1678 and 1655 cm−1. OTA has weak acid properties with pKa1 4.2-4.4 and pKa2 7.0-7.3, from the carboxyl group of phenylalanine and from the phenolic hydroxyl group of the isocoumarin, respectively [7]. OTA produces a green-bluish fluorescence when excited by UV light (366 nm) using TLC, which changes to a dark blue fluorescence when exposed to ammonia vapors, aqueous NaHCO3 or NaOH [10]. The fluorescence properties are commonly used for detection and identification purposes in TLC and HPLC. LC-MS can be employed to detect OTA. The identity of OTA can be confirmed by converting it to the methyl ester [11], and/or to OTα using carboxypeptidase A. OTA is highly soluble in polar organic solvents, soluble in aqueous sodium hydrogen carbonate and slightly soluble in water.
are 214 and 332 nm (ε 37,200 and 6330, respectively) [9]. The infrared spectrum in chloroform presents major peaks at 3380, 1723, 1678 and 1655 cm−1. OTA has weak acid properties with pKa1 4.2-4.4 and pKa2 7.0-7.3, from the carboxyl group of phenylalanine and from the phenolic hydroxyl group of the isocoumarin, respectively [7]. OTA produces a green-bluish fluorescence when excited by UV light (366 nm) using TLC, which changes to a dark blue fluorescence when exposed to ammonia vapors, aqueous NaHCO3 or NaOH [10]. The fluorescence properties are commonly used for detection and identification purposes in TLC and HPLC. LC-MS can be employed to detect OTA. The identity of OTA can be confirmed by converting it to the methyl ester [11], and/or to OTα using carboxypeptidase A. OTA is highly soluble in polar organic solvents, soluble in aqueous sodium hydrogen carbonate and slightly soluble in water.1.2. Fungi

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi species | References |
|---|---|
| Aspergillus section Circumdati | |
| A. cretensis | [16] |
| A. flocculosus | [16] |
| A. melleus | [17] |
| A. ochraceus | [3,16] |
| A. ostianus | [17] |
| A. persii | [18] |
| A. petrakii | [17] |
| A. pseudoelegans | [16] |
| A. roseoglobulosus | [16] |
| A. sclerotiorum | [16,17,19] |
| A. steynii | [16] |
| A. sulphureus | [16,17,19] |
| A. westerdijkiae | [16] |
| Aspergillus section Flavi | |
| A. alliaceus (Petromycesalliaceus) | [19,20] |
| Petromyces albertensis | [15] |
| Aspergillus section Nigri | |
| A. carbonarius | [21,22,23] |
| A. lacticoffeatus | [23] |
| A. niger | [23,24] |
| A. sclerotioniger | [23] |
| Penicillium | |
| P. nordicum | [25] |
| P. verrucosum | [12,26,27] |
1.3. Biosynthetic pathway
1.4. Physiology of OTA production
1.5. Presence in commodities
| Food products | Contamination levels | References |
|---|---|---|
| Beans | 0.25-0.92 µg/Kg | [47] |
| Cocoa beans | 0.35-14.8 µg/Kg | [48] |
| Corn | 0.11-0.15 µg/Kg | [49] |
| Dried figs | <0.1-35.1 µg/Kg | [50] |
| Dried fruits | 0.1-30 µg/Kg | [51] |
| Grapes | 0.008-1.6 µg/Kg | [52] |
| Green coffee beans | 0-48 µg/Kg | [53] |
| Milk | 0.011-0.058 µg/L | [54] |
| Pork kidneys | 0-15 µg/Kg | [55] |
| Pork meat | 0-2.9 µg/Kg | [55] |
| Raisins | 0.2-53.6 μg/Kg | [56] |
| Rice | 1.0-27.3 µg/Kg | [57] |
| Spices | 4.2-103.2 µg/Kg | [58] |
| Wheat, Barley, oats | 0.1-17.8 µg/Kg | [59] |
| Wheat, oats and rye | 0.03-27 µg/Kg | [60] |
| Baby food | 0.06-2.4 µg/Kg | [61] |
| Beer | <0.01-0.135 µg/L | [62] |
| Breakfast cereals | 0.4-8.8 µg/Kg | [63] |
| Cocoa products | 0.22-0.77 µg/Kg | [64] |
| Grape juice | <0.003-0.311 µg/L | [65] |
| Pork products | <0.03-10.0 µg/Kg | [66] |
| Roasted coffee | 3.2-17.0 µg/Kg | [67] |
| Salami | <0.006-0.40 µg/Kg | [68] |
| Wine | <0.003-0.388 µg/L | [65] |
| Feed products | OTA levels average (µg/Kg) | References |
|---|---|---|
| Poultry feed | 0.5 | [71] |
| Bovine feed | 0.55 | |
| Corn grains | 3.95 | |
| Corn gluten | 1.95 | |
| Cotton seed | 6.19 | |
| Palm kernel | 3.19 | |
| Poultry feed | 27 | [72] |
| Pig feed | 34 | |
| Rabbit feed | 21.8 | |
| Groundnut cake | 50-100 | [73] |
| Millets | >100 | |
| Rice bran | 10-100 | |
| Sorghum | 10->100 | |
| Sunflower | 30-49 |
1.6. Toxicity
1.7. Elimination strategies
2. Biodegradation of Ochratoxin A

2.1. Microorganisms which degrade ochratoxin A
2.1.1. Protozoa
2.1.2. Bacteria
2.1.3. Yeasts
2.1.4. Filamentous fungi
2.1.5. Plant cell cultures
