Monoclonal Antibodies and Toxins—A Perspective on Function and Isotype

1
Department of Microbiology and Immunology, Albert Einstein College of Medicine, 1300 Morris Park Ave, Bronx, NY 10461, USA
2
Division of Infectious Diseases of the Department of Medicine, Albert Einstein College of Medicine, 1300 Morris Park Ave, Bronx, NY 10461, USA
*
Author to whom correspondence should be addressed.
Toxins 2012, 4(6), 430-454; https://doi.org/10.3390/toxins4060430
Submission received: 29 April 2012
/
Revised: 6 June 2012
/
Accepted: 7 June 2012
/
Published: 11 June 2012
(This article belongs to the Special Issue Toxin-Antibody Interactions)
Abstract
:Antibody therapy remains the only effective treatment for toxin-mediated diseases. The development of hybridoma technology has allowed the isolation of monoclonal antibodies (mAbs) with high specificity and defined properties, and numerous mAbs have been purified and characterized for their protective efficacy against different toxins. This review summarizes the mAb studies for 6 toxins—Shiga toxin, pertussis toxin, anthrax toxin, ricin toxin, botulinum toxin, and Staphylococcal enterotoxin B (SEB)—and analyzes the prevalence of mAb functions and their isotypes. Here we show that most toxin-binding mAbs resulted from immunization are non-protective and that mAbs with potential therapeutic use are preferably characterized. Various common practices and caveats of protection studies are discussed, with the goal of providing insights for the design of future research on antibody-toxin interactions.
Keywords:
antibody; neutralization; protection; clearance; vaccine; therapeutics; isotype; animal model; in vivo; disease enhancement1. Introduction
Humoral immunity against bacterial toxins has been studied for over a century. This research journey started when Behring and Kitasato established the existence of humoral immunity by showing that passive transfer of antibodies from the blood of immunized animals could protect non-immune animals against diphtheria [1]. Serum therapy was commonly used to treat infectious diseases until the discovery of sulfonamide in the 1930s [2]. Today serum therapy remains the only prophylactic and therapeutic option against many toxin-mediated and viral diseases. Toxins are excellent target antigens for antibodies, as they are usually structurally distinct from the self-antigens expressed by the host cells. In contrast, anti-infective drugs are usually not selective enough to differentiate pathogenic strains from non-pathogenic flora and thereby have the potential to cause a long-term disruption on the flora dynamics [3]. To date specific antibody is the only compound that can neutralize toxin and they are attractive for therapeutic development because antimicrobial drugs can kill the microbes but does not eradicate pre-formed toxins.
The development of hybridoma technology in 1975 allowed, for the first time, the functional and structural analysis of individual immunoglobulin molecules [4]. Individual antibody molecules can now be isolated in monoclonal preparations and produced in an unlimited supply. The availability of monoclonal antibodies (mAbs) with defined specificity and expressing a single isotype significantly reduces its toxicity relative to immune serum [5], despite the fact that non-human mAbs used directly as therapeutics against human diseases are still immunogenic [6]. The high purity of the preparation also enhances the biological activity per mass of protein [7], since all immunoglobulins are specific for the target toxin. This is particularly important for the development of prophylactic and therapeutic antibodies against infectious and toxin-mediated diseases [8]. In addition to the neutralizing activity that is mediated by the binding of the variable region to toxin, the Fc region of mAb mediates various effector functions including antibody-dependent cell-mediated cytotoxicity (ADCC), complement activation, and opsonization [9]. The combined potential of an immunoglobulin molecule for microbe eradication, toxin neutralization, and the enhancement of host immune system make mAbs attractive candidates for the next generation of magic bullet against infectious diseases.
