Isolation and Biochemical Characterization of Rubelase, a Non-Hemorrhagic Elastase from Crotalus ruber ruber (Red Rattlesnake) Venom

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Biochemical Properties
2.2. Enzyme Activities and Pharmacological Activities
2.3. Determination of Primary Structure and Computer Modeling
2.4. Toxicity Test on Cultured Cells
3. Results and Discussion
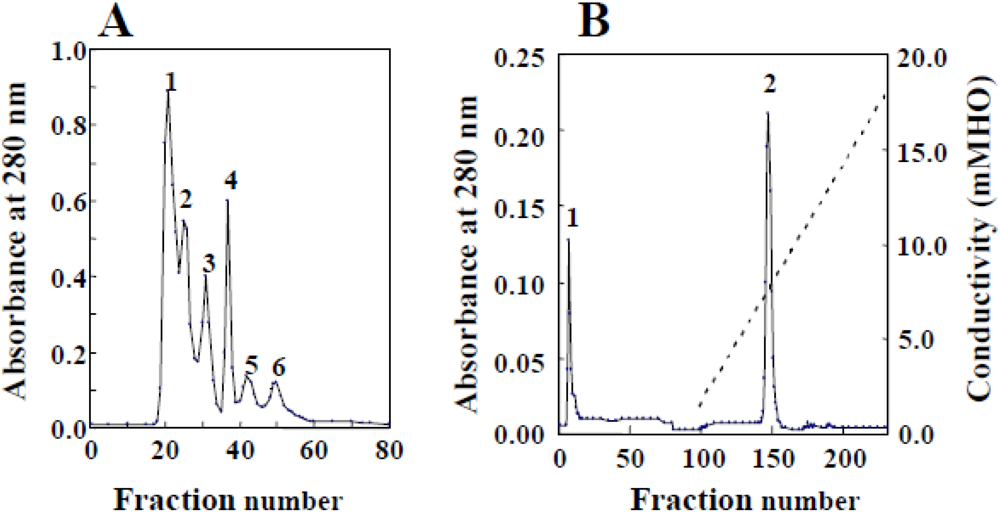
3.1. Isolation and Properties


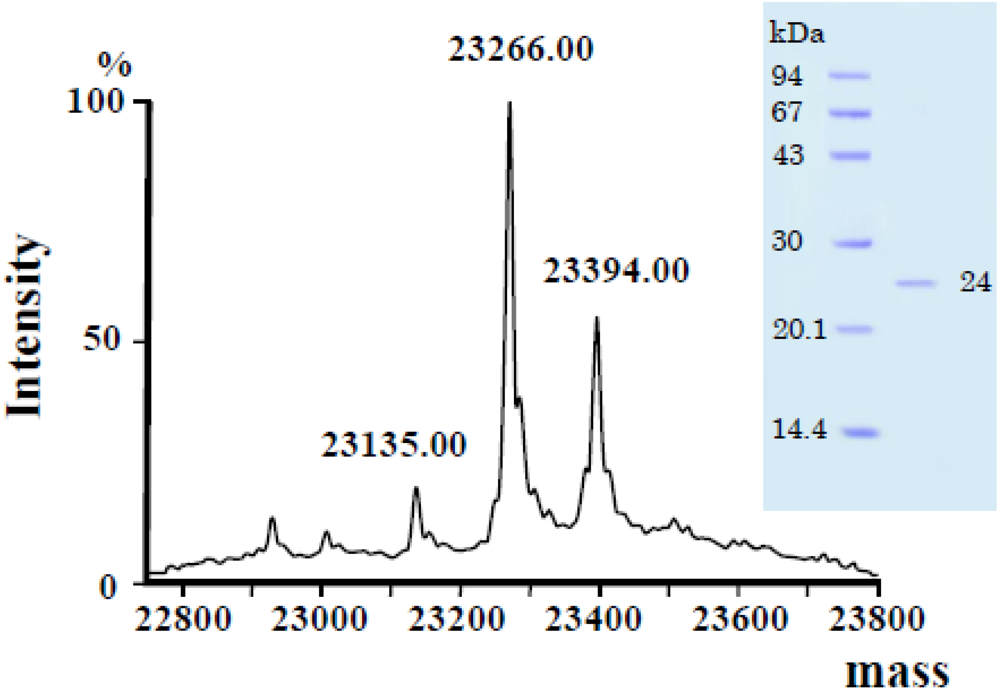
3.2. Primary Structure
3.3. Comparison of Rubelase and Rubelysin (HT-2)
3.4. Comparison of Rubelase with the Low Molecular Weight Metalloproteinases

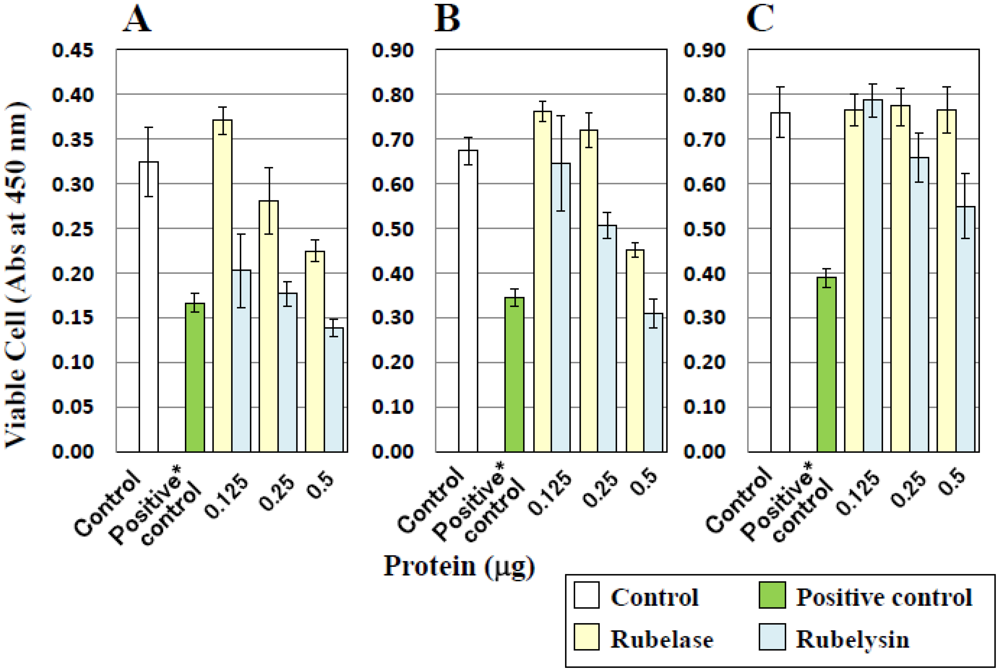
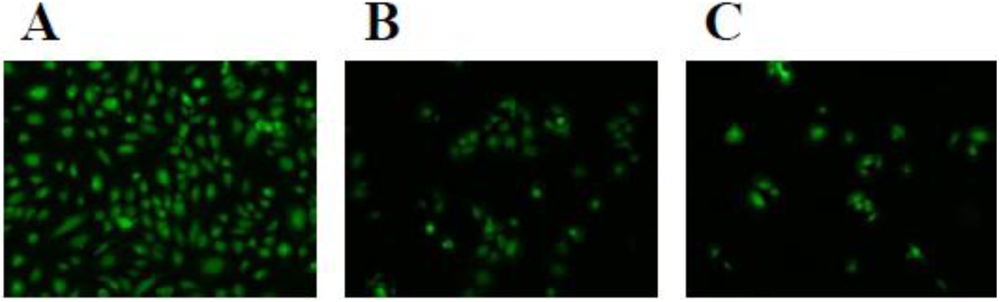
3.5. Toxicity on Cultured Cells


3.6. Molecular Modeling of Rubelase

4. Conclusions
References
- Tu, A.T. Venoms. Chemistry and Molecular Biology; John Wiley & Sons: New York, NY, USA, 1977; pp. 372–399. [Google Scholar]
- Tu, A.T. Rattlesnake Venom: Their Actions and Treatment; Marcel Dekker, Inc.: New York, NY, USA, 1982. [Google Scholar]
- Omori-Satoh, T.; Sadahiro, S. Resolution of the major hemorrhagic component of Trimeresurus flavoviridis venom into two parts. Biochim. Biophys. Acta 1979, 580, 392–404. [Google Scholar] [PubMed]
- Sugihara, H.; Moriura, M.; Nikai, T. Purification and properties of a lethal, hemorrhagic protein “Mucrotoxin A” from the venom of the Chinese habu snake (Trimeresurus mucrosquamatus). Toxicon 1983, 21, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Nikai, T.; Sugihara, H.; Tanaka, T. Enzymochemical studies on snake venoms. II. Purification of lethal protein Ac1-Proteinase in the venom of Agkistrodon acutus. 1977, 97, 507–514. [Google Scholar]
- Bjarnason, J.B.; Tu, A.T. Hemorrhagic toxins from western diamondback rattlesnake (Crotalus atrox) venoms: Isolation and characterization of five toxins and the role of zinc in hemorrhagic toxin e. Biochemistry 1978, 17, 3395–3404. [Google Scholar] [CrossRef] [PubMed]
- Nikai, T.; Mori, N.; Kishida, M.; Sugihara, H.; Tu, A.T. Isolation and biochemical characterization of hemorrhagic toxin f from the venom of Crotalus atrox (western diamondback rattlesnake). Arch. Biochem. Biophys. 1984, 231, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Nikai, T.; Sugihara, H.; Tum, A.T. Biochemical characterization of hemorrhagic toxins with fibrinogenase activity isolated from Crotalus ruber ruber venom. Arch. Biochem. Biophys. 1987, 253, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Imai, K.; Nikai, T.; Sugihara, H.; Ownby, C.L. Hemorrhagic toxin from the venom of Agkistrodon bilineatus (common cantil). Int. J. Biochem. 1989, 21, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Miyata, T.; Takeya, H.; Ozeki, Y.; Arakawa, M.; Tokunaga, F.; Iwanaga, S.; Omori-Satoh, T. Primary structure of hemorrhagic protein, HR2a, isolated from the venom of Trimeresurus flavoviridis. J. Biochem. 1989, 105, 847–853. [Google Scholar] [PubMed]
- Shannon, J.D.; Baramova, E.N.; Bjarnason, J.B.; Fox, J.W. Amino acid sequence of a Crotalus atrox venom metalloproteinase which cleaves type IV collagen and gelatin. J. Biol. Chem. 1989, 264, 11575–11583. [Google Scholar] [PubMed]
- Takeya, H.; Oda, K.; Miyata, T.; Omori-Satoh, T.; Iwanaga, S. The complete amino acid sequence of the high molecular mass hemorrhagic protein HR1B isolated from the venom of Trimeresurus flavoviridis. J. Biol. Chem. 1990, 265, 16068–16073. [Google Scholar] [PubMed]
- Takeya, H.; Onikura, A.; Nikai, T.; Sugihara, H.; Iwanaga, S. Primary structure of a hemorrhagic metalloproteinase, HT-2, isolated from the venom of Crotalus ruber ruber. J. Biochem. 1990, 108, 711–719. [Google Scholar] [PubMed]
- Takeya, H.; Nishida, S.; Nishino, N.; Makinose, Y.; Omori-Sato, T.; Nikai, T.; Sugihara, H.; Iwanaga, S. Primary structures of platelet aggregation inhibitors (disintegrins) autoproteolytically released from snake venom hemorrhagic metalloproteinases and new fluorogenic peptide substrates for these enzymes. J. Biochem. 1993, 113, 473–483. [Google Scholar]
- Hite, L.A.; Jia, L.G.; Bjarnason, J.B.; Fox, J.W. cDNA sequences for four snake venom metalloproteinases: Structure, classification, and their relationship to mammalian reproductive proteins. Arch. Biochem. Biophys. 1994, 308, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Nikai, T.; Kato, C.; Komori, Y.; Nodani, H.; Homma, M.; Sugihara, H. Primary structure of Ac1-Proteinase from the venom of Deinagkistrodon acutus (hundred-pace snake) from Taiwan. Biol. Pharm. Bull. 1995, 18, 631–633. [Google Scholar] [CrossRef] [PubMed]
- Nikai, T.; Taniguchi, K.; Komori, Y.; Masuda, K.; Fox, J.W.; Sugihara, H. Primary structure and functional characterization of bilitoxin-1, a novel dimeric P-II snake venom metalloproteinase from Agkistrodon bilineatus venom. Arch. Biochem. Biophys. 2000, 378, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, L.; Shannon, J.D.; Valente, R.H.; Rucavado, A.; Alape-Giron, A.; Kamiguti, A.S.; Theakston, R.D.; Fox, J.W.; Gutierrez, J.M.; Arni, R.K. Amino acid sequence and crystal structure of BaP1, a metalloproteinase from Bothrops asper snake venom that exerts multiple tissue-damaging activities. Protein Sci. 2003, 12, 2273–2281. [Google Scholar] [PubMed]
- Gong, W.; Zhu, X.; Liu, S.; Teng, M.; Niu, L. Crystal structures of acutolysin A, a three-disulfide hemorrhagic zinc metalloproteinase from the snake venom of Agkistrodon acutus. J. Mol. Biol. 1998, 283, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Gomis-Ruth, F.X.; Kress, L.F.; Kellermann, J.; Mayer, L.; Lee, X.; Huber, R.; Bode, W. Refined 2.0 Å X-ray crystal structure of the snake venom zinc-endopeptidase adamalysin II. Primary and tertiary structure determination, refinement, molecular structure and comparison with astacin, collagenase and thermolysin. J. Mol. Biol. 1994, 239, 513–544. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.F.; Chiou, S.H.; Ko, T.P.; Yuann, J.M.; Wang, A.H. The 1.35 Å structure of cadmium-substituted TM-3, a snake-venom metalloproteinase from Taiwan habu: Elucidation of a TNFa-converting enzyme-like active-site structure with a distorted octahedral geometry of cadmium. Acta Crystallogr. Sect. D: Biol. Crystallogr. 2002, 58, 1118–1128. [Google Scholar] [CrossRef]
- Kumasaka, T.; Yamamoto, M.; Moriyama, H.; Tanaka, N.; Sato, M.; Katsube, Y.; Yamakawa, Y.; Omori-Satoh, T.; Iwanaga, S.; Ueki, T. Crystal structure of H2-proteinase from the venom of Trimeresurus flavoviridis. J. Biochem. 1996, 119, 49–57. [Google Scholar] [PubMed]
- Zhang, D.; Botos, I.; Gomis-Rüth, F.X.; Doll, R.; Blood, C.; Njoroge, F.G.; Fox, J.W.; Bode, W.; Meyer, E.F. Structural interaction of natural and synthetic inhibitors with the venom metalloproteinase, atrolysin C (form d). Proc. Natl. Acad. Sci. USA 1994, 91, 8447–8451. [Google Scholar]
- Fox, J.W.; Serrano, S.M. Structural considerations of the snake venom metalloproteinases, key members of the M12 reprolysin family of metalloproteinases. Toxicon 2005, 45, 969–985. [Google Scholar]
- Fox, J.W.; Serrano, S.M. Insights into and speculations about snake venom metalloproteinase (SVMP) synthesis, folding and disulfide bond formation and their contribution to venom complexity. FEBS J. 2008, 275, 3016–3030. [Google Scholar]
- Mackessy, S.P. Fractionation of red diamond rattlesnake (Crotalus ruber ruber) venom: Protease, phospho- diesterase, L-amino acid oxidase activities and effects of metal ions and inhibitors on protease activity. Toxicon 1985, 23, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, Y.; Nikai, T.; Okumura, Y.; Ogawa, K.; Sugihara, H. Characterization of elastolytic proteinase from Aspergillus flavus: Comparison of the elastolytic proteinase from Aspergillus fumigatus. Jpn. J. Med. Mycol. 1997, 38, 53–59. [Google Scholar] [CrossRef]
- Murata, Y.; Satake, M.; Suzuki, T. Studies on snake venom: XII. Distribution of proteinase activities among Japanese and Formosan snake venoms. J. Biochem. 1963, 53, 431–437. [Google Scholar]
- Ouyang, C.; Teng, C.M. Fibrinogenolytic enzymes of Trimeresurus mucrosquamatus venom. Biochim. Biophys. Acta 1976, 420, 298–308. [Google Scholar] [PubMed]
- Roberts, P.S. Measurement of the rate of plasmin action on synthetic substrates. J. Biol. Chem. 1958, 232, 285–291. [Google Scholar]
- Rinderknecht, H.; Geokas, M.C.; Silverman, P.; Haverback, B.J. A new ultrasensitive method for the determination of proteolytic activity. Clin. Chim. Acta 1968, 21, 197–203. [Google Scholar]
- Charney, J.; Tomarelli, R.M. A colorimetric method for the determination of the proteolytic activity of duodenal juice. J. Biol. Chem. 1947, 171, 501–505. [Google Scholar]
- Moore, G.L. Use of azo-dye-bound collagen to measure reaction velocities of proteolytic enzymes. Anal. Biochem. 1969, 23, 122–127. [Google Scholar]
- Tanzer, M.L.; Gilvarg, C. Creatine and creatine kinase measurement. J. Biol. Chem. 1959, 234, 3201–3204. [Google Scholar]
- Ishiyama, M.; Shiga, M.; Sasamoto, K.; Mizogucgi, M.; He, P. A new sulfonated tetrazolium salt that produces a highly water-soluble formazan dye. Chem. Pharm. Bull. 1993, 41, 1118–1122. [Google Scholar]
- Kurecki, T.; Laskowski, M., Sr.; Kress, L.F. Purification and some properties of two proteinases from Crotalus adamanteus venom that inactivate human a1-proteinase inhibitor. J. Biol. Chem. 1978, 253, 8340–8345. [Google Scholar]
- Nikai, T.; Kato, C.; Komori, Y.; Nodani, H.; Homma, M.; Sugihara, H. Primary structure of Ac1-proteinase from the venom of Deinagkistrodon acutus (hundred-pace snake) from Taiwan. Biol. Pharm. Bull. 1995, 18, 631–633. [Google Scholar] [CrossRef] [PubMed]
- Takeya, H.; Arakawa, M.; Miyata, T.; Iwanaga, S.; Omori-Satoh, T. Primary structure of H2-proteinase, a non-hemorrhagic metalloproteinase, isolated from the venom of the habu snake, Trimeresurus flavoviridis. J. Biochem. 1989, 106, 151–157. [Google Scholar] [PubMed]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Komori, Y.; Sakai, K.; Masuda, K.; Nikai, T. Isolation and Biochemical Characterization of Rubelase, a Non-Hemorrhagic Elastase from Crotalus ruber ruber (Red Rattlesnake) Venom. Toxins 2011, 3, 900-910. https://doi.org/10.3390/toxins3070900
Komori Y, Sakai K, Masuda K, Nikai T. Isolation and Biochemical Characterization of Rubelase, a Non-Hemorrhagic Elastase from Crotalus ruber ruber (Red Rattlesnake) Venom. Toxins. 2011; 3(7):900-910. https://doi.org/10.3390/toxins3070900
Chicago/Turabian StyleKomori, Yumiko, Kaname Sakai, Katsuyoshi Masuda, and Toshiaki Nikai. 2011. "Isolation and Biochemical Characterization of Rubelase, a Non-Hemorrhagic Elastase from Crotalus ruber ruber (Red Rattlesnake) Venom" Toxins 3, no. 7: 900-910. https://doi.org/10.3390/toxins3070900