2.2. Enzymes which degrade ochratoxin A

| Commercial name | Origin | Main activity | Supplier | Reference |
|---|---|---|---|---|
| Carboxypeptidase A | Bovinus bovis | exopeptidase | Boehringer | [142] |
| Carboxypeptidase Y | Saccharomyces cerevisiae | exopeptidase | Sigma | not reported |
| Lipase | Aspergillus niger | Lipase | Amano Inc. | [138] |
| Enzyme preparations | - | Proteolysis | - | [143] |
| Protease A | Aspergillus niger | Acid protease | Amano Inc. | [139] |
| Prolyve PAC | Aspergillus niger | Acid protease | Lyven | [139] |
| Pancreatin 4XNF-P211P | Porcine pancreas | Amylase, lipase and protease | Biocatalysts | [139] |
| Crude extract | Aspergillus niger | OTA-hydrolase | - | [140] |
3. Conclusions
Acknowledgements
References
- Sargeant, K.; Sheridan, A.; Okelly, J. Toxicity associated with certain samples of groundnuts. Nature 1961, 192, 1096–1097. [Google Scholar]
- van der Merwe, K.J.; Steyn, P.S.; Fourie, L. Mycotoxins. Part II. The constitution of ochratoxins A, B, and C, metabolites of Aspergillus ochraceus Wilh. J. Chem. Soc. 1965, 7083–7088. [Google Scholar]
- van der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a toxic metabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar]
- Scott, D.B. Toxigenic fungi isolated from cereal and legume products. Mycopathol. Mycol. Appl. 1965, 25, 213. [Google Scholar]
- Hutchison, R.D.; Steyn, P.S.; Thompson, D.L. The isolation and structure of 4-hydroxyochratoxin A and 7-carboxy-3,4-dihydro-8-hydroxy-3-methylisocoumarin from Penicillium viridicatum. Tetrahedron Lett. 1971, 12, 4033–4036. [Google Scholar]
- Xiao, H.; Marquardt, R.R.; Abramson, D.; Frohlich, A.A. Metabolites of ochratoxins in rat urine and in a culture of Aspergillus ochraceus. Appl. Environ. Microbiol. 1996, 62, 648–655. [Google Scholar]
- Ringot, D.; Chango, A.; Schneider, Y.J.; Larondelle, Y. Toxicokinetics and toxicodynamics of ochratoxin A, an update. Chem. Biol. Interact. 2006, 159, 18–46. [Google Scholar]
- Cole, R.J.; Jarvis, B.B.; Schweikert, M.A. Ochratoxins and related metabolites. In Handbook of Secondary Fungal Metabolites; Academic: San Diego, CA, USA, 2003; Volume 3, pp. 615–624. [Google Scholar]
- Betina, V. Ochratoxins and related dihydroisocoumarins. In Mycotoxins: Chemical, Biological and Environmental Aspects; Elsevier: Amsterdam, The Netherland, 1989; Volume 9, pp. 151–173. [Google Scholar]
- Betina, V. Thin-layer chromatography of mycotoxins. In Chromatography of Mycotoxins: Techniques andApplications; Elsevier: Amsterdam, The Netherlands, 1993. [Google Scholar]
- Li, S.Z.; Marquardt, R.R.; Frohlich, A.A. Confirmation of ochratoxins in biological samples by conversion into methyl esters in acidified methanol. J. Agric. Food Chem. 1998, 46, 4307–4312. [Google Scholar]
- Pitt, J.I. Penicillium viridicatum, Penicillium verrucosum, and production of ochratoxin. Appl. Environ. Microbiol. 1987, 53, 266–269. [Google Scholar]
- Paterson, R.R.; Venâncio, A.; Lima, N. Solutions to Penicillium taxonomy crucial to mycotoxin research and health. Res. Microbiol. 2004, 155, 507–513. [Google Scholar]
- Vega, F.E.; Posada, F.; Peterson, S.W.; Gianfagna, T.J.; Chaves, F. Penicillium species endophytic in coffee plants and ochratoxin A production. Mycologia 2006, 98, 31–42. [Google Scholar]
- Frisvad, J.C.; Thrane, U.; Samson, R.A.; Pitt, J.I. Important mycotoxins and the fungi which produce them. Adv. Food Mycol. 2006, 571, 3–31. [Google Scholar]
- Frisvad, J.C.; Frank, J.M.; Houbraken, J.A.M.P.; Kuijpers, A.F.A.; Samson, R.A. New ochratoxin A producing species of Aspergillus section Circumdati. Stud. Mycol. 2004, 50, 23–43. [Google Scholar]
- Hesseltine, C.W.; Vandegraft, E.E.; Fennell, D.I.; Smith, M.L.; Shotwell, O.L. Aspergilli as ochratoxin producers. Mycologia 1972, 64, 539–550. [Google Scholar]
- Ciegler, A. Bioproduction of ochratoxin A and penicillic acid by members of the Aspergillus ochraceus group. Can. J. Microbiol. 1972, 18, 631–636. [Google Scholar]
- Varga, J.; Kevei, E.; Rinyu, E.; Téren, J.; Kozakiewicz, Z. Ochratoxin production by Aspergillus species. Appl. Environ. Microbiol. 1996, 62, 4461–4464. [Google Scholar]