The toxin neutralizing efficacy of an antibody molecule is usually defined by the activity of its monoclonal preparation. In general, a mAb can be classified as protective, indifferent, or disease-enhancing depending on how it modifies the course of infection or toxemia measured by various types of protection assays. Protective mAb benefits the host primarily by neutralizing the toxin, while disease-enhancing mAb intensifies its toxicity. Indifferent mAb binds to the toxin with no apparent effect on toxicity and host damage. This paper reviews the literature on mAbs to six toxins listed on Pubmed and analyzes the prevalence of the various functional types and isotypes. All the mAbs described result from immunization with whole toxin, toxin component/subunit, or clinical vaccine. Uncharacterized mAbs were not included in our analysis. Only those studies that described the first appearance of a particular mAb are listed—the follow up studies about the chimerization or humanization of a previously reported mAb will not be discussed in this review. In addition, we acknowledge that a significant number of toxin-binding fragment antigen binding (Fab) and single-chain variable fragment (scFv) have been generated as alternative forms of antibody-derived reagents that possess great potential in prophylaxis and therapeutics [10,11,12], but this paper will focus only on mAbs that originate from hybridoma cells.
2. Selective Characterization of mAbs
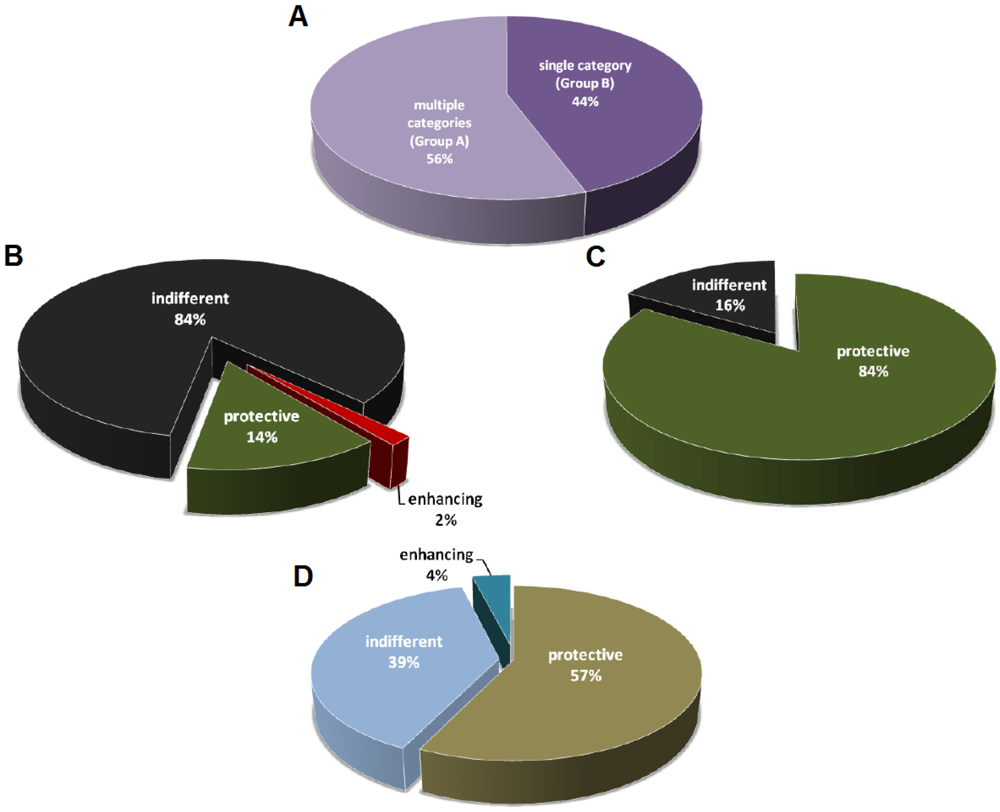
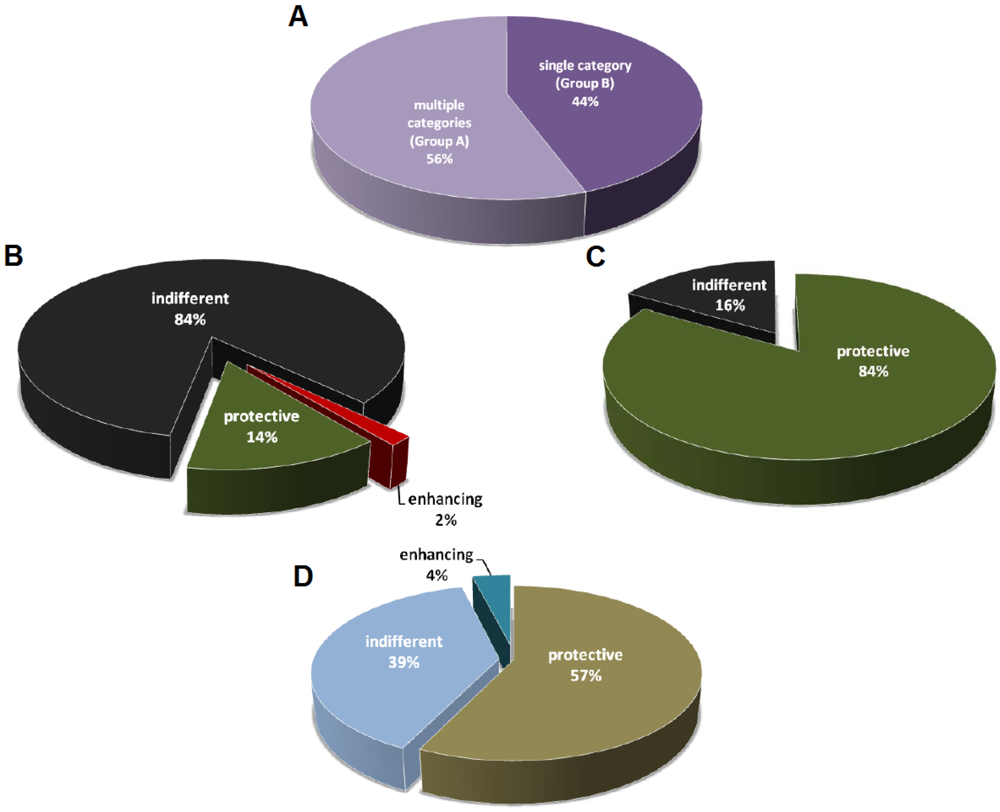
The mAb studies listed in Table 1 are divided into two groups, with the first one (Group A) describing two or more categories of mAbs (among protective, indifferent, and disease-enhancing mAbs), and the second (Group B) describing a single category of mAbs (Figure 1A). In Group A 14% of the mAbs are protective, 2% are disease-enhancing, and the remainders are indifferent with no measurable effect on the host (Figure 1B). In contrast, for the studies that described only one category of mAbs, 84% of which are protective against toxins (Figure 1C). Humoral responses produce complex polyclonal mixtures with different specificities, isotypes, and