Overexpression of the Trichoderma brevicompactum tri5 Gene: Effect on the Expression of the Trichodermin Biosynthetic Genes and on Tomato Seedlings

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. Media and Culture Conditions
2.3. DNA and RNA Manipulations
2.4. Gene Expression Studies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Name | Sequence (5´-3´) |
|---|---|---|
| erg1 | erg1-f | CGCTCCGTGCTTCTTCTC |
| erg1 | erg1-r | CTTCTTCTCTCCCGTCTCC |
| tri3 | Tri3-F | CATTCAGCCACCTAACCTAACCG |
| tri3 | Tri3-R | CCATCCTTCAACCACCGTCGGC |
| tri4 | Tri4-F | CTTGATGGAGCCTTCTCAGC |
| tri4 | Tri4-R | CATCAAGATAGTCCTTATGTTC |
| tri6 | Tri6-F | CGTGCTGACGTGGTTCGAGTGC |
| tri6 | Tri6-R | CTATGGAATGGGTCGGCGAATC |
| tri10 | Tri10-F | CGCTCTCATATGAGTACGTTGGC |
| tri10 | Tri10-R | CCATGAATGGTGAAGATGGGC |
| tri11 | Tri11-F | CGCGAGTACGCTTATTACCG |
| tri11 | Tri11-R | GCAGAGCGCACTTCTTCAGTC |
| tri12 | Tri12-F | GTTCCATATCTTCCGCCATATTC |
| tri12 | Tri12-R | GCGATTGACAGAAGCCATTGC |
| tri14 | Tri14-F | GCTGATGCTGAGCTTGCAAGTG |
| tri14 | Tri14-R | GCCAAGAGGCTCTTGGACGAAG |
| β-Tubulin | T-tub-F | GAATATCAACAATACCAGGATGG |
| β-Tubulin | T-tub-R | AGGATTGGTATTGATCATCAGCA |
2.5. Tomato Seedling Assay
3. Results
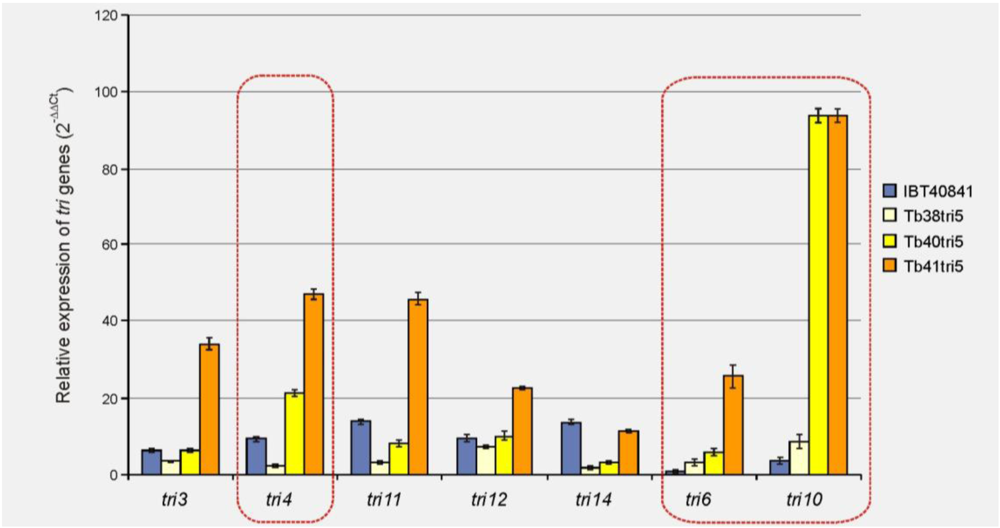
3.1. Effect of tri5 Gene Overexpression on the Expression of Trichodermin Biosynthetic Genes

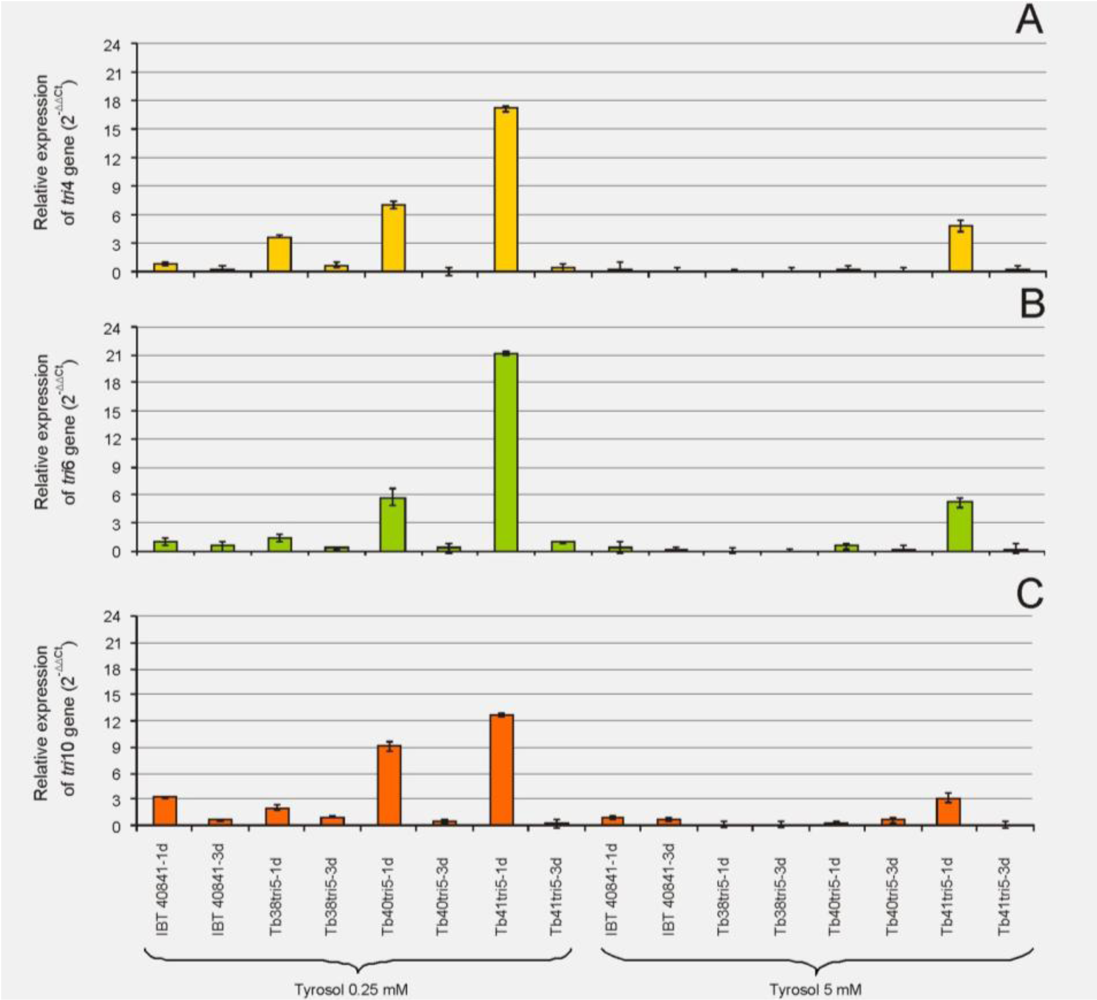
3.2. Effect of Tyrosol on the Expression of the tri4, tri6 and tri10 Genes in the tri5 Overexpressing Transformants

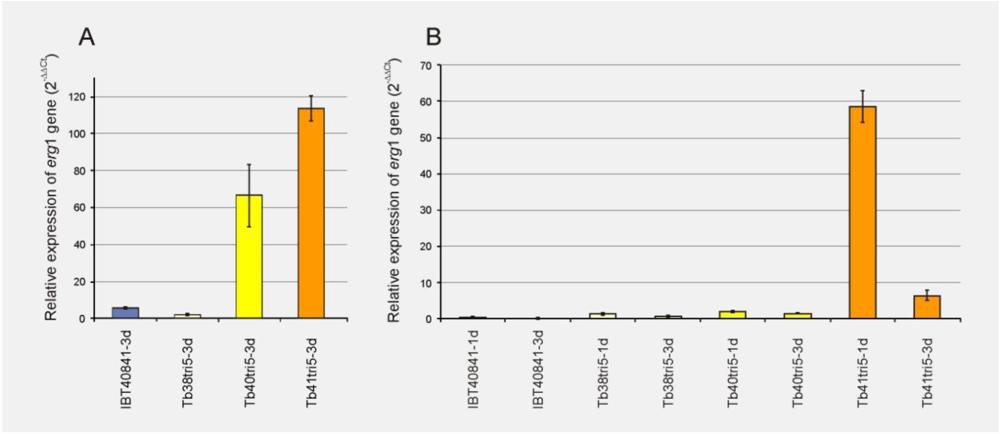
3.3. Effect of the Overexpression of the Tbtri5 Gene in the Expression of erg1 Gene

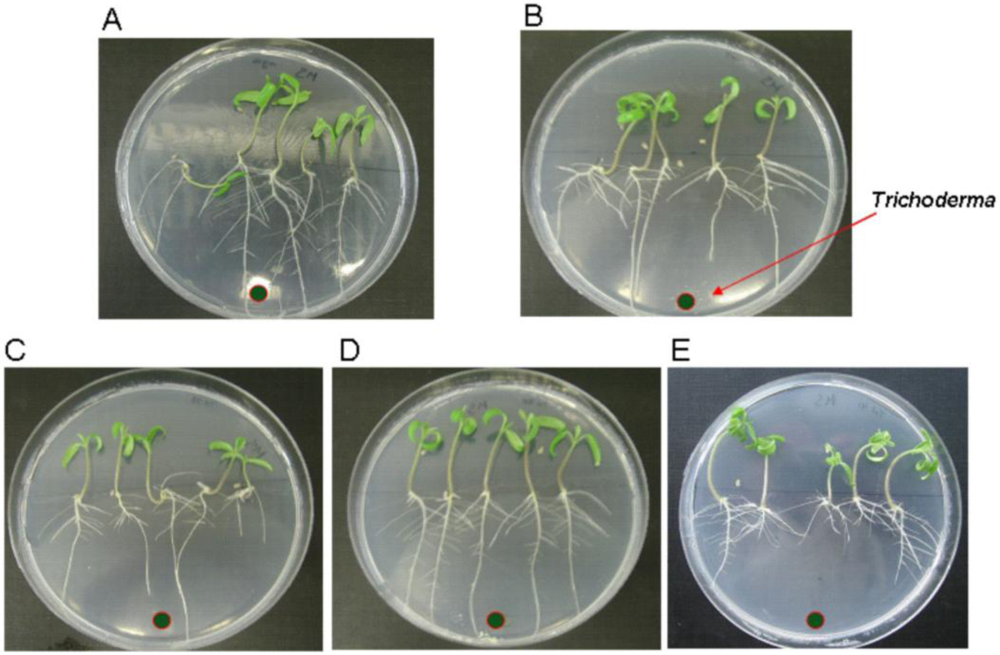
3.4. Effect of the tri5 Overexpression on Tomato Seedlings

4. Discussion

5. Conclusions
Acknowledgments
References
- Monte, E. Understanding Trichoderma: Between biotechnology and microbial ecology. Int. Microbiol. 2001, 4, 1–4. [Google Scholar] [PubMed]
- Weindling, R. Trichoderma lignorum as a parasite of other soil fungi. Phytopathology 1932, 22, 837–845. [Google Scholar]
- Kubicek, C.P.; Penttilä, M. Regulation of Production of Plant Polysacchride Degrading Enzymes by Trichoderma. In Trichoderma and Gliocladium: Enzymes, Biological Control and Commercial Applications; Kubicek, C.P., Harman, G.E., Eds.; Taylor & Francis: London, UK, 1998; Volume 2, pp. 49–71. [Google Scholar]
- Hjeljord, L.; Tronsmo, A. Trichoderma and Gliocladium in Biological Control: An Overview. In Trichoderma and Gliocladium: Enzymes, Biological Control and Commercial Applications; Kubicek, C.P., Harman, G.E., Eds.; Taylor and Francis: London, UK, 1998; Volume 2, pp. 131–153. [Google Scholar]
- Vizcaíno, J.A.; Sanz, L.; Basilio, A.; Vicente, F.; Gutiérrez, S.; Hermosa, M.R.; Monte, E. Screening of antimicrobial activities in Trichoderma isolates representing three Trichoderma sections. Mycol. Res. 2005, 109, 1397–1406. [Google Scholar] [PubMed]
- Goldstein, J.L.; Brown, M.S. Regulation of the mevalonate pathway. Nature 1990, 343, 425–430. [Google Scholar]
- Chang, M.C.; Keasling, J.D. Production of isoprenoid pharmaceuthicals by engineering isoprenoids. Nat. Chem. Biol. 2006, 2, 674–681. [Google Scholar]
- Kirby, J.; Keasling, J.D. Biosynthesis of plant isoprenoids: Perspectives for microbial engineering. Annu. Rev. Plant Biol. 2009, 60, 335–355. [Google Scholar]
- Beremand, M.N.; McCormick, S.P. Biosynthesis and Regulation of Trichothecene Production by Fusarium Species. In Handbook of Applied Mycology. Mycotoxins in Ecological Systems; Bhatnagar, D., Lilehoj, E.B., Arora, D.K., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1992; Volume 5, pp. 360–389. [Google Scholar]
- Kimura, M.; Tokai, T.; Takahashi-Ando, N.; Ohsato, S.; Fujita, M. Molecular and genetic studies of Fusarium trichothecene biosynthesis: Pathways, genes, and evolution. Biosci. Biotechnol. Biochem. 2007, 71, 2105–2123. [Google Scholar]
- Cardoza, R.E.; Malmierca, M.G.; Hermosa, M.R.; Alexander, N.J.; McCormick, S.P.; Proctor, R.H.; Tijerino, A.M.; Rumbero, A.; Monte, E.; Gutiérrez, S. Identification of loci and functional characterization of trichothecene biosynthetic genes in the filamentous fungus Trichoderma. Appl. Environ. Microbiol. 2011, 77, 4867–4877. [Google Scholar] [PubMed]
- Degenkolb, T.; Dieckmann, R.; Nielsen, K.F.; Gräfenhan, T.; Theis, Ch.; Zafari, D.; Chaverri, P.; Ismaiel, A.; Brückner, H.; Von Döhren, H.; et al. The Trichoderma brevicompactum clade: A separate lineage with new species, new peptaibiotics, and mycotoxins. Mycol. Prog. 2008, 7, 177–219. [Google Scholar] [CrossRef]
- Tijerino, A.; Cardoza, R.E.; Moraga, J.; Malmierca, M.G.; Vicente, F.; Aleu, J.; Collado, I.G.; Gutiérrez, S.; Monte, E.; Hermosa, R. Overexpression of the trichodiene synthase gene tri5 increases trichodermin production and antimicrobial activity in Trichoderma brevicompactum. Fungal Genet. Biol. 2011, 48, 285–296. [Google Scholar] [PubMed]
- Brown, D.W.; Dyer, R.B.; McCormick, S.P.; Kendra, D.F.; Plattner, R.D. Functional demarcation of the Fusarium core trichothecene gene cluster. Fungal Genet. Biol. 2004, 41, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Trapp, S.C.; Hohn, T.M.; McCormick, S.; Jarvis, B.B. Characterization of the gene cluster for biosynthesis of macrocyclic trichothecenes in Myrothecium roridum. Mol. Gen. Genet. 1998, 257, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Tarus, P.K.; Langát-Thoruwa, C.C.; Wanyonyi, A.W.; Chhlabra, S.C. Bioactive metabolites from Trichoderma harzianum and Trichoderma longibrachiatum. Bull. Chem. Soc. Ethiop. 2003, 17, 185–190. [Google Scholar]
- Alem, M.A.; Oteef, M.D.; Flowers, T.H.; Douglas, L.J. Production of tyrosol by Candidda albicans biofilms and its role in quorum sensing and biofilm development. Eukaryot. Cell 2006, 5, 1770–1779. [Google Scholar] [CrossRef] [PubMed]
- Daoubi, M.; Pinedo-Rivilla, C.; Rubio, M.B.; Hermosa, R.; Monte, E.; Aleu, J.; Collado, I.G. Hemisynthesis and absolute configuration of novel 6-pentyl-2H-pyran-2-one derivatives from Trichoderma spp. Tetrahedron 2009, 65, 4834–4840. [Google Scholar]
- Sentheshanmuganathan, E.; Elsden, S.R. The mechanism of the formation of tyrosol by Saccharomyces cerevisiae. Biochem. J. 1958, 69, 210–218. [Google Scholar] [PubMed]
- Covas, M.I.; Miró-Casas, E.; Fitó, M.; Farré-Albadalejo, M.; Gimeno, E.; Marrugat, J.; de la Torre, R. Bioavailability of tyrosol, an antioxidant phenolic compound present in wine and olive oil, in humans. Drug. Exp. Clin. Res. 2003, 29, 203–206. [Google Scholar]
- Ahn, E.Y.; Jiang, Y.; Zhang, Y.; Son, E.M.; You, S.; Kang, S.W.; Pardk, J.; Jung, J.H.; Lee, B.J.; Kim, D. Citotoxicity of p-tyrosol and its derivatives may correlate with the inhibition of DNA replication initiation. Oncol. Rep. 2008, 19, 527–534. [Google Scholar] [PubMed]
- Cardoza, R.E.; Vizcaíno, J.A.; Hermosa, M.R.; Sousa, S.; González, F.J.; Llobell, A.; Monte, E.; Gutiérrez, S. Cloning and characterization of the erg1 gene of Trichoderma harzianum: Effect of the erg1 silencing on ergosterol biosynthesis and resistance to terbinafine. Fungal Genet. Biol. 2006, 43, 164–178. [Google Scholar] [CrossRef] [PubMed]
- Vizcaíno, J.A.; Cardoza, R.E.; Dubost, L.; Bodo, B.; Gutiérrez, S.; Monte, E. Detection of peptaibols and partial cloning of a putative peptaibol synthetase gene from T. harzianum CECT 2413. Folia Microbiol. 2006, 51, 114–120. [Google Scholar] [CrossRef]
- Penttilä, M.; Nevalainen, H.; Ratto, M.; Salminen, E.; Knowles, J. A versatile transformation system for the cellulolytic filamentous fungus Trichoderma reesei. Gene 1987, 61, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russel, D.W. Molecular Cloning; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta CT). Methods 2001, 25, 402–408. [Google Scholar]
- Desjardins, A.E.; Hohn, T.M.; McCormick, S.P. Trichothecene biosynthesis in Fusarium species: Chemistry, genetics, and significance. Microbiol. Rev. 1993, 57, 595–604. [Google Scholar] [PubMed]
- De Sordi, L.; Mühlschlegel, F.A. Quorum sensing and fungal-bacterial interactions in Candida albicans: A communicative network regulating microbial coexistence and virulence. FEMS Yeast Res. 2009, 9, 990–999. [Google Scholar] [CrossRef] [PubMed]
- Tokai, T.; Koshino, H.; Takahashi-Ando, N.; Sato, M.; Fujimura, M.; Kimura, M. Fusarium tri4 encodes a key multifunctional cytochrome P450 monooxygenase for four consecutive oxygenation steps in trichothecene biosynthesis. Biochem. Biophs. Res. Commun. 2007, 353, 412–417. [Google Scholar] [CrossRef]
- Kruppa, M. Quorum sensing and Candida albicans. Mycoses 2009, 51, 1–10. [Google Scholar] [CrossRef]
- Nickerson, K.W.; Atkin, A.L.; Hornby, J.M. Quorum sensing in Dimorphic fungi: farnesol and beyond. Appl. Environ. Microbiol. 2006, 72, 3805–3813. [Google Scholar]
- Hornby, J.M.; Kebaara, B.W.; Nickerson, K.W. Farnesol biosynthesis in Candida albicans: Cellular response to sterol inhibition by zaragozic acid. B. Antimicrob. Agents Chemother. 2003, 47, 2366–2369. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 84, 377–393. [Google Scholar]
- Harman, G.E. Overview of mechanisms and uses of Trichoderma spp. Phytopathology 2006, 96, 190–194. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma-plant-pathogen interactions in soil agro-ecoystems. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol. Plant Microbe Interact. 1995, 8, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Cutler, H.G.; LeFiles, J. Trichodermin: Effects on plants. Plant Cell Physiol. 1978, 19, 177–182. [Google Scholar]
- Nielsen, K.F.; Gräfenhan, T.; Zafari, D.; Thrane, U. Trichothecene production by Trichoderma brevicompactum. J. Agric. Food Chem. 2005, 53, 8190–8196. [Google Scholar] [PubMed]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tijerino, A.; Hermosa, R.; Cardoza, R.E.; Moraga, J.; Malmierca, M.G.; Aleu, J.; Collado, I.G.; Monte, E.; Gutierrez, S. Overexpression of the Trichoderma brevicompactum tri5 Gene: Effect on the Expression of the Trichodermin Biosynthetic Genes and on Tomato Seedlings. Toxins 2011, 3, 1220-1232. https://doi.org/10.3390/toxins3091220
Tijerino A, Hermosa R, Cardoza RE, Moraga J, Malmierca MG, Aleu J, Collado IG, Monte E, Gutierrez S. Overexpression of the Trichoderma brevicompactum tri5 Gene: Effect on the Expression of the Trichodermin Biosynthetic Genes and on Tomato Seedlings. Toxins. 2011; 3(9):1220-1232. https://doi.org/10.3390/toxins3091220
Chicago/Turabian StyleTijerino, Anamariela, Rosa Hermosa, Rosa E. Cardoza, Javier Moraga, Monica G. Malmierca, Josefina Aleu, Isidro G. Collado, Enrique Monte, and Santiago Gutierrez. 2011. "Overexpression of the Trichoderma brevicompactum tri5 Gene: Effect on the Expression of the Trichodermin Biosynthetic Genes and on Tomato Seedlings" Toxins 3, no. 9: 1220-1232. https://doi.org/10.3390/toxins3091220