- Bayman, P.; Baker, J.L.; Doster, M.A.; Michailides, T.J.; Mahoney, N.E. Ochratoxin production by the Aspergillus ochraceus group and Aspergillus alliaceus. Appl. Environ. Microbiol. 2002, 68, 2326–2329. [Google Scholar]
- Horie, Y. Productivity of ochratoxin A of Aspergillus carbonarius in Aspergillus section Nigri. Nippon Kingakkai Kaiho 1995, 36, 73–76. [Google Scholar]
- Téren, J.; Varga, J.; Hamari, Z.; Rinyu, E.; Kevei, F. Immunochemical detection of ochratoxin A in black Aspergillus strains. Mycopathologia 1996, 134, 171–176. [Google Scholar]
- Samson, R.A.; Houbraken, J.A.M.P.; Kuijpers, A.F.A.; Frank, J.M.; Frisvad, J.C. New ochratoxin A or sclerotium producing species in Aspergillus section Nigri. Stud. Mycol. 2004, 50, 45–61. [Google Scholar]
- Abarca, M.L.; Bragulat, M.R.; Castellá, G.; Cabañes, F.J. Ochratoxin A production by strains of Aspergillus niger var. niger. Appl. Environ. Microbiol. 1994, 60, 2650–2652. [Google Scholar]
- Larsen, T.O.; Svendsen, A.; Smedsgaard, J. Biochemical characterization of ochratoxin A-producing strains of the genus Penicillium. Appl. Environ. Microbiol. 2001, 67, 3630–3635. [Google Scholar]
- van Walbeek, W.; Scott, P.M.; Harwig, J.; Lawrence, J.W. Penicillium viridicatum Westling: A new source of ochratoxin A. Can. J. Microbiol. 1969, 15, 1281–1285. [Google Scholar]
- Ciegler, A.; Fennell, D.I.; Sansing, G.A.; Detroy, R.W.; Bennett, G.A. Mycotoxin-producing strains of Penicillium viridicatum: Classification into subgroups. Appl. Microbiol. 1973, 26, 271–278. [Google Scholar]
- Serra, R.; Mendonca, C.; Venâncio, A. Fungi and ochratoxin A detected in healthy grapes for wine production. Lett. Appl. Microbiol. 2006, 42, 42–47. [Google Scholar] [Green Version]
- Pitt, J.I.; Taniwaki, M.H.; Teixeira, A.A.; Iamanaka, B.T. Distribution of Aspergillus ochraceus, A. niger and A. carbonarius in coffee in four regions of Brazil. In Proceedings of the 19th Colloquium: Moisture Management for Mould Prevention in Coffee, Trieste, Italy; 2001. [Google Scholar]
- Esteban, A.; Abarca, M.L.; Bragulat, M.R.; Cabañes, F.J. Effects of temperature and incubation time on production of ochratoxin A by black aspergilli. Res. Microbiol. 2004, 155, 861–866. [Google Scholar]
- Ferreira, N.P.; Pitout, M.J. Biogenesis of Ochratoxin. J. S. Afr. Chem. I 1969, 22, S1. [Google Scholar]
- Searcy, J.W.; Davis, N.D.; Diener, U.L. Biosynthesis of Ochratoxin A. Appl. Microbiol. 1969, 18, 622–627. [Google Scholar]
- Steyn, P.S.; Holzapfel, C.W.; Ferreira, N.P. The biosynthesis of the ochratoxins, metabolites of Aspergillus ochraceus. Phytochemistry 1970, 9, 1977–1983. [Google Scholar]
- Wei, R.D.; Strong, F.M.; Smalley, E.B. Incorporation of chlorine-36 into ochratoxin A. Appl. Microbiol. 1971, 22, 276–277. [Google Scholar]
- Karolewiez, A.; Geisen, R. Cloning a part of the ochratoxin A biosynthetic gene cluster of Penicillium nordicum and characterization of the ochratoxin polyketide synthase gene. Syst. Appl. Microbiol. 2005, 28, 588–595. [Google Scholar]
- O'Callaghan, J.; Caddick, M.X.; Dobson, A.D.W. A polyketide synthase gene required for ochratoxin A biosynthesis in Aspergillus ochraceus. Microbiology 2003, 149, 3485–3491. [Google Scholar]
- Atoui, A.; Dao, H.P.; Mathieu, F.; Lebrihi, A. Amplification and diversity analysis of ketosynthase domains of putative polyketide synthase genes in Aspergillus ochraceus and Aspergillus carbonarius producers of ochratoxin. Mol. Nutr. Food Res. 2006, 50, 488–493. [Google Scholar]
- Mitchell, D.; Parra, R.; Aldred, D.; Magan, N. Water and temperature relations of growth and ochratoxin A production by Aspergillus carbonarius strains from grapes in Europe and Isra. J. Appl. Microbiol. 2004, 97, 439–445. [Google Scholar]
- Ramos, A.J.; Labernia, N.; Marin, S.; Sanchís, V.; Magan, N. Effect of water activity and temperature on growth and ochratoxin production by three strains of Aspergillus ochraceus on a barley extract medium and on barley grains. Int. J. Food Microbiol. 1998, 44, 133–140. [Google Scholar]
- Northolt, M.D.; Vanegmond, H.P.; Paulsch, W.E. Ochratoxin A production by some fungal species in relation to water activity and temperature. J. Food Prot. 1979, 42, 485–490. [Google Scholar]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage; Blackie Academic & Professional: London, UK, 1997. [Google Scholar]
- Baxter, E.D.; Slaiding, I.R.; Kelly, B. Behavior of ochratoxin A in brewing. J. Am. Soc. Brew. Chem. 2001, 59, 98–100. [Google Scholar]
- Scott, P.M.; Kanhere, S.R.; Lawrence, G.A.; Daley, E.F.; Farber, J.M. Fermentation of wort containing added ochratoxin A and fumonisins B1 and B2. Food Addit. Contam. 1995, 12, 31–40. [Google Scholar]
- Scudamore, K.A.; Banks, J.; MacDonald, S.J. Fate of ochratoxin A in the processing of whole wheat grains during milling and bread production. Food Addit. Contam. 2003, 20, 1153–1163. [Google Scholar]
- Suarez-Quiroz, M.; De Louise, B.; Gonzalez-Rios, O.; Barel, M.; Guyot, B.; Schorr-Galindo, S.; Guiraud, J.P. The impact of roasting on the ochratoxin A content of coffee. Int. J. Food Sci. Tech. 2005, 40, 605–611. [Google Scholar]
- Fernandes, A.; Ratola, N.; Cerdeira, A.; Alves, A.; Venancio, A. Changes in ochratoxin A concentration during winemaking. Am. J. Enol. Vitic. 2007, 58, 92–96. [Google Scholar]
- Domijan, A.-M.; Peraica, M.; Zlender, V.; Cvjetkovic, B.; Jurjevic, Z.; Topolovec-Pintaric, S.; Ivic, D. Seed-borne fungi and ochratoxin A contamination of dry beans (Phaseolus vulgaris L.) in the Republic of Croatia. Food Chem. Toxicol. 2005, 43, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Amezqueta, S.; González-Peñas, E.; Murillo, M.; De Cerain, A.L. Validation of a high-performance liquid chromatography analytical method for ochratoxin A quantification in cocoa beans. Food Addit. Contam. 2004, 21, 1096–1106. [Google Scholar]
- Shotwell, O.L.; Hesseltine, C.W.; Goulden, M.L. Ochratoxin A: Occurrence as natural contaminant of a corn sample. Appl. Microbiol. 1969, 17, 765–766. [Google Scholar]
- Senyuva, H.Z.; Gilbert, J.; Ozcan, S.; Ulken, U. Survey for co-occurrence of ochratoxin A and aflatoxin B1, in dried figs in Turkey by using a single laboratory-validated alkaline extraction method for ochratoxin. J. Food Prot. 2005, 68, 1512–1515. [Google Scholar]
- Iamanaka, B.T.; Taniwaki, M.H.; Menezes, H.C.; Vicente, E.; Fungaro, M.H.P. Incidence of toxigenic fungi and ochratoxin A in dried fruits sold in Brazil. Food Addit. Contam. 2005, 22, 1258–1263. [Google Scholar]
- Serra, R.; Mendonca, C.; Venancio, A. Ochratoxin A occurrence and formation in Portuguese wine grapes at various stages of maturation. Int. J. Food Microbiol. 2006, 111, S35–S39. [Google Scholar]
- Romani, S.; Sacchetti, G.; Chaves Lopez, C.; Pinnavaia, G.G.; Dalla Rosa, M. Screening on the occurrence of ochratoxin A in green coffee beans of different origins and types. J. Agric. Food Chem. 2000, 48, 3616–3619. [Google Scholar]
- Skaug, M.A. Analysis of Norwegian milk and infant formulas for ochratoxin A. Food Addit. Contam. 1999, 16, 75–78. [Google Scholar] [Green Version]
- Jørgensen, K.; Petersen, A. Content of ochratoxin A in paired kidney and meat samples from healthy Danish slaughter pigs. Food Addit. Contam. 2002, 19, 562–567. [Google Scholar]
- MacDonald, S.; Wilson, P.; Barnes, K.; Damant, A.; Massey, R.; Mortby, E.; Shepherd, M.J. Ochratoxin A in dried vine fruit: Method development and survey. Food Addit. Contam. 1999, 16, 253–260. [Google Scholar]
- Gonzalez, L.; Juan, C.; Soriano, J.M.; Molto, J.C.; Manes, J. Occurrence and daily intake of ochratoxin A of organic and non-organic rice and rice products. Int. J. Food Microbiol. 2006, 107, 223–227. [Google Scholar]
- Thirumala-Devi, K.; Mayo, M.A.; Reddy, G.; Emmanuel, K.E.; Larondelle, Y.; Reddy, D.V. Occurrence of ochratoxin A in black pepper, coriander, ginger and turmeric in India. Food Addit. Contam. 2001, 18, 830–835. [Google Scholar]
- Scudamore, K.A.; Patel, S.; Breeze, V. Surveillance of stored grain from the 1997 harvest in the United Kingdom for ochratoxin A. Food Addit. Contam. 1999, 16, 281–290. [Google Scholar]
- Scott, P.M.; van Walbeek, W.; Kennedy, B.; Anyeti, D. Mycotoxins (ochratoxin A, citrinin, and sterigmatocystin) and toxigenic fungi in grains and other agricultural products. J. Agric. Food Chem. 1972, 20, 1103–1109. [Google Scholar]
- Lombaert, G.A.; Pellaers, P.; Roscoe, V.; Mankotia, M.; Neil, R.; Scott, P.M. Mycotoxins in infant cereal foods from the Canadian retail market. Food Addit. Contam. 2003, 20, 494–504. [Google Scholar]
- Visconti, A.; Pascale, M.; Centonze, G. Determination of ochratoxin A in domestic and imported beers in Italy by immunoaffinity clean-up and liquid chromatograph. J. Chromatogr. A 2000, 888, 321–326. [Google Scholar]
- Molinié, A.; Faucet, V.; Castegnaro, M.; Pfohl-Leszkowicz, A. Analysis of some breakfast cereals on the French market for their contents of ochratoxin A, citrinin and fumonisin B1: development of a method for simultaneous extraction of ochratoxin A and citrinin. Food Chem. 2005, 92, 391–400. [Google Scholar]
- Tafuri, A.; Ferracane, R.; Ritieni, A. Ochratoxin A in Italian marketed cocoa products. Food Chem. 2004, 88, 487–494. [Google Scholar]
- Zimmerli, B.; Dick, R. Ochratoxin A in table wine and grape-juice: Occurrence and risk assessment. Food Addit. Contam. 1996, 13, 655–668. [Google Scholar]
- Pietri, A.; Bertuzzi, T.; Gualla, A.; Piva, G. Occurrence of ochratoxin A in raw ham muscles and in pork products from northern Italy. Ital. J. Food Sci. 2006, 18, 99–106. [Google Scholar]
- Tsubouchi, H.; Terada, H.; Yamamoto, K.; Hisada, K.; Sakabe, Y. Ochratoxin A found in commercial roast coffee. J. Agric. Food Chem. 1988, 36, 540–542. [Google Scholar]
- Monaci, L.; Palmisano, F.; Matrella, R.; Tantillo, G. Determination of ochratoxin A at part-per-trillion level in Italian salami by immunoaffinity clean-up and high-performance liquid chromatography with fluorescence detection. J. Chromatogr. A 1090, 184–187. [Google Scholar]
- Scudamore, K.A.; Banks, J.N.; Guy, R.C.E. Fate of ochratoxin A in the processing of whole wheat grain during extrusion. Food Addit. Contam. 2004, 21, 488–497. [Google Scholar]
- Scudamore, K.A. Prevention of ochratoxin A in commodities and likely effects of processing fractionation and animal feed. Food Addit. Contam. 2005, 22, 17–25. [Google Scholar]
- Jaimez, J.; Fente, C.A.; Franco, C.M.; Cepeda, A.; Vazquez, B.I. A survey of the fungal contamination and presence of ochratoxin A and zearalenone on Spanish feed and raw material. J. Sci. Food Agric. 2004, 84, 832–840. [Google Scholar]
- Dalcero, A.; Magnoli, C.; Hallak, C.; Chiacchiera, S.M.; Palacio, G.; Rosa, C.A.d.R. Detection of ochratoxin A in animal feeds and capacity to produce this mycotoxin by Aspergillus section Nigri in Argentina. Food Addit. Contam. 2002, 19, 1065–1072. [Google Scholar]
- Thirumala-Devi, K.; Mayo, M.A.; Reddy, G.; Reddy, D.V.R. Occurrence of aflatoxins and ochratoxin A in Indian poultry feeds. J. Food Prot. 2002, 65, 1338–1340. [Google Scholar]
- Xiao, H.; Madhyastha, S.; Marquardt, R.R.; Li, S.; Vodela, J.K.; Frohlich, A.A.; Kemppainen, B.W. Toxicity of ochratoxin A, Its opened lactone form and several of its analogs: Structure-activity relationship. Toxicol. Appl. Pharmacol. 1996, 137, 182–192. [Google Scholar]
- Muller, G.; Rosner, H.; Rohrmann, B.; Erler, W.; Geschwend, G.; Grafe, U.; Burkert, B.; Moller, U.; Diller, R.; Sachse, K.; Kohler, H. Effects of the mycotoxin ochratoxin A and some of its metabolites on the human cell line THP-1. Toxicology 2003, 184, 69–82. [Google Scholar]
- Fuchs, R.; Hult, K.; Peraica, M.; Radic, B.; Plestina, R. Conversion of ochratoxin C into ochratoxin A in vivo. Appl. Environ. Microbiol. 1984, 48, 41–42. [Google Scholar]
- Li, S.; Marquardt, R.R.; Frohlich, A.A.; Vitti, T.G.; Crow, G. Pharmacokinetics of ochratoxin A and its metabolites in rats. Toxicol. Appl. Pharmacol. 1997, 145, 82–90. [Google Scholar]
- Krogh, P. Role of ochratoxin in disease causation. Food Chem. Toxicol. 1992, 30, 213–224. [Google Scholar]
- Pfohl-Leszkowicz, A.; Petkova-Bocharova, T.; Chernozemsky, I.N.; Castegnaro, M. Balkan endemic nephropathy and associated urinary tract tumours: a review on aetiological causes and the potential role of mycotoxin. Food Addit. Contam. 2002, 19, 282–302. [Google Scholar]
- Pfohl-Leszkowicz, A.; Manderville, R.A. Ochratoxin A: An overview on toxicity and carcinogenicity in animals and human. Mol. Nutr. Food Res. 2007, 51, 61–99. [Google Scholar]
- Pfohl-Leszkowicz, A. Ochratoxin A and Aristolochic Acid Involvement in Nephropathies and Associated Urothelial Tract Tumour. Arhiv za Higijenu Rada i Toksikologiju 2009, 60, 465–483. [Google Scholar]
- IARC, Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; World Health Organization: Lyon, France, 1993; Volume 56, pp. 489–521.
- O'Brien, E.; Dietrich, D.R. Ochratoxin A: The continuing enigma. Crit. Rev. Microbiol. 2005, 35, 33–60. [Google Scholar]
- Chu, F.S. A comparative study of the interaction of ochratoxins with bovine serum albumin. Biochem. Pharmacol. 1974, 23, 1105–1113. [Google Scholar]
- Perry, J.L.; Christensen, T.; Goldsmith, M.R.; Toone, E.J.; Beratan, D.N.; Simon, J.D. Binding of ochratoxin A to human serum albumin stabilized by a protein-ligand ion pair. J. Phys. Chem. B 2003, 107, 7884–7888. [Google Scholar]
- Kumagai, S. Ochratoxin A: Plasma concentration and excretion into bile and urine in albumin-deficient rats. Food Chem. Toxicol. 1985, 23, 941–943. [Google Scholar]
- Creppy, E.E.; Stormer, F.C.; Roschenthaler, R.; Dirheimer, G. Effects of two metabolites of ochratoxin A, (4R)-4-hydroxyochratoxin A and ochratoxin α, on immune response in mice. Infect. Immun. 1983, 39, 1015–1018. [Google Scholar]
- Bruinink, A.; Rasonyi, T.; Sidler, C. Differences in neurotoxic effects of ochratoxin A, ochracin and ochratoxin-α in vitro. Nat. Toxins 1998, 6, 173–177. [Google Scholar]
- Zimmerli, B.; Dick, R. Determination of ochratoxin A at the ppt level in human blood, serum, milk and some foodstuffs by high-performance liquid chromatography with enhanced fluorescence detection and immunoaffinity column cleanup: methodology and Swiss data. J. Chromatogr. B 1995, 666, 85–99. [Google Scholar]
- Ueno, Y.; Maki, S.; Lin, J.; Furuya, M.; Sugiura, Y.; Kawamura, O. A 4-year study of plasma ochratoxin A in a selected population in Tokyo by immunoassay and immunoaffinity column-linked HPLC. Food Chem. Toxicol. 1998, 36, 445–449. [Google Scholar]
- Sangare-Tigori, B.; Moukha, S.; Kouadio, J.H.; Dano-Djedje, S.; Betbeder, A.-M.; Achour, A.; Creppy, E.E. Ochratoxin A in human blood in Abidjan, Cote d'Ivoire. Toxicon 2006, 47, 894–900. [Google Scholar]
- Skaug, M.A.; Helland, I.; Solvoll, K.; Saugstad, O.D. Presence of ochratoxin A in human milk in relation to dietary intake. Food Addit. Contam. 2001, 18, 321–327. [Google Scholar] [Green Version]
- Turconi, G.; Guarcello, M.; Livieri, C.; Comizzoli, S.; Maccarini, L.; Castellazzi, A.; Pietri, A.; Piva, G.; Roggi, C. Evaluation of xenobiotics in human milk and ingestion by the newborn: An epidemiological survey in Lombardy (Northern Italy). Eur. J. Nutr. 2004, 43, 191–197. [Google Scholar]
- Micco, C.; Miraglia, M.; Brera, C.; Corneli, S.; Ambruzzi, A. Evaluation of ochratoxin A level in human milk in Italy. Food Addit. Contam. 1995, 12, 351–354. [Google Scholar]
- Amezqueta, S.; Gonzalez-Penas, E.; Murillo-Arbizu, M.; De Cerain, A.L. Ochratoxin A decontamination: A review. Food Control 2009, 20, 326–333. [Google Scholar]
- Riley, R.T.; Norred, W.P. Mycotoxin prevention and decontamination-A case study on maize. Food Nutr. Agric. 1999, 23, 25–32. [Google Scholar]
- Karlovsky, P. Biological detoxification of fungal toxins and its use in plant breeding, feed and food production. Nat. Toxins 1999, 7, 1–23. [Google Scholar]
- Shimizu, S.; Kataoka, M.; Honda, K.; Sakamoto, K. Lactone-ring-cleaving enzymes of microorganisms: their diversity and applications. J. Biotechnol. 2001, 92, 187–194. [Google Scholar]
- Hult, K.; Teiling, A.; Gatenbeck, S. Degradation of ochratoxin A by a ruminant. Appl. Environ. Microbiol. 1976, 32, 443–444. [Google Scholar]
- Kiessling, K.H.; Pettersson, H.; Sandholm, K.; Olsen, M. Metabolism of aflatoxin, ochratoxin, zearalenone, and three trichothecenes by intact rumen fluid, rumen protozoa, and rumen bacteria. Appl. Environ. Microbiol. 1984, 47, 1070–1073. [Google Scholar]
- zpinar, H.; Bilal, T.; Abas, I.; Kutay, C. Degradation of ochratoxin A in rumen fluid in vitro. Med. Biol. 2002, 9, 66–69. [Google Scholar]
- Hohler, D.; Sudekum, K.H.; Wolffram, S.; Frohlich, A.A.; Marquardt, R.R. Metabolism and excretion of ochratoxin A fed to sheep. J. Anim. Sci. 1999, 77, 1217–1223. [Google Scholar]
- Wegst, W.; Lingens, F. Bacterial degradation of ochratoxin A. FEMS Microbiol. Lett. 1983, 17, 341–344. [Google Scholar]
- Hwang, C.A.; Draughon, F.A. Degradation of ochratoxin A by Acinetobacter calcoaceticus. J. Food Prot. 1994, 57, 410–414. [Google Scholar]
- Petchkongkaew, A.; Taillandier, P.; Gasaluck, P.; Lebrihi, A. Isolation of Bacillus spp. from Thai fermented soybean (Thua-nao): Screening for aflatoxin B1 and ochratoxin A detoxification. J. Appl. Microbiol. 2008, 104, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Skrinjar, M.; Rasic, J.L.; Stojicic, V. Lowering of ochratoxin A level in milk by yoghurt bacteria and bifidobacteria. Folia Microbiol. (Praha) 1996, 41, 26–28. [Google Scholar] [CrossRef] [PubMed]
- Böhm, J.; Grajewski, J.; Asperger, H.; Rabus, B.; Razzazi, E. Study on biodegradation of some trichothecenes (NIV DON, DAS, T-2) and ochratoxin A by use of probiotic microorganisms. Mycot. Res. 2000, 16, 70–74. [Google Scholar]
- Piotrowska, M.; Zakowska, Z. The biodegradation of ochratoxin A in food products by lactic acid bacteria and baker's yeast. In Progress in Biotechnology (Food Biotechnology); Bielecki, S., Tramper, J., Polak, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; Volume 17, pp. 307–310. [Google Scholar]
- Turbic, A.; Ahokas, J.T.; Haskard, C.A. Selective in vitro binding of dietary mutagens, individually or in combination, by lactic acid bacteria. Food Addit. Contam. 2002, 19, 144–152. [Google Scholar]
- Piotrowska, M.; Zakowska, Z. The elimination of ochratoxin A by lactic acid bacteria strains. Pol. J. Microbiol. 2005, 54, 279–286. [Google Scholar]
- Del Prete, V.; Rodriguez, H.; Carrascosa, A.V.; Rivas, B.D.L.; Garcia-Moruno, E.; Munoz, R. In vitro removal of ochratoxin A by wine lactic acid bacteria. J. Food Prot. 2007, 70, 2155–2160. [Google Scholar]
- Fuchs, S.; Sontag, G.; Stidl, R.; Ehrlich, V.; Kundi, M.; Knasmuller, S. Detoxification of patulin and ochratoxin A, two abundant mycotoxins, by lactic acid bacter. Food Chem. Toxicol. 2008, 46, 1398–1407. [Google Scholar]
- Shetty, P.H.; Jespersen, L. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci. Tech. 2006, 17, 48–55. [Google Scholar]
- Bejaoui, H.; Mathieu, F.; Lebrihi, A.; Taillandier, P. Ochratoxin A removal in synthetic and natural grape juices by selected oenological Saccharomyces strains. J. Appl. Microbiol. 2004, 97, 1038–1044. [Google Scholar]
- Bizaj, E.; Mavri, J.; Cus, F.; Raspor, A. Removal of ochratoxin A in Saccharomyces cerevisiae liquid cultures. S. Afr. J. Enol. Vitic. 2009, 30, 151–155. [Google Scholar]
- Cecchini, F.; Morassut, M.; Garcia Moruno, E.; Di Stefano, R. Influence of yeast strain on ochratoxin A content during fermentation of white and red must. Food Microbiol. 2006, 23, 411–417. [Google Scholar]
- Moruno, E.G.; Sanlorenzo, C.; Boccaccino, B.; Di Stefano, R. Treatment with yeast to reduce the concentration of ochratoxin A in red wine. Am. J. Enol. Vitic. 2005, 56, 73–76. [Google Scholar]
- Ringot, D.; Lerzy, B.; Bonhoure, J.P.; Auclair, E.; Oriol, E.; Larondelle, Y. Effect of temperature on in vitro ochratoxin A biosorption onto yeast cell wall derivatives. Process Biochem. 2005, 40, 3008–3016. [Google Scholar]
- Yiannikouris, A.; andre, G.; Poughon, L.; Francois, J.; Dussap, C.G.; Jeminet, G.; Bertin, G.; Jouany, J.P. Chemical and conformational study of the interactions involved in mycotoxin complexation with β-D-glucans. Biomacromolecules 2006, 7, 1147–1155. [Google Scholar]
- Raju, M.V.L.N.; Devegowda, G. Esterified-glucomannan in broiler chicken diets-contaminated with aflatoxin, ochratoxin and T-2 toxin: Evaluation of its binding ability (in vitro) and efficacy as immunomodulator. Asian Australas. J. Anim. Sci. 2002, 15, 1051–1056. [Google Scholar]
- Oguz, H.; Parlat, S.S. Effects of dietary mannanoligosaccharide on performance of Japanese quail affected by aflatoxicosis. S. Afr. J. Anim. Sci. 2004, 34, 144–148. [Google Scholar]
- Schatzmayr, G.; Heidler, D.; Fuchs, E.; Mohnl, M.; Täubel, M.; Loibner, A.P.; Braun, R.; Binder, E.M. Investigation of different yeast strains for the detoxification of ochratoxin A. Mycot. Res. 2003, 19, 124–128. [Google Scholar]
- Molnar, O.; Schatzmayr, G.; Fuchs, E.; Prillinger, H. Trichosporon mycotoxinivorans sp nov., a new yeast species useful in biological detoxification of various mycotoxins. Syst. Appl. Microbiol. 2004, 27, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Politis, I.; Fegeros, K.; Nitsch, S.; Schatzmayr, G.; Kantas, D. Use of Trichosporon mycotoxinivorans to suppress the effects of ochratoxicosis on the immune system of broiler chicks. Br. Poult. Sci. 2005, 46, 58–65. [Google Scholar]
- Hofstetter, U.; Schatzmayr, D.; Schatzmayr, G.; Binder, E.M. Successful detoxification of ochratoxin A in weaning piglets. J. Anim. Sci. 2006, 84, 308. [Google Scholar]
- Hickey, P.W.; Sutton, D.A.; Fothergill, A.W.; Rinaldi, M.G.; Wickes, B.L.; Schmidt, H.J.; Walsh, T.J. Trichosporon mycotoxinivorans: A novel respiratory pathogen in patients with cystic fibrosis. J. Clin. Microbiol. 2009, 47, 3091–3097. [Google Scholar]
- Peteri, Z.; Teren, J.; Vagvolgyi, C.; Varga, J. Ochratoxin degradation and adsorption caused by astaxanthin-producing yeasts. Food Microbiol. 2007, 24, 205–210. [Google Scholar]
- de Felice, D.V.; Solfrizzo, M.; De Curtis, F.; Lima, G.; Visconti, A.; Castoria, R. Strains of Aureobasidium pullulans can lower ochratoxin A contamination in wine grapes. Phytopathology 2008, 98, 1261–1270. [Google Scholar]
- Hawkes, M.; Rennie, R.; Sand, C.; Vaudry, W. Aureobasidium pullulans infection: Fungemia in an infant and a review of human cases. Diagn. Microbiol. Infect. Dis. 2005, 51, 209–213. [Google Scholar]
- Varga, J.; Rigó, K.; Téren, J. Degradation of ochratoxin A by Aspergillus species. Int. J. Food Microbiol. 2000, 59, 1–7. [Google Scholar]
- Abrunhosa, L.; Serra, R.; Venâncio, A. Biodegradation of ochratoxin A by fungi isolated from grapes. J. Agric. Food Chem. 2002, 50, 7493–7496. [Google Scholar]
- Varga, J.; Peteri, Z.; Tabori, K.; Teren, J.; Vagvolgyi, C. Degradation of ochratoxin A and other mycotoxins by Rhizopus isolates. Int. J. Food Microbiol. 2005, 99, 321–328. [Google Scholar]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Biodegradation of ochratoxin A by Aspergillus section Nigri species isolated from French grapes: A potential means of ochratoxin A decontamination in grape juices and musts. FEMS Microbiol. Lett. 2006, 255, 203–208. [Google Scholar]
- Valero, A.; Sanchis, V.; Ramos, A.J.; Marin, S. Brief in vitro study on Botrytis cinerea and Aspergillus carbonarius regarding growth and ochratoxin. Lett. Appl. Microbiol. 2008, 47, 327–332. [Google Scholar]
- Engelhardt, G. Degradation of ochratoxin A and B by the white rot fungus Pleurotus ostreatus. Mycot. Res. 2002, 18, 37–43. [Google Scholar]
- Ruhland, M.; Engelhardt, G.; Wallnofer, P.R. Transformation of the mycotoxin ochratoxin A in plants. 2. Time course and rates of degradation and metabolite production in cell-suspension cultures of different crop plants. Mycopathologia 1996, 134, 97–102. [Google Scholar] [CrossRef] [PubMed]
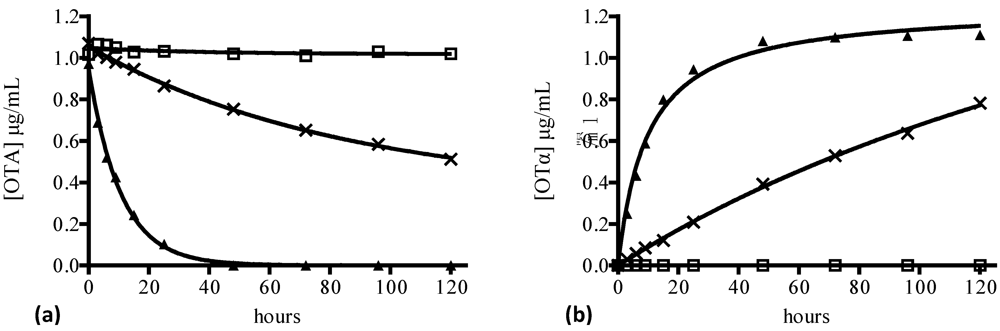
- Pitout, M.J. The hydrolysis of ochratoxin A by some proteolytic enzymes. Biochem. Pharmacol. 1969, 18, 485–491. [Google Scholar]
- Stander, M.A.; Bornscheuer, U.T.; Henke, E.; Steyn, P.S. Screening of commercial hydrolases for the degradation of ochratoxin A. J. Agric. Food Chem. 2000, 48, 5736–5739. [Google Scholar]
- Abrunhosa, L.; Santos, L.; Venâncio, A. Degradation of ochratoxin A by proteases and by a crude enzyme of Aspergillus niger. Food Biotechnol. 2006, 20, 231–242. [Google Scholar]
- Abrunhosa, L.; Venâncio, A. Isolation and purification of an enzyme hydrolyzing ochratoxin A from Aspergillus niger. Biotechnol. Lett. 2007, 29, 1909–1914. [Google Scholar]
- Abrunhosa, L.; Venâncio, A. 2008. Unpublished work.
- Stander, M.A.; Steyn, P.S.; van der Westhuizen, F.H.; Payne, B.E. A kinetic study into the hydrolysis of the ochratoxins and analogues by carboxypeptidase A. Chem. Res. Toxicol. 2001, 14, 302–304. [Google Scholar]
- Schatzmayr, G.; Binder, J.; Thimm, N.; Heidler, D.; Fuchs, E.; Braun, R.; Binder, E.M. Enzymatic degradation of ochratoxin A. In Proceedings of the X. Intern. IUPAC Symposium on Mycotoxins, Phytotoxins, Guaruja, Brazil; 2000. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Abrunhosa, L.; Paterson, R.R.M.; Venâncio, A. Biodegradation of Ochratoxin A for Food and Feed Decontamination. Toxins 2010, 2, 1078-1099. https://doi.org/10.3390/toxins2051078
Abrunhosa L, Paterson RRM, Venâncio A. Biodegradation of Ochratoxin A for Food and Feed Decontamination. Toxins. 2010; 2(5):1078-1099. https://doi.org/10.3390/toxins2051078
Chicago/Turabian StyleAbrunhosa, Luís, Robert R.M. Paterson, and Armando Venâncio. 2010. "Biodegradation of Ochratoxin A for Food and Feed Decontamination" Toxins 2, no. 5: 1078-1099. https://doi.org/10.3390/toxins2051078