affinities, and it is possible that the antibodies reported in Group A are more representative of natural antibody responses from immunization. The fact that a high percentage of the mAbs described in Group B is protective suggests that these studies are skewed to report primarily mAbs with prophylactic or therapeutic potential. A positive result publication bias could have led to significantly fewer publications for those studies that generated non-productive or indifferent mAbs. Consequently, the indifferent mAbs with no apparent benefits to the host are usually suggested as potential diagnostic or research tools. Not surprisingly, given the lack of therapeutic potential, disease-enhancing mAbs are never described on their own in any toxin-mAb studies. Disease-enhancing mAbs have been historically labeled as deleterious by-products of immunizations, while some of them are shown to enhance toxicity by increasing the activation rate of toxin components and through the interaction with the Fc region [13,14,15,16,17,18,19,20,21].
The field of antibody studies revolves around the discovery of novel and useful immunoglobulins against diseases, so it is not surprising that most effort is selectively given to characterize and report those mAbs with therapeutic potential (Figure 1D). While more than 20 mAbs have been developed and marketed as therapeutics against multiple types of cancers and inflammatory diseases, to date only one mAb is licensed for an infectious disease [22]. Palivizumab was approved by FDA in 1998 for the prevention of respiratory syncytial virus (RSV) infection in high-risk pediatric patients [23,24]. Despite of the very low approval rate of mAb against infectious diseases, research institutes and companies still devote large amount of time and resources in that area given that they remain the most reliable means to neutralize toxins. However, the effort in mAb discovery and development has always been spent on the candidates with the greatest potential given limited resources and keen competition [25], and this is evident in the fact that the majority of studies in Group B is focused on protective antibodies (Figure 1C).

{kind=link}
{kind=link}
{kind=link}
| Toxin | Authors | Reference | Binding sites/ domains/subunits | Protective | Indifferent | Enhancing | Uncharacterized | In vivo model |
|---|---|---|---|---|---|---|---|---|
| Shiga Toxin / Shiga-like Toxin | Griffin et al. (1983) | [26] | Stx | 25 (IgG1) | 63 | - | - | - |
| Donohue-Rolfe et al. (1984) | [27] | Stx | 3 (IgG1) | - | - | - | - | |
| Strockbine et al. (1985) | [28] | SLT | 3 (IgG1) | - | - | - | mouse (CD-1) | |
| Downes et al. (1988) | [29] | SLT-II | 5 (IgG1) | - | - | - | - | |
| Perera et al. (1988) | [30] | SLT-II | 5 (80% IgM and 20% IgG1) | - | - | - | - | |
| Donohue-Rolfe et al. (1989) | [31] | Stx, SLT-II | 3 (66.6% IgG1 and 33.3% IgG2b) | 2 (IgM) | - | - | - | |
| Padhye et al. (1989) | [32] | SLT-I, SLT-II | 3 (66.6% IgG1 and 33.3% IgG2b) | 11 (90.9% IgM and 9.1% IgG2b) | - | - | mouse | |
| Islam and Stimson (1990) | [33] | Stx | 4 (IgG1) | - | - | - | mouse | |
| Qadri et al. (1993) | [34] | Stx | - | 1 (IgM) | - | - | - | |
| Nakao et al. (1999) | [35] | SLT-II | 1 (IgG1) | - | - | - | - | |
| Mukherjee et al. (2002) | [36] | SLT-I | 5 (40% IgM and 60% IgG1) | 5 (IgM) | - | - | mouse (Swiss Webster) | |
| Mukherjee et al. (2002) | [37] | SLT-II | 8 (IgG1) | 28 (96.4% IgG1 and 3.6% IgG3) | - | - | mouse (Swiss Webster) and gnotobiotic piglet | |
| Nakao et al. (2002) | [38] | SLT-I | 1 | - | - | - | - | |
| Tanikawa et al. (2008) | [39] | SLT-I | 1 (IgG1) | 1 (IgA) | - | - | - | |
| Pertussis Toxin | Sato et al. (1984) | [40] | S1, S4 | 1 (IgG2a) | 2 (IgG1) | - | - | mouse (Slc:ddY) |
| Frank and Parker (1984) | [41] | S2 | - | 2 (IgG1) | - | - | mouse (CFW) | |
| Sato et al. (1987) | [42] | S2, S3 | 2 (IgG1) | - | - | - | - | |
| Kenimer et al. (1989) | [43] | S1, S4 | 3 (IgG1) | 3 (IgG1) | - | - | - | |
| Lang et al. (1989) | [44] | S2, S3, S4 | 5 (40% IgG1 and 60% IgG2a) | 8 (25% IgG1, 62.5% IgG2a and 12.5% IgG2b) | - | - | - | |
| Sato and Sato (1990) | [45] | S1, S2, S3, S2–3, S4 | 5 (IgG1) | 15 (IgG1) | - | - | mouse | |
| Halperin et al. (1991) | [46] | S1, S3 | 3 | - | - | - | mouse (CFW) | |
| Kenimer et al. (1991) | [47] | S1 | - | 2 (IgG1 and IgG3) | - | - | - | |
| Sato et al. (1991) | [48] | S2, S3, S2–3, S4, S5 | 2 | 10 | - | - | mouse | |
| Walker et al. (1991) | [49] | S1, S2–3, S4 | 3 (IgG1) | - | - | - | - | |
| Zaccolo et al. (1992) | [50] | S3 | 1 (IgG1) | - | - | - | mouse (BALB/c) | |
| Lee et al. (1999) | [51] | Adenylate cyclase toxin | 4 (75% IgG1 and 25% IgG2a) | 8 (62.5% IgG1, 25% IgG2a, and 12.5% IgG2b) | - | - | - | |
| Pootong et al. (2007) | [52] | S1 | 1 (IgG1) | - | - | 6 | - | |
| Anthrax Toxin | Little et al. (1988) | [53] | PA | 2 (IgG1) | 35 (17.1% IgM, 62.9% IgG1, 11.4% IgG2a, 8.6% IgG2b) | - | - | Fisher 344 rat |
| Little et al. (1990) | [54] | LF | 4 (IgG1) | 59 (30.5% IgM, 40.7% IgG1, 22% IgG2a, 3.4% IgG3, 3.4% IgA) | - | - | Fisher 344 rat | |
| Little et al. (1994) | [55] | EF | 1 (IgG1) | 9 (88.9% IgG1, 11.1% IgG2a) | - | - | - | |
| Little et al. (1996) | [56] | PA63 | 2 (IgG1 and IgG2b) | - | - | - | Fisher 344 rat | |
| Zhao et al. (2003) | [57] | LF | 1 | - | - | - | mouse (nude) | |
| Belova et al. (2004) | [17] | PA | 2 | - | 1 | - | - | |
| Brossier et al. (2004) | [58] | PA | 9 | 87 | - | - | mouse (OF1) | |
| Kozel et al. (2004) | [59] | poly γ-D-glutamic acid | 1 (IgG1) | - | - | 4 (IgG3) | mouse (BALB/c) | |
| Mohamed et al. (2004) | [18] | PA | - | 22 | 21 (IgG2a) | - | - | |
| Sawada-Hirai et al. (2004) | [60] | PA | 3 (IgG1) | 1 (IgG1) | - | - | Fisher 344 rat | |
| Lim et al. (2005) | [61] | LF | 2 (IgG1) | - | - | - | Fisher 344 rat | |
| Chen et al. (2006) | [62] | PA, LF | 2 | 4 | - | - | Fisher 344 rat | |
| Gubbins et al. (2006) | [63] | PA | 3 (IgG1) | 8 (IgG1) | - | - | - | |
| Rivera et al. (2006) | [64] | PA | 2 (IgG1 and IgG2b) | - | - | - | mouse (BALB/c) | |
| Vitale et al. (2006) | [65] | PA | 1 (IgG1) | - | - | - | rabbit | |
| Albrecht et al. (2007) | [66] | PA, LF | 2 (IgG1) | - | - | - | mouse (A/J) | |
| Kozel et al. (2007) | [67] | poly γ-D-glutamic acid | 5 (IgG3) | 1 (IgG1) | - | - | mouse (BALB/c) | |
| Staats et al. (2007) | [68] | PA, LF | 2 | 2 | - | - | mouse (BALB/c) | |
| Abboud et al. (2009) | [69] | PA | 1 (IgG1) | 3 (33.3% IgM, 66.7% IgG1) | - | - | mouse (BALB/c) | |
| Chen et al. (2009) | [70] | LF | 2 (IgG1) | 1 | - | 89 | Fisher 344 rat | |
| Kelly-Cirino and Mantis (2009) | [71] | PA | 1 (IgG1) | 2 (IgG1 and IgG2a) | - | 2 | mouse (BALB/c) | |
| Rosenfeld et al. (2009) | [72] | PA | 101 | 499 | - | - | Fisher 344 rat, Hartley guinea pig | |
| Winterroth et al. (2010) | [73] | EF | 1 (IgM) | 5 (20% IgM, 80% IgG1) | - | - | mouse (A/JCr) | |
| Chen et al. (2011) | [74] | poly γ-D-glutamic acid | 2 (IgG1 and IgG3) | - | - | 3 | mouse (BALB/c) | |
| Kulshreshtha and Bhatnagar (2011) | [75] | LF and EF | 1 (IgG2b) | - | - | - | mouse (BALB/c) | |
| Leysath et al. (2011) | [76] | EF | 3 (IgG1) | 1 (IgG1) | - | 78 | mouse (BALB/cJ, C57BL/6J) | |
| Little et al. (2011) | [19] | PA | - | 56 | 17 | - | Fisher 344 rat | |
| vor dem Esche et al. (2011) | [77] | LF | 1 (IgG1) | 17 | - | - | mouse (A/J) | |
| Chow et al. (manuscript in prep.) | - | PA | 2 (IgG2a) | 16 (IgG1) | 6 (83.3% IgG1, 16.7% IgG2a) | - | mouse (BALB/c) | |
| Ricin Toxin | Colombatti et al. (1986) | [20] | RT, RTA | 3 | 4 | 1 | 13 | - |
| Colombatti et al. (1987) | [78] | RTB | 1 (IgG2a) | - | - | - | - | |
| Chanh et al. (1993) | [79] | RT | 1 (IgG1) | 19 | - | - | mouse (BALB/c) | |
| Lemley et al. (1994) | [80] | RTA | 2 (IgG1) | - | - | - | mouse (BALB/c) | |
| Maddaloni et al. (2004) | [21] | RT, RTA, RTB | 18 | 19 | 1 (IgG1) | - | mouse (CD-1) | |
| Dertzbaugh et al. (2005) | [81] | RT, RTA, RTB | 6 (IgG1) | 23 | - | - | - | |
| Mantis et al. (2006) | [82] | RTA, RTB | 4 (IgA) | - | - | 20 (IgA) | - | |
| McGuinness and Mantis (2006) | [83] | RTB | 1 (IgG1) | - | - | - | - | |
| Pelat et al. (2009) | [84] | RTA | 1 | 18 | - | - | - | |
| Neal et al. (2010) | [85] | RTA | 1 (IgG1) | - | - | - | mouse (BALB/c) | |
| O'Hara et al. (2010) | [86] | RTA | 24 | 394 | - | - | mouse (BALB/c) | |
| Dai et al. (2011) | [87] | RTA | 3 (IgG1) | 14 | - | - | mouse | |
| Prigent et al. (2011) | [88] | RTA, RTB | 7 | 24 | - | - | - | |
| Yermakova and Mantis (2011) | [89] | RTB | 2 | ~100 | - | - | mouse (BALB/c) | |
| Botulinum Toxin | Oguma et al. (1982) | [90] | Type C1 | 2 (IgG1) | 2 (IgG1) | - | - | mouse (ddY) |
| Oguma et al. (1984) | [91] | Type C1, D | 17 | 11 | - | - | - | |
| Kozaki et al. (1986) | [92] | Type E | 3 (IgG1) | 3 (66.7% IgG1, 33.3% IgG2b) | - | - | - | |
| Ferreira et al. (1987) | [93] | Type A | - | 1 (IgG1) | - | 60 | mouse (Swiss Webster) | |
| Simpson et al. (1990) | [94] | Type E | 3 | 1 | - | - | mouse | |
| Toratani et al. (1993) | [95] | ADP-ribosyltransferase C3 | 4 (IgG2b and IgG3) | - | - | - | - | |
| Cenci Di Bello et al. (1994) | [96] | Type A | - | 7 (85.7% IgG1, 14.3% IgG2b) | - | - | mouse | |
| Noah et al. (1995) | [97] | Type B | - | 4 | - | - | mouse | |
| Amersdorfer et al. (1997) | [98] | Type A | 2 | 3 | - | - | - | |
| Brown et al. (1997) | [99] | Type F | 3 | 23 | - | - | mouse | |
| Pless et al. (2001) | [100] | Type A | 33 | 455 | - | - | mouse | |
| Wu et al. (2001) | [101] | Type A | 2 (IgG1) | 14 | - | - | mouse (ICR) | |
| Kamata et al. (2002) | [102] | ADP-ribosyltransferase C3 | 1 (IgG1) | - | - | - | - | |
| Yang et al. (2004) | [103] | Type B | 1 (IgG1) | - | - | - | mouse (ICR) | |
| Adekar et al. (2008) | [104] | Type A | 1 (IgG1) | 19 | - | - | mouse (Swiss Webster) | |
| Adekar et al. (2008) | [105] | Type A | 1 (IgG1) | - | - | - | mouse | |
| Adekar et al. (2008) | [106] | Type A | 1 (IgM) | 1 (IgM) | - | - | mouse (Swiss Webster) | |
| Zhou et al. (2009) | [107] | Type B | 1 | - | - | - | - | |
| Mazuet et al. (2010) | [108] | Type A | 12 (66.7% IgG1, 33.3% IgG2a) | 2 (IgG1) | - | - | mouse (Swiss Webster) | |
| Corbett et al. (2011) | [109] | Type A | 1 (IgG1) | - | - | 7 | mouse (Swiss Webster) | |
| Montgomery et al. (2011) | [110] | Type C | - | 1 | - | - | mouse (CD-1) | |
| Enterotoxin B (SEB) | Lin et al. (1988) | [111] | SEB | 4 (25% IgM, 75% