Phytotoxins Produced by Fungi Associated with Grapevine Trunk Diseases


 , and
, and
Abstract
:1. Introduction
2. Eutypa Dieback

2.1. Toxins Produced by Eutypa lata


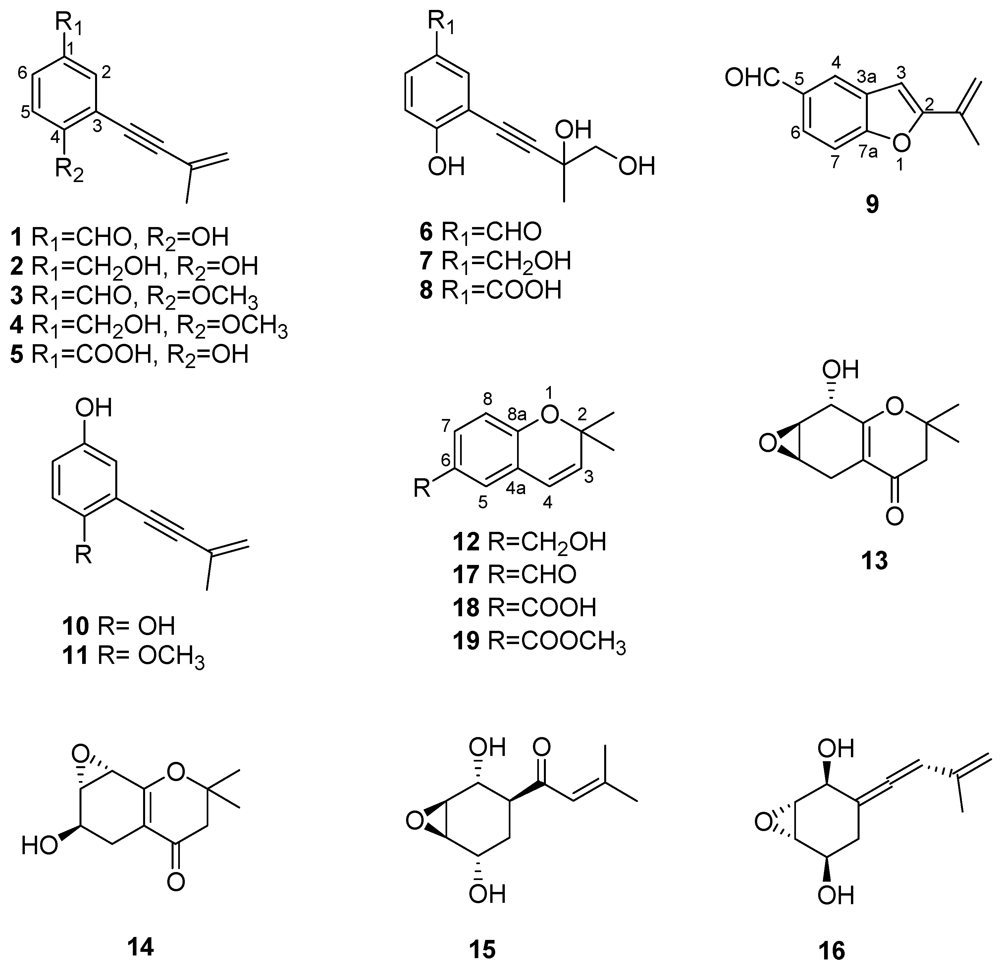
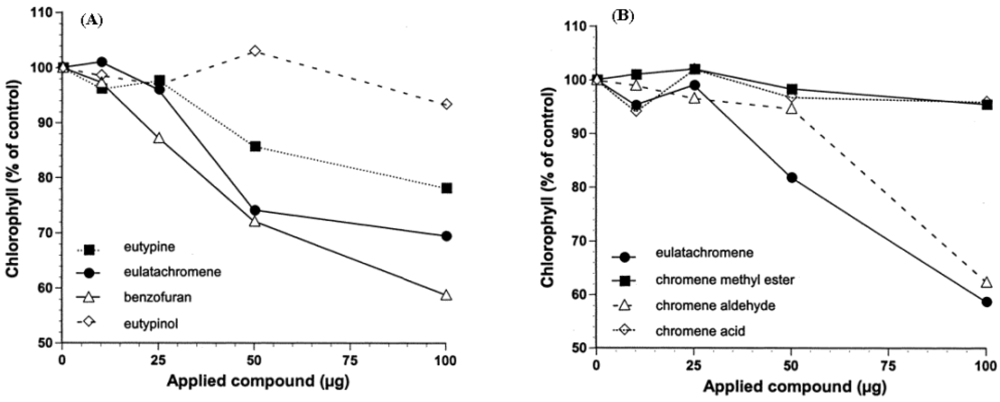
2.2. Structure-Toxicity Relationship Studies

2.3. Mechanism of Action of Eutypa lata Toxins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite (Figure 2) | Plant matrix used for phytotoxicity assays | References |
|---|---|---|
| Eutypine (1) | Grapevine leaves, tomato leaves, grapevine, mimosa and sugar beet membranes, mitochondria | [26,27,28,31,33,34,35] |
| Eutypinol (2) | Grapevine and tomato leaves | [26,27,28,31] |
| O-Methyleutypine (3) | Grapevine and tomato leaves, mitochondria | [26,34,35,36] |
| O-Methyleutypinol (4) | Grapevine leaves, tomato leaves | [26] |
| Eutypin carboxylic acid anologue (5) | Grapevine leaves, tomato leaves | [26] |
| 3-(3,4-Dihydroxy-3-methyl-1-butynyl)-4-hydroxy-benzaldehyde (6) | Grapevine leaves, tomato leaves | [26] |
| 2-(3,4-Dihydroxy-3-methyl-1-butynyl)-4-hydroxymethyl-phenol (7) | Grapevine and tomato leaves | [26] |
| 3-(3,4-Dihydroxy-3-methyl-1-butynyl)-4-hydroxy-benzoic acid (8) | Grapevine leaves, tomato leaves | [26] |
| 2-iso-Propenyl-5-formylbenzofuran (9) | Grapevine and tomato leaves | [26,28,31] |
| Siccayne (10) | Grapevine leaves | [28,31] |
| Eulatinol (11) | Grapevine leaves | [28] |
| Eulatachromene (12) | Grapevine leaves | [28,31] |
| Epoxidised chromanones (13, 14) | Grapevine leaves | [26] |
| Eutypoxide B (15) | Grapevine leaves | [29] |
| Allenic epoxycyclohexane (16) | Grapevine leaves | [29] |
2.4. Eutyta lata Toxins as Virulence Factors
2.5. Analytical Detection of Eutypa lata Toxins in Vitro and in Planta
3. Esca, A Complex of Diseases

3.1. Toxins Produced by Phaeomoniella chlamydospora and Phaeoacremonium aleophilum, Main Causal Agents of Leaf Stripe Disease
3.1.1. Lipophilic Low-Molecular Weight Metabolites


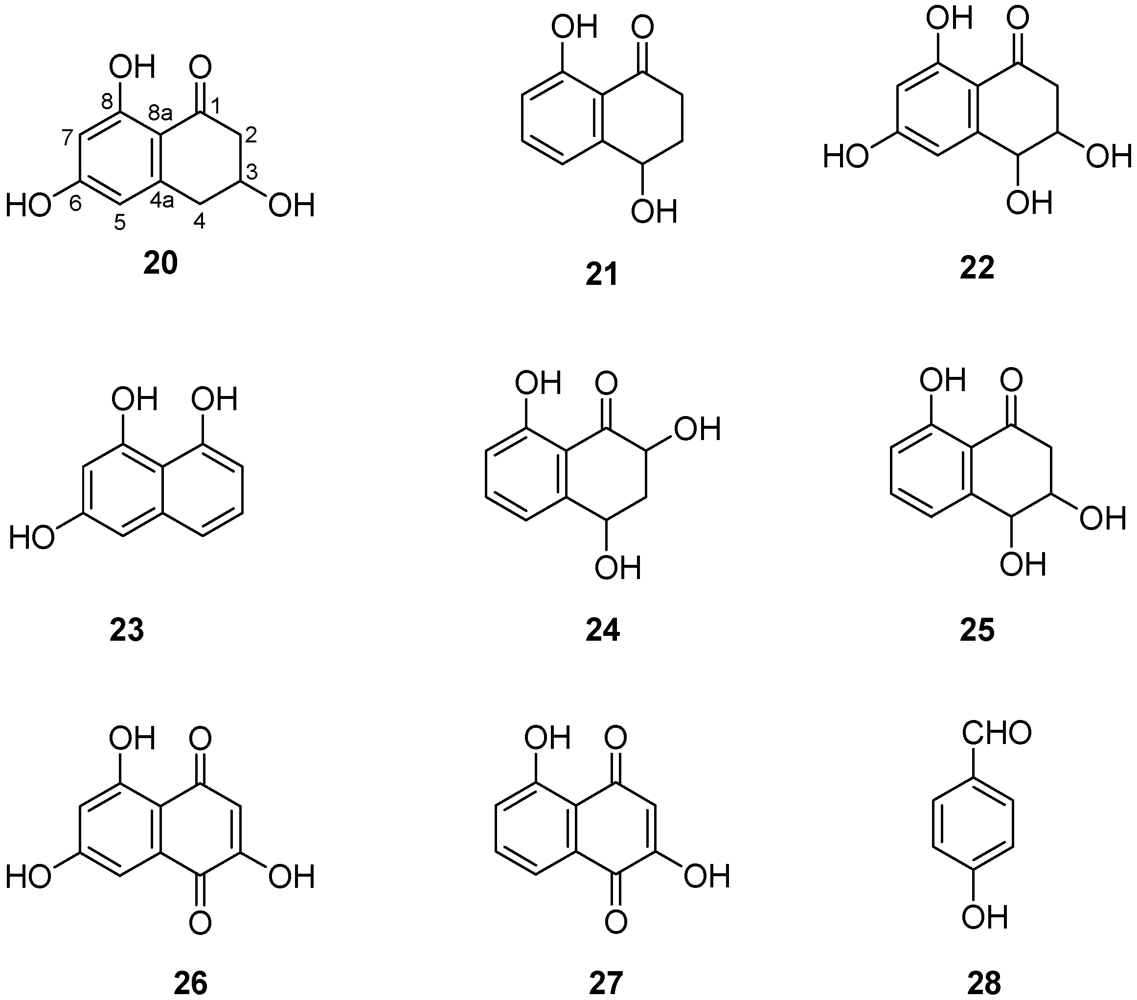
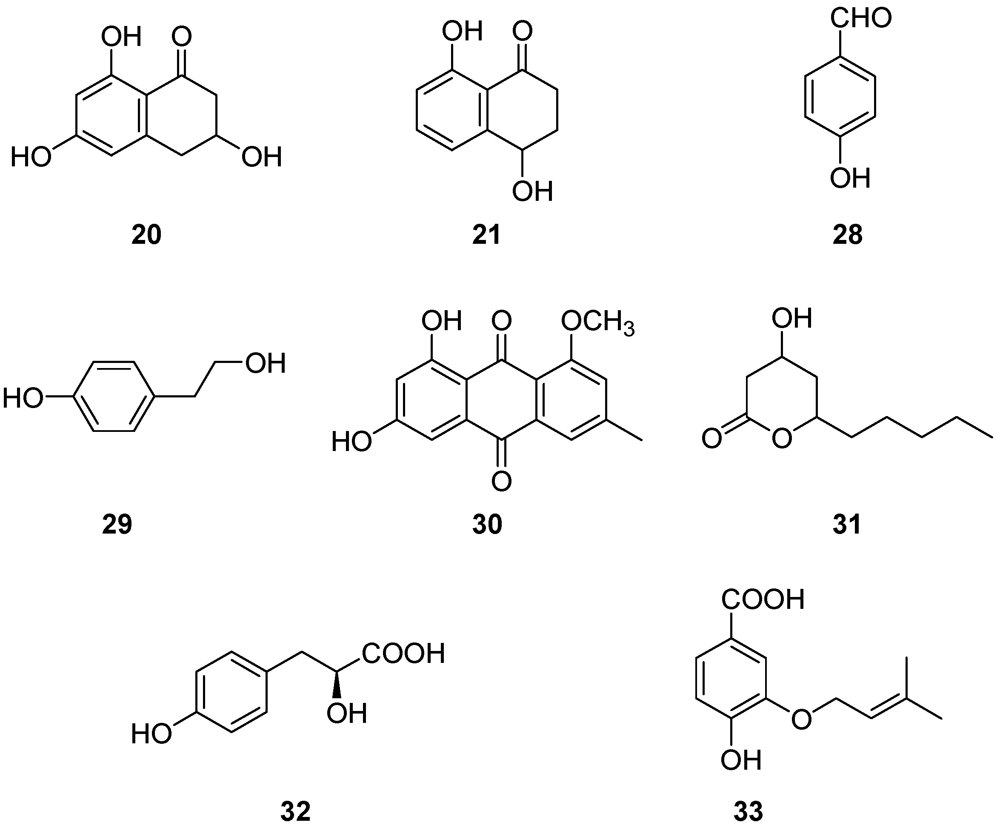
| Metabolite (Figure 6 and Figure 7) | Plant matrix used for phytotoxicity assays | References |
|---|---|---|
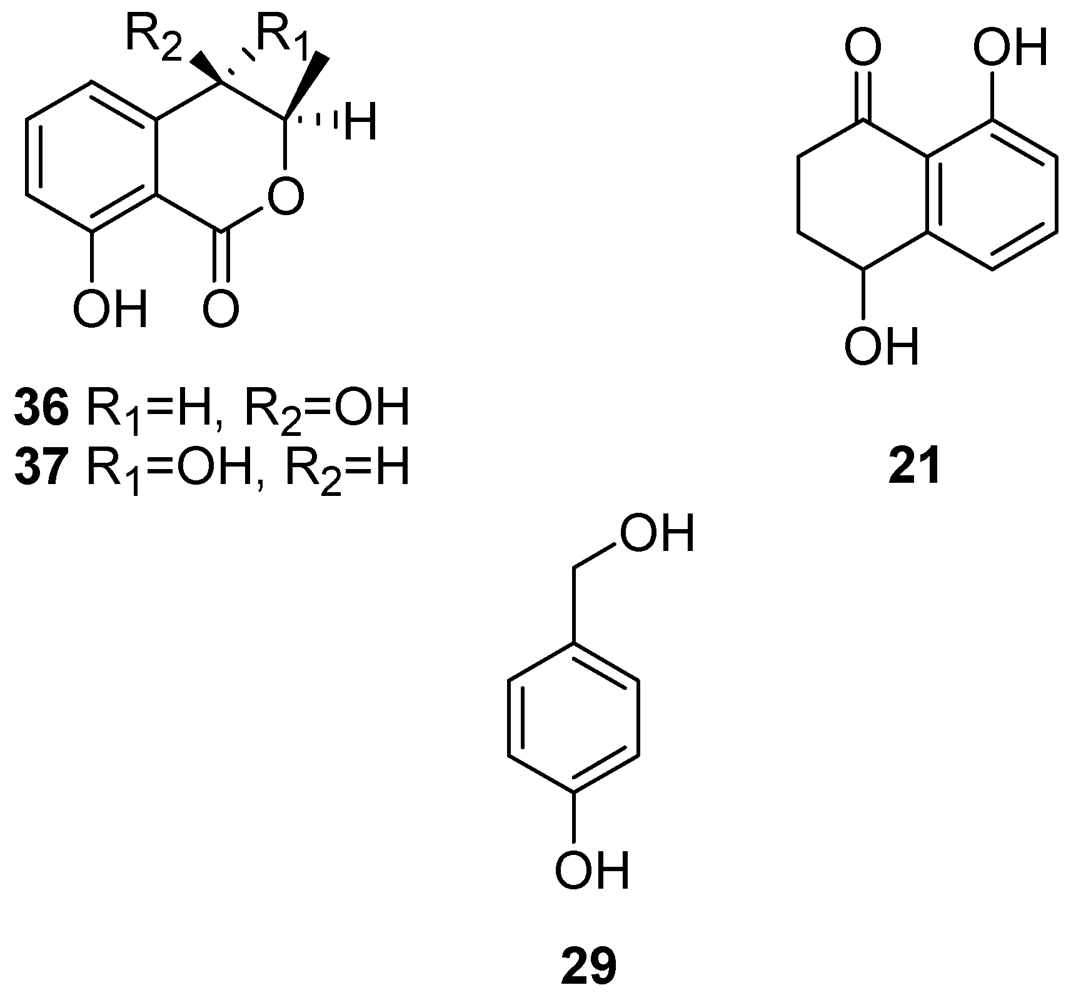
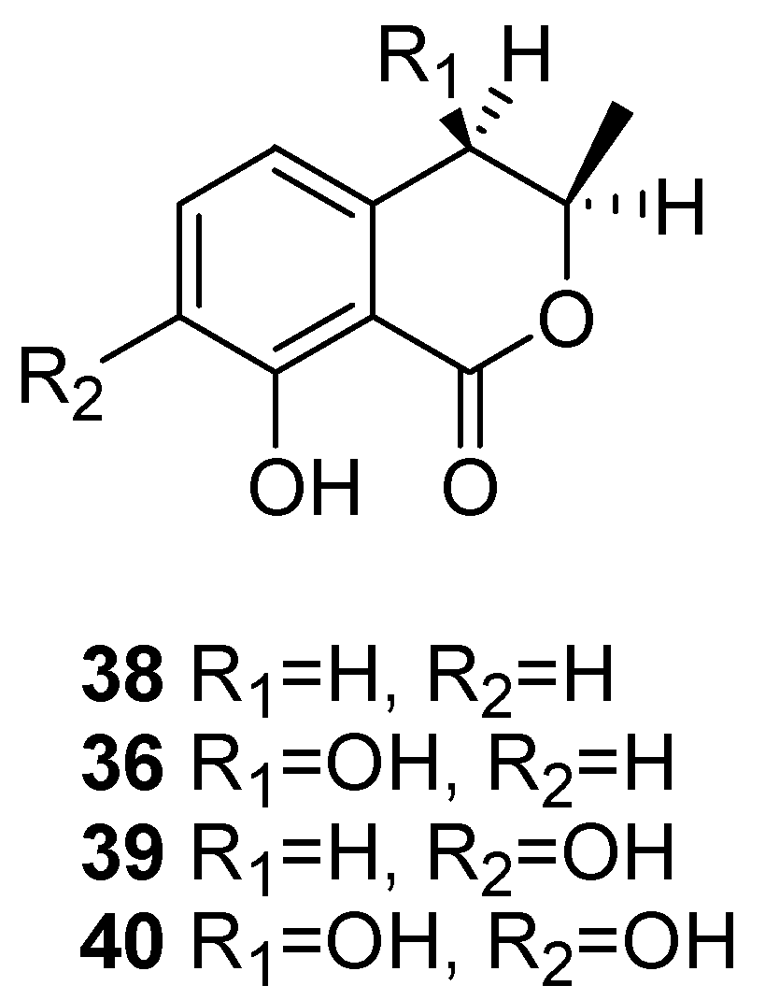
| Scytalone (20) | Grapevine leaves Grapevine callus and protoplasts Arabidopsis thaliana | [54,55,56,57] |
| Isosclerone (21) | Grapevine leaves Grapevine callus and protoplasts Arabidopsis thaliana | [54,55,56,57] |
| cis-4-Hydroxyscytalone (22) | Grapevine callus and protoplasts | [57] |
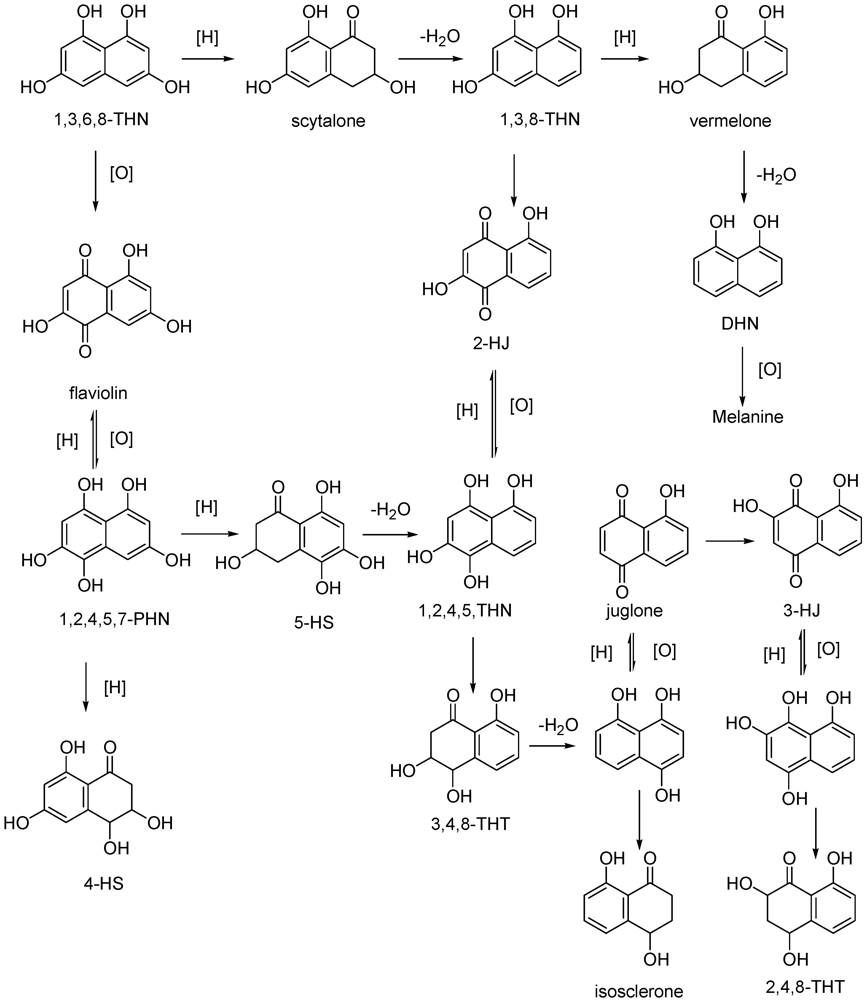
| 1,3,8-Trihydroxynaphthalene (1,3,8-THN) (23) | Grapevine callus | [57] |
| 2,4,8-Trihydroxy-tetralone (2,4,8-THT) (24) | Grapevine callus Arabidopsis thaliana | [57] |
| 3,4,8-Trihydroxytetralone (3,4,8-THT) (25) | Grapevine callus Arabidopsis thaliana | [57] |
| Flavioline (26) | Grapevine callus Arabidopsis thaliana | [55] |
| 2-Hydroyjuglone (27) | Grapevine callus | [57] |
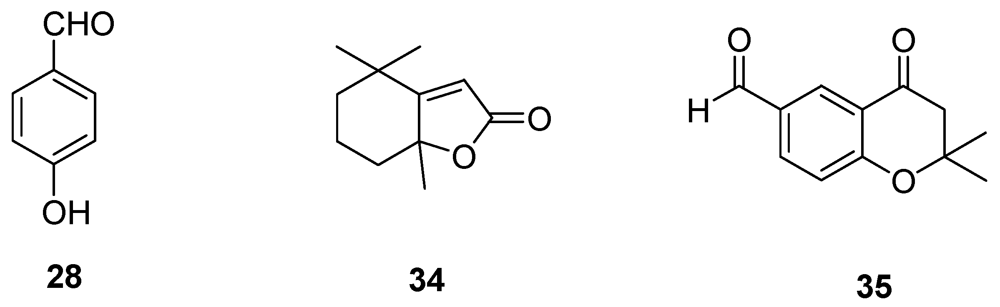
| 4-Hydroxybenzaldehyde (28) | Grapevine callus and protoplasts | [55,57] |
| (S)-4-Hydroxy-phenyllatic acid (32) | Grapevine callus and protoplasts | [55] |
| 3-(3-Methyl-2-butenyloxy)-4-hydroxybenzoic acid (33) | Grapevine callus and protoplasts | [55] |

3.1.2. Hydrophilic High-Molecular Weight Metabolites
3.1.2.1. Polysaccharides
3.1.2.2. Polypeptide Toxic Metabolites
3.2. Toxins Produced by Fomitiporia mediterranea, a Causal Agent of Wood Rot in Grapevine.

3.3. Mechanism of Action of Toxins Involved in the Esca Complex (Vascular Disease and Wood Rot)

3.4. Chemical and Immunological Detection of Esca-Related Metabolites
3.4.1. HPLC Technique
3.4.2. Immunological Techniques: Flow Cytometry


3.4.3. Immunological Techniques: Serological Test
3.5. NMR Metabolomics of Esca
4. Botryosphaeriaceae Species Involved in Grapevine Trunk Diseases

4.1. Phytotoxic Metabolites from Botryosphaeriaceae Species


4.2. Toxins Produced by Neofusicoccum parvum

| Metabolite | Concentration (M) | Toxicity rating a | Mean toxicity rating ± S.E. | ||
|---|---|---|---|---|---|
| Rep 1 | Rep 2 | Rep 3 | |||
| (3R,4R)-(-)-4-Hydroxymellein (36) | 0.26 × 10−3 | 2 | 2 | 3 | 2.3 ± 0.33 |
| 0.13 × 10−3 | 1 | 1 | 3 | 1.7 ± 0.67 | |
| 0.26 × 10−4 | 1 | 1 | 2 | 1.3 ± 0.33 | |
| (3R,4S)-(-)-4-Hydroxymellein (37) | 0.26 × 10−3 | 0 | 1 | 2 | 1.0 ± 0.58 |
| 0.13 × 10−3 | 0 | 1 | 1 | 0.7 ± 0.33 | |
| 0.26 × 10−4 | 0 | 1 | 0 | 0.3 ± 0.33 | |
| Isosclerone (21) | 0.31 × 10−3 | 2 | 3 | 1 | 2.0 ± 0.57 |
| Tyrosol (29) | 1.60 × 10−3 | 2 | 2 | 1 | 1.7 ± 0.33 |
| Control/Czapek-Dox | 0 | 0 | 0 | 0.0 ± 0.00 | |
| Control/H2O | 0 | 1 | 0 | 0.3 ± 0.33 | |
4.3. Toxins Produced by Diplodia Seriata

5. Grouping in Different Chemical Families of Phytoxins Produced by Different Fungi
6. Conclusions
References
- Carter, M.V. The Status of Eutypa Lata as a Pathogen. In Phytopathological Paper No. 32; CAB International: Wallingford, Oxon, UK, 1991. [Google Scholar]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (black measles) and brown wood-streaking: Two old and elusive diseases of grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar]
- Fischer, M. Biodiversity and geographic distribution of basidiomycetes causing esca-associated white rot in grapevine: A worldwide perspective. Phytopathol. Mediterr. 2006, 45, S30–S42. [Google Scholar]
- Halleen, F.; Fourie, P.H.; Crous, P.W. A review of black foot disease of grapevine. Phytopathol. Mediterr. 2006, 45, S55–S67. [Google Scholar]
- Surico, G.; Mugnai, L.; Marchi, G. Older and more recent observations on esca: A critical overview. Phytopathol. Mediterr. 2006, 45, S68–S86. [Google Scholar]
- Van Niekerk, J.M.; Fourie, P.H.; Halleen, F.; Crous, P.W. Botryosphaeria spp. as grapevine trunk pathogens. Phytopathol. Mediterr. 2006, 45, S43–S54. [Google Scholar]
- Mostert, L.; Halleen, F.; Fourie, P.; Crous, P.W. A review of Phaeoacremonium species involved in Petri disease and esca of grapevines. Phytopathol. Mediterr. 2006, 45, S12–S29. [Google Scholar]
- Trouillas, F.P.; Gubler, W.D. Pathogenicity of Diatrypaceae species in grapevines in California. Plant Dis. 2010, 94, 867–872. [Google Scholar]
- Trouillas, F.P.; Úrbez-Torres, J.R.; Gubler, W.D. Diversity of diatrypaceous fungi associated with grapevine canker diseases in California. Mycologia 2010, 102, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Lardner, R.; Mahoney, N.; Zanker, T.P.; Molyneux, R.J.; Scott, E.S. Secondary metabolite production by the fungal pathogen Eutypa lata: Analysis of the extracts from grapevine cultures and detection of those metabolites in planta. Aust. J. Grape Wine Res. 2006, 12, 107–114. [Google Scholar] [CrossRef]
- Halleen, F.; Mostert, L.; Crous, P.W. Pathogenicity testing of lesser-known vascular fungi of grapevines. Australas. Plant Pathol. 2007, 36, 277–285. [Google Scholar]
- Gramaje, D.; Mostert, L.; Armengol, J. Characterization of Cadophora luteo-olivacea and C. melinii isolates obtained from grapevines and environmental samples from grapevine nurseries in Spain. Phytopathol. Mediterr. 2011, 50, S112–S126. [Google Scholar]
- Sosnowski, M.R.; Wicks, T.W.; Scott, E.S. Control of Eutypa dieback in grapevines using remedial surgery Phytopathol. Mediterr. 2011, 50, S277–S284. [Google Scholar]
- Herche, R. Control Strategies for Trunk Diseases of Grapevine (Vitis vinifera L.). 2009, 56, M.Agr. Thesis, University of California-Davis, CA, USA. [Google Scholar]
- Rolshausen, P.E.; Gubler, W.D. Use of boron for the control of Eutypa dieback of grapevines. Plant Dis. 2005, 89, 734–738. [Google Scholar] [CrossRef]
- John, S.; Wicks, T.J.; Hunt, J.S.; Lorimer, M.F.; Oakey, H.; Scott, E.S. Protection of grapevine pruning wounds from infection by Eutypa lata using Trichoderma harzianum and Fusarium lateritium. Australas. Plant Pathol. 2005, 34, 569–575. [Google Scholar] [CrossRef]
- Bester, W.; Crous, P.W.; Fourie, P.H. Evaluation of fungicides as potential grapevine pruning wound protectants against Botryosphaeria species. Australas. Plant Pathol. 2007, 36, 73–77. [Google Scholar]
- Sosnowski, M.R.; Creaser, M.L.; Wicks, T.J.; Lardner, R.; Scott, E.S. Protection of grapevine pruning wounds from infection by Eutypa lata. Aust. J. Grape Wine Res. 2008, 14, 134–142. [Google Scholar] [CrossRef]
- Kotze, C.; Van Niekerk, J.; Mostert, L.; Halleen, F.; Fourie, P. Evaluation of biocontrol agents for grapevine pruning wound protection against trunk pathogen infection. Phytopathol. Mediterr. 2011, 50, S247–S263. [Google Scholar]
- Fourie, P.H.; Halleen, F. Proactive control of Petri disease of grapevine through treatment of propagation material. Plant Dis. 2004, 88, 1241–1245. [Google Scholar]
- Fourie, P.H.; Halleen, F. Chemical and biological protection of grapevine propagation material from trunk disease pathogens. Eur. J. Plant Pathol. 2006, 116, 255–265. [Google Scholar]
- Gramaje, D.; Armengol, J. Fungal trunk pathogens in the grapevine propagation process: Potential inoculum sources, detection, identification, and management strategies. Plant Dis. 2011, 95, 1040–1055. [Google Scholar] [CrossRef]
- Carter, M.V. Eutypa Dieback. In Compendium of Grape Diseases; Pearson, R.C., Goheen, A.C., Eds.; APS Press: St. Paul, MN, USA, 1988; pp. 32–34. [Google Scholar]
- Tsoupras, G.; de Angelis, P.; Zesiger, T.; Renoud, J.M.; Tabacchi, R. Biologically-active natural acetylenes from the culture medium of Eutypa lata. Bioact. Mol. 1988, 93–100. [Google Scholar]
- Renoud, J.M.; Tsoupras, G.; Stoeckli-Evans, H.; Tabacchi, R. A novel allenic epoxycyclohexane and related compounds from Eutypa lata (Pers:F.) Tul. Helv.Chim. Act 1989, 72, 1262–1267. [Google Scholar] [CrossRef]
- Renoud, J.M.; Tsoupras, G.; Tabacchi, R. Biologically active natural acetylenic compounds from Eutypa lata (Pers:F.) Tul. Helv. Chim. Acta 1989, 72, 929–932. [Google Scholar]
- Molyneux, R.J.; Mahoney, N.; Bayman, P.; Wong, R.Y.; Meyer, K.; Irelan, N. Eutypa dieback in grapevines: Differential production of acetylenic phenol metabolites by strains of Eutypa lata. J. Agric. Food Chem. 2002, 50, 1393–1399. [Google Scholar] [PubMed]
- Mahoney, N.; Lardner, R.; Molyneux, R.J.; Scott, E.S.; Smith, L.R.; Schoch, T.K. Phenolic and heterocyclic metabolite profiles of the grapevine pathogen Eutypa lata. Phytochemistry 2003, 64, 475–484. [Google Scholar] [PubMed]
- Defrancq, E.; Gordon, J.; Brodard, A.; Tabacchi, R. The synthesis of a novel epoxycyclohexane from the fungus Eutypa lata (Pers:F) Tul. Helv. Chim. Acta 1992, 75, 276–281. [Google Scholar] [CrossRef]
- Chapuis, L.; Corio-Costet, M.F.; Malosse, C. Sterol composition of the woody plant pathogenic fungus Eutypa lata. Phytochemistry 1996, 42, 1599–1601. [Google Scholar] [CrossRef]
- Smith, L.R.; Mahoney, N.; Molyneux, R.J. Synthesis and structure-phytotoxicity relationships of acetylenic phenols and chromene metabolites, and their analogues, from the grapevine pathogen Eutypa lata. J. Nat. Prod. 2003, 66, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Mahoney, N.; Chan, K.L.; Molyneux, R.J.; Campbell, B.C. Secondary metabolites of the grapevine pathogen Eutypa lata inhibit mitochondrial respiration, based on a model bioassay using the yeast Saccharomyces cerevisiae. Curr. Microbiol. 2004, 49, 282–287. [Google Scholar] [PubMed]
- Amborabé, B.E.; Fleurat-Lessard, P.; Bonmort, J.; Roustan, J.P.; Roblin, G. Effects of eutypine, a toxin from Eutypa lata, on plant cell plasma membrane: Possible subsequent implication in disease development. Plant Physiol. Biochem. (Paris) 2001, 39, 51–58. [Google Scholar]
- Deswarte, C.; Eychenne, J.; Davy de Virville, J.; Roustan, J.; Moreau, F.; Fallot, J. Protonophoric activity of eutypine, a toxin from Eutypa lata, in plant mitochondri. Arch. Biochem. Biophys. 1996, 334, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Deswarte, C.; Canut, H.; Klaebe, A.; Roustain, J.P.; Fallot, J. Transport, cytoplasmatic accumulation, and mechanism of action of the toxic eutypine in Vitis vinifera cell. J. Plant Pathol. 1996, 149, 336–342. [Google Scholar]
- Fallot, J.; Deswarte, C.; Dalmayrac, S.; Colrat, S.; Roustan, J.P. L’Eutypiose de la vigne: Isolement d’une molécule synthétisée par Eutypa lata et toxique pour la vigne. Comptes Rendus Acad. Sci. Ser. III 1997, 320, 149–158. [Google Scholar]
- Colrat, S.; Deswarte, C.; Latché, A.; Klaebe, A.; Bouzayen, M.; Fallot, J.; Roustan, J.P. Enzymic detoxification of eutypine, a toxin from Eutypa lata, by Vitis vinifera cells. Partial purification of an NADPH-dependent aldehyde reductase. Planta 1999, 207, 544–550. [Google Scholar] [CrossRef]
- Afifi, M.; Monje, M.C.; Legrand, V.; Roustan, J.P. Metabolisation of eutypine by plant tissues: An HPLC determination. Anal. Chim. Acta 2004, 513, 21–27. [Google Scholar]
- Colrat, S.; Latché, A.; Guis, M.; Pech, J.C.; Bouzayen, M.; Fallot, J.; Roustan, J.P. Purification and characterization of a NADPH-dependent aldehyde reductase from mung bean that detoxifies eutypine, a toxin from Eutypa lata. Plant Physiol. 1999, 119, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Legrand, V.; Dalmayrac, S.; Latché, A.; Pech, J.C.; Bouzayen, M.; Fallot, J.; Torregrosa, L.; Bouquet, A.; Roustan, J.P. Constitutive expression of Vr-ERE gene in transformed grapevines confers enhanced resistance to eutypine, a toxin from Eutypa lata. Plant Sci. 2003, 164, 809–814. [Google Scholar] [CrossRef]
- Guillen, P.; Guis, M.; Martínez-Reina, G.; Colrat, S.; Dalmayrac, S.; Deswarte, C.; Bouzayen, M.; Roustan, J.P.; Fallot, J.; Pech, J.C.; et al. A novel NADPH-dependent aldehyde reductase gene from Vigna radiata confers resistance to the grapevine fungal toxin eutypine. Plant J. 1998, 16, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Lecomte, P.; Péros, J.P.; Blancard, D.; Bastien, N.; Délye, C. PCR assays that identify the grapevine dieback fungus Eutypa lata. Appl. Environ. Microbiol. 2000, 66, 4475–4480. [Google Scholar] [PubMed]
- Lardner, R.; Stummer, B.E.; Sosnowski, M.R.; Scott, E.S. Molecular identification and detection of Eutypa lata in grapevine. Mycol. Res. 2005, 109, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Sosnowski, M.R.; Shtienberg, D.; Creaser, M.L.; Wicks, T.J.; Lardner, R.; Scott, E.S. The influence of climate on foliar symptoms of eutypa dieback in grapevines. Phytopathology 2007, 97, 1284–1289. [Google Scholar] [PubMed]
- Viala, P. Recherches sur les maladies de la vigne. Esca. Annales des Epiphyties 1926, 12, 5–108. [Google Scholar]
- Marsais, P. Maladie de l’esca. Rev. de Vit. 1923, 59, 8–14. [Google Scholar]
- Surico, G. Towards a redefinition of the diseases within the esca complex of grapevine. Phytopathol. Mediterr. 2009, 48, S487–S490. [Google Scholar]
- Mostert, L.; Groenewald, J.Z.; Summerbell, R.C.; Gams, W.; Crous, P.W. Taxonomy and pathology of Togninia (Diaporthales) and its Phaeoacremonium anamorphs. Stud. Mycol. 2006, 54, 1–115. [Google Scholar] [CrossRef] [Green Version]
- Essakhi, S.; Mugnai, L.; Crous, P.W.; Groenewald, J.Z.; Surico, G. Molecular and phenotypic characterisation of novel Phaeoacremonium species isolated from esca diseased grapevines. Persoonia 2008, 21, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Graham, A.B.; Johnston, P.R.; Weir, B.S. Three new Phaeoacremonium species on grapevines in New Zealand. Australas. Plant Pathol. 2009, 38, 505–513. [Google Scholar] [CrossRef]
- Gramaje, D.; Armengol, J.; Mohammadi, H.; Banihashemi, Z.; Mostert, L. Novel Phaeoacremonium species associated with Petri disease and esca of grapevine in Iran and Spain. Mycologia 2009, 101, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Surico, G.; Mugnai, L.; Marchi, G. The Esca Disease Complex. In Integrated Management of Diseases Caused by Fungi, Phytoplasma and Bacteria; Springer: Heidelberg, Germany, 2008; pp. 119–136. [Google Scholar]
- Luque, J.; Martos, S.; Aroca, A.; Raposo, R.; Garcia-Figueres, F. Symptoms and fungi associated with declining mature grapevine plants in northeast Spain. J. Plant Pathol. 2009, 91, 381–390. [Google Scholar]
- Evidente, A.; Sparapano, L.; Andolfi, A.; Bruno, G. Two naphthalenone pentaketides from liquid cultures of Phaeoacremonium aleophilum, a fungus associated with esca of grapevine. Phytopathol. Mediterr. 2000, 39, 162–168. [Google Scholar]
- Tabacchi, R.; Fkyerat, A.; Poliart, C.; Dubin, G.M. Phytotoxins from fungi of esca of grapevine. Phytopathol. Mediterr. 2000, 39, 156–161. [Google Scholar]
- Evidente, A.; Peduto, F.; Andolfi, A.; Marchi, G.; Mugnai, L.; Surico, G. Fattori di Virulenza dei Funghi Dell’esca. Fitotossine da Phaeomoniella chlamydospora, Phaeoacremonium aleophilum e Fomitiporia mediterranea: Produzione e Attività Biologica. In Il Mal dell’Esca della Vite: Interventi di ricerca e Sperimentazione per il Contenimento Della Malattia. Progetto MesVit; Surico, G., Mugnai, L., Eds.; Arsia Regione Toscana: Firenze, Italy, 2010; pp. 214–231. [Google Scholar]
- Abou-Mansour, E.; Couche, E.; Tabacchi, R. Do fungal naphthalenones have a role in the development of esca symptoms? Phytopathol. Mediterr. 2004, 43, 75–82. [Google Scholar]
- Sparapano, L.; Bruno, G.; Graniti, A. Effects on plants of metabolites produced in culture by Phaeoacremonium chlamydosporum, P. aleophilum and Fomitiporia punctata. Phytopathol. Mediterr. 2000, 39, 169–177. [Google Scholar]
- Seviour, T.; Lambert, L.K.; Pijuan, M.; Yuan, Z. Structural determination of a key exopolysaccharide in mixed culture aerobic sludge granules using NMR spectroscopy. Environ. Sci. Technol. 2010, 44, 8964–8970. [Google Scholar] [PubMed]
- Hogdson, R.; Peterson, W.H.; Riker, A.J. The toxicity of polysaccharides and other large molecules to tomato cuttings. Phytopathology 1949, 39, 47–62. [Google Scholar]
- Harborne, J.B. Toxins of Plant-Fungal. In Interaction. In Handbook of Natural Toxins: Plant and Fungal Toxins; Keelr, R.F., Tu, A., Eds.; Marcel Dekker: New York, NY, USA, 1983. [Google Scholar]
- Denny, T.P. Involvement of bacterial polysaccharides in plant pathogenesis. Ann. Rev. Phytopathol. 1995, 33, 173–197. [Google Scholar]
- van Alfen, N.K. Reassessment of plant wilt toxins. Ann. Rev. Phytopathol. 1989, 27, 533–550. [Google Scholar]
- Spalding, D.H.; Bruehl, G.W.; Foster, R.J. Possible role of pectinolytic enzymes and polysaccharide in pathogenesis by Cephalosporium gramineum in wheat. Phytopathology 1961, 51, 227–235. [Google Scholar]
- McWain, P.; Gregory, G.F. A neutral mannan from Ceratocystis fagacearum culture filtrate. Phytochemistry 1972, 11, 2609–2612. [Google Scholar] [CrossRef]
- Strobel, G.A.; van Alfen, N.; Hapner, K.D.; McNeil, M.; Albersheim, P. Some phytotoxic glycopeptides from Ceratocystis ulmi, the Dutch Elm Disease pathogen. Biochim. Biophys. Acta 1978, 538, 60–75. [Google Scholar] [PubMed]
- Thomas, C.A. A wilt-inducing polysaccharide from Fusarium solani f. eumartii. Phytopathology 1949, 39, 572–579. [Google Scholar]
- Keen, N.T.; Wang, M.C.; Bartnicki-Garcia, S.; Zentmeyer, G.A. Phytotoxicity of myco-laminarans-β-l,3-glucans from Phytophthora spp. Physiol. Plant Pathol. 1975, 7, 91–97. [Google Scholar]
- Woodward, J.R.; Keane, P.J.; Stone, B.A. Structures and properties of wilt-inducing polysaccharides from Phytophthora spp. Physiol. Plant Pathol. 1980, 16, 439–454. [Google Scholar]
- Barbosa, A.M.; Steluti, R.M.; Dekker, R.F.H.; Cardoso, M.S.; Corradi da Silva, M.L. Structural characterization of Botryosphaeran: A (1→3;1→6)-β-D-glucan produced by the ascomyceteous fungus, Botryosphaeria sp. Carbohyd. Res. 2003, 338, 1691–1698. [Google Scholar] [CrossRef]
- Rudolph, K.W.E.; Gross, M.; Neugebauer, M.; Hokawat, S.; Zachowski, A.; Wanddra, K.; Klement, Z. Extracellular Polysaccharides as Determinants of Leaf Spot Diseases Caused by Pseudomonads and Xanthomonads. In Phytotoxins and Plant Pathogenesis, NATO ASI Series, Series H: Cell Biology; Graniti, A., Durbin, R.D., Ballio, A., Eds.; Springer Verlag: Berlin, Germany, 1989; pp. 177–218. [Google Scholar]
- Corsaro, M.M.; de Castro, C.; Evidente, A.; Lanzetta, R.; Molinaro, A.; Parrilli, M.; Sparapano, L. Phytotoxic extracellular polysaccharide fractions from Cryphonectria parasitica (Murr.). Barr. Carbohyd. Polym. 1998, 37, 167–172. [Google Scholar] [CrossRef]
- Corsaro, M.M.; de Castro, C.; Evidente, A.; Lanzetta, R.; Molinaro, A.; Mugnai, L.; Parrilli, M.; Surico, G. Chemical structure of two phytotoxic exopolysaccharides produced by Phomopsis foeniculi. Carbohydr. Res. 1998, 308, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Martos, S.; Andolfi, A.; Luque, J.; Mugnai, L.; Surico, G.; Evidente, A. Production of phytotoxic metabolites by five species of Botryosphaeriaceae causing decline on grapevine, with special interest in the species Neofusicoccum luteum and N. parvum. Eur. J. Plant Pathol. 2008, 121, 451–461. [Google Scholar] [CrossRef]
- Leathers, T.D. Pullulan. In Polysaccharides II: Polysaccharides from Eukaryotes; Vandamme, E.J., de Baets, S., Steinbuchel, A., Eds.; Wiley-VCH: Weinheim, Germany, 2002. [Google Scholar]
- Campbell, B.S.; Siddique, A.B.M.; McDougall, B.M.; Seviour, R.J. Which morphological forms of the fungus Aureobasidium pullulans are responsible for pullulan production? FEMS Microb. Lett. 2004, 232, 225–228. [Google Scholar] [CrossRef]
- Bruno, G.; Sparapano, L.; Graniti, A.A. Effects of three esca-associated fungi on Vitis vinifera L.: IV. Diffusion through the xylem of metabolites produced by two tracheiphilous fungi in the woody tissue of grapevine leads to esca-like symptoms on leaves and berries. Physiol. Mol. Plant Pathol 2007, 71, 106–124. [Google Scholar] [CrossRef]
- Luini, E.; Fleurat-Lessard, P.; Rousseau, L.; Roblin, G.; Berjeaud, J. Inhibitory effects of polypeptides secreted by the grapevine pathogens Phaeomoniella chlamydospora and Phaeoacremonium aleophilum on plant cell activities. Physiol. Mol. Plant Pathol. 2010, 74, 403–411. [Google Scholar] [CrossRef]
- Fleurat-Lessard, P.; Luini, E.; Berjeaud, J.M.; Roblin, G. Diagnosis of grapevine esca disease by immunological detection of Phaeomoniella chlamydospora. Aust. J. Grape Wine Res. 2010, 16, 455–463. [Google Scholar] [CrossRef]
- White, C.L. The Characterization of the Basidiomycetes and Other Fungi Associated with Esca of Grapevines in South Africa. 2010. [Google Scholar]
- Castro, F.A.V.; Mariani, D.; Panek, A.D.; Eleutherio, E.C.A.; Pereira, M.D. Cytotoxicity mechanism of two naphthoquinones (menadione and plumbagin) in Saccharomyces cerevisiae. PLoS One 2008, 3. [Google Scholar] [CrossRef]
- Butler, M.; Gardiner, R.; Day, A. Melanin synthesis by Sclerotinia sclerotiorum. Mycologia 2009, 101, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, M.H.; Abramczyk, D.; Puckhaber, L.S.; Naruse, M.; Ebizuka, Y.; Fujii, I.; Szaniszlo, P.J. New biosynthetic step in the melanin pathway of Wangiella (Exophiala) dermatitidis: Evidence for 2-acetyl-1,3,6,8-tetrahydroxynaphthalene as a novel precursor. Eukaryot. Cell 2008, 7, 1699–1711. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Polacheck, I. Melanin-lacking mutants of Cryptococcus neoformans and their virulence for mice. J. Bacteriol. 1982, 150, 1414–1421. [Google Scholar] [PubMed]
- Geis, P.A.; Wheeler, M.H.; Szaniszlo, P.J. Pentaketide metabolites of melanin synthesis in the dematiaceous fungus Wangiella dermatitidis. Arch. Microbiol. 1984, 137, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.M.; Szaniszlo, P.J.; Polak, A. Dihydroxynaphthalene (DHN) Melanin and its Relationship with Virulence in the Early Stages of Phaeohyphomycosis. In The Fungal Spore and Disease Initiation in Plants and Animals; Cole, G.T., Hock, H.C., Eds.; Plenum Press: New York, NY, USA, 1991; pp. 297–318. [Google Scholar]
- Durrell, L.W. The composition and structure of walls of dark fungus spores. Mycopathol. Mycol. Appl. 1964, 23, 339–345. [Google Scholar]
- Bürki, N.; Michel, A.; Tabacchi, R. Naphthalenones and isocoumarins of the fungus Ceratocystis fimbriata sp. platani. Phytopathol. Mediterr. 2003, 42, 191–198. [Google Scholar]
- Oberth, C.H.; Jones, A.D.; Shibamoto, T. Retro-Michael Fragmentation in Tandem Mass Spectrometry of Modified Peptides. In Proceedings of the 40th ASMS Conference on Mass SpectrometryWashington, DC, USA, 31 May-5 June 1992; p. 1715.
- Scandalios, J.G. Response of plant antioxidant defense genes to environment stress. Adv. Genet. 1990, 28, 2–35. [Google Scholar]
- Dangl, J.L.; Deetrich, R.A.; Richberg, M.H. Death don’t have no mercy: Cell death programs in plant-microbe interactions. Plant Cell 1996, 8, 1793–1807. [Google Scholar]
- Thomson, R.H. Naturally Occuring Quinones; Academic Press: London, UK, 1971; pp. 39–92. [Google Scholar]
- Medentsev, A.G.; Akimenko, V.K. Naphthoquinone metabolites of the fungi. Phytochemistry 1998, 47, 933–959. [Google Scholar]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of Phenolics in the Resistance Mechanisms of Plants Against Fungal Pathogens and insects. In Phytochemistry: Advances in Research; Imperato, F., Ed.; Research Signpost: Kerala, India, 2006; pp. 23–67. [Google Scholar]
- Kurahashi, Y.; Sakawa, S.; Kinbara, T.; Tanaka, K.; Kagabu, S. Biological activity of carpropamid (KTU 3616): A new fungicide for rice blast disease. J. Pest. Sci. 1997, 22, 108–112. [Google Scholar]
- Yumita, T.; Shojiand, A.; Yamamoto, I. Metabolism of mepronil (basitac) in rice plants. J. Pest. Sci. 1983, 5, 347–349. [Google Scholar]
- Petit, A.N.; Vaillant, N.; Boulay, M.; Clement, C.; Fontaine, F. Alteration of photosynthesis in grapevines affected by esca. Phytopathology 2006, 96, 1060–1066. [Google Scholar]
- Mattii, G.B.; Bardi, E.; Calabrese, C.; Giorgini, A. Alterazioni Degli Scambi Gassosi Causati dal Mal Dell’esca-Misurazioni su Foglia Singola e SULL’INTERA Chioma di Piante di Vite in Vaso. 2010. In Il mal Dell’esca Della Vite: Interventi di Ricerca e Sperimentazione per il Contenimento Della Malattia.Progetto MesVit; Surico, G., Mugnai, L., Eds.; Arsia Regione Toscana: Firenze, Italy, 2010; pp. 241–260. [Google Scholar]
- Calamai, L.; Mattii, G.; Mugnai, L.; Surico, G. Physiological alterations in esca-diseased vines and the detection of scytalone and isosclerone in the tiger-striped vine leaves. Phytopathol. Mediterr. 2009, 48, 168–169. [Google Scholar]
- Calamai, L.; Zini, E.; Cantalupo, L.; Comparini, C. Monitoraggio di Marcatori Chimici di Stress Idrico in Foglie di Vite Con Sintomi di Esca. In Il Mal dell’esca Della Vite: Interventi di Ricerca e Sperimentazione per il Contenimento Della Malattia.Progetto MesVit; Surico, G., Mugnai, L., Eds.; Arsia Regione Toscana: Firenze, Italy, 2010; pp. 232–240. [Google Scholar]
- Dimond, A.E.; Waggoner, P.E. On the nature and role of vivotoxins in plant diseases. Phytopathology 1953, 43, 229–235. [Google Scholar]
- Bruno, G.; Sparapano, L. Effects of three esca-associated fungi on Vitis vinifera L. II.Characterization of biomolecules in xylem sap and leaves of healthy and diseased vines. Physiol. Mol. Plant Pathology 2007, 69, 195–208. [Google Scholar]
- Andolfi, A.; Cimmino, A.; Evidente, A.; Iannacone, M.; Capparelli, R.; Mugnai, L.; Surico, G. A new flow cytometry technique to identify Phaeomoniella chlamydospora exopolysaccharides and study mechanisms of esca grapevine foliar symptoms. Plant Dis. 2009, 93, 680–684. [Google Scholar] [CrossRef]
- Santos, C.; Fragoeiro, S.; Valentim, H.; Phillips, A. Phenotypic characterisation of Phaeacremoniumand Phaemoniella strains isolated from grapevines: Enzyme production and virulence of extra-cellular filtrate on grapevine callus. Sci. Hortic. 2006, 107, 123–130. [Google Scholar]
- Bruno, G.; Sparapano, L. Effects of three esca associated fungi on Vitis vinifera L.: III. Enzymes produced by the pathogens and their role in fungus-to-plant or in fungus-to-fungus interactions. Physiol. Mol. Plant Pathol. 2006, 69, 182–194. [Google Scholar] [CrossRef]
- Lima, M.R.M.; Felgueiras, M.L.; Graça, G.; João, E.A.R.; Barros, A.; Gil, G.; Dias, A.C.P. NMR metabolomics of esca disease-affected Vitis vinifera cv. Alvarinho leaves. J. Exp. Bot. 2010, 61, 4033–4042. [Google Scholar] [CrossRef] [PubMed]
- Úrbez-Torres, J.R.; Gubler, W.D.; Luque, J. First report of Botryosphaeria iberica and Botryosphaeria viticola associated with grapevine decline in California. Plant Dis. 2007, 91. [Google Scholar] [CrossRef]
- rbez-Torres, J.R. The status of Botryosphaeriaceae species infecting grapevines. Phytopathol. Mediterr. 2011, 50, S5–S45. [Google Scholar]
- Hewitt, R.D. Diplodia Cane Dieback and Bunch Ro. In Compendium of Grape Diseases; The American Phytopathological Society (APS) Press: St. Paul, MN, USA, 1988; pp. 25–26. [Google Scholar]
- Phillips, A.J.L. Botryosphaeria dothidea and other fungi associated with excoriose and dieback of grapevines in Portugal. J. Phytopathol. 1998, 146, 327–332. [Google Scholar] [CrossRef]
- Phillips, A.J.L. Botryosphaeria species associated with diseases of grapevines in Portugal. Phytopathol. Mediterr. 2002, 41, 3–18. [Google Scholar]
- Larignon, P.; Fulchic, R.; Cere, L.; Dubos, B. Observation on black dead arm in French vineyards. Phytopathol. Mediterr. 2001, 40, S336–S342. [Google Scholar]
- Castillo-Pando, M.; Somers, A.; Green, C.D.; Priest, M.; Sriskanthades, M. Fungi associated with dieback of Semillon grapevines in the Hunter Valley of New South Wales. Australas. Plant Pathol. 2001, 30, 59–63. [Google Scholar]
- Taylor, A.; Hardy, G.E.S.; Wood, P.; Burgess, T. Identification and pathogenicity of Botryosphaeria species associated with grapevine decline in Western Australia. Australas. Plant Pathol. 2005, 34, 187–195. [Google Scholar] [CrossRef]
- Wunderlich, N.; Steel, C.C.; Ash, G.; Raman, H.; Savocchia, S. Identification of Botryosphaeria spp. and First Report of Dothiorella Viticola (“Botryosphaeria”Viticola) Associated with Bunch Rot in Australia. In Proceedings of the 6th International Workshop on Grapevine Trunk Diseases, Florence, Italy, 1-3 September 2008.
- Savocchia, S.; Steel, C.C.; Stodart, B.J.; Somers, A. Pathogenicity of Botryosphaeria species isolated from declining grapevines in sub-tropical regions of eastern Australia. Vitis 2007, 46, 27–32. [Google Scholar]
- Corradi da Silva, M.L.; Izeli, N.L.; Martinez, P.F.; Silva, I.R.; Costantino, C.J.L.; Cardoso, M.S.; Barbosa, A.M.; Dekker, R.F.H.; da Silva, G.V.J. Purification and structural characterisation of (1→3;1→6)-β-D-glucans (botryosphaerans) from Botryosphaeria rhodina grown on sucrose and fructose as carbon sources. a comparative study. Carbohyd. Polym. 2005, 61, 10–17. [Google Scholar] [CrossRef]
- Evidente, A.; Motta, A. Phytotoxins from Fungi Pathogenic for Agrarian, Forestal and Weedy Plants. In Bioactive Compounds from Natural Sources: Isolation, Characterization, and Biological Properties; Taylor & Francis Group: London, UK, 2001; pp. 473–525. [Google Scholar]
- Evidente, A.; Punzo, B.; Andolfi, A.; Cimmino, A.; Melck, D.; Luque, J. Lipophilic phytotoxins produced by Neofusicoccum parvum, a grapevine canker agent. Phytopathol. Mediterr. 2010, 49, 74–79. [Google Scholar]
- Aldridge, D.C.; Galt, S.; Giles, D.; Turner, W.B. Metabolites of Lasiodiplodia theobromae. J. Chem. Soc. 1971, 1623–1627. [Google Scholar]
- Kimura, Y.; Tamura, S. Isolation of l-β-phenyllactic acid and tyrosol as plant-growth regulators from Gloeosporium laeticolor. Agric. Biol. Chem. 1973, 37, 2925–2925. [Google Scholar] [CrossRef]
- Morita, T.; Aoki, H. Isosclerone, a new metabolite of Sclerotinia sclerotiorum (Lib.) De Bary. Agric. Biol. Chem. 1974, 38, 1501–1504. [Google Scholar] [CrossRef]
- Cole, R.J.; Cox, R.H. Handbook of Toxic Fungal Metabolites; Academic Press: New York, NY, USA, 1981. [Google Scholar]
- Turner, W.B.; Aldridge, D.C. Fungal Metabolites II; Academic Press: London, UK, 1983; pp. 82–109. [Google Scholar]
- Venkatasubbaiah, P.; Chilton, W.S. Phytotoxins of Botryosphaeria obtusa. J. Nat. Prod. 1990, 53, 1628–1630. [Google Scholar] [CrossRef]
- Devys, M.; Barbier, M.; Bousquet, J.F.; Kollmann, A. Isolation of the new (-)-(3R,4S)-4-hydroxymellein from the fungus Septoria nodorum Berk. Z. Naturforsch. B. 1992, 47, 779–881. [Google Scholar]
- Cabras, A.; Mannoni, M.A.; Serra, S.; Andolfi, A.; Fiore, M. Occurrence, isolation and biological activity of phytotoxic metabolites produced in vitro by Sphaeropsis sapinea, pathogenic of Pinus radiata. Eur. J. Plant Pathol. 2006, 115, 187–193. [Google Scholar] [CrossRef]
- Venkatasubbaiah, P.; Sutton, T.B.; Chilton, W.S. Effect of phytotoxins produced by Botryosphaeria obtusa, the cause of black rot of apple fruit and frogeye leaf spot. Phytopathology 1991, 81, 243–247. [Google Scholar] [CrossRef]
- Lecomte, P.; Leyo, M.; Louvet, G.; Corio-Costet, M.F.; Gaudillère, J.P.; Blancard, D. Le Black Dead Arm, genèse des symptômes. Phytoma La Défense des Végetaux 2005, 587, 29–37. [Google Scholar]
- Djoukeng, J.D.; Polli, S.; Larignon, P.; Mansour, E.A. Identification of phytotoxins from Botryosphaeria obtusa, a pathogen of black dead arm disease of grapevine. Eur. J. Plant Pathol. 2009, 124, 303–308. [Google Scholar] [CrossRef]
- Capasso, R.; Cristinzio, G.; Evidente, A.; Scognamiglio, F. Isolation, spectroscopy and selective phytotoxic effects of polyphenols from vegetable waste waters. Phytochemistry 1992, 31, 4125–4128. [Google Scholar]
- Spagnolo, A.; Marchi, G.; Peduto, F.; Phillips, A.J.L.; Surico, G. Detection of Botryosphaeriaceae species within grapevine woody tissues by nested PCR, with particular emphasis on the Neofusicoccum parvum/N. ribis complex. Eur. J. Plant Pathol. 2011, 129, 485–500. [Google Scholar] [CrossRef]
- Cristinzio, G. Gravi attacchi di Botryosphaeria obtusa su vite in provincia di Isernia. Inf. Fitopatol. 1978, 6, 21–23. [Google Scholar]
- Lehoczky, J. Black Dead Arm. In Compendium of Grape Diseases; Pearson, R.C., Goheen, A.C., Eds.; The American Phytopathological Society (APS) Press: St. Paul, MN, USA, 1988; p. 35. [Google Scholar]
- Marchi, G.; Peduto, F.; Mugnai, L.; di Marco, S.; Calzarano, F.; Surico, G. Some observations on the relationship of manifest and hidden esca to rainfall. Phytopathol. Mediterr. 2006, 45, 117–126. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Andolfi, A.; Mugnai, L.; Luque, J.; Surico, G.; Cimmino, A.; Evidente, A. Phytotoxins Produced by Fungi Associated with Grapevine Trunk Diseases. Toxins 2011, 3, 1569-1605. https://doi.org/10.3390/toxins3121569
Andolfi A, Mugnai L, Luque J, Surico G, Cimmino A, Evidente A. Phytotoxins Produced by Fungi Associated with Grapevine Trunk Diseases. Toxins. 2011; 3(12):1569-1605. https://doi.org/10.3390/toxins3121569
Chicago/Turabian StyleAndolfi, Anna, Laura Mugnai, Jordi Luque, Giuseppe Surico, Alessio Cimmino, and Antonio Evidente. 2011. "Phytotoxins Produced by Fungi Associated with Grapevine Trunk Diseases" Toxins 3, no. 12: 1569-1605. https://doi.org/10.3390/toxins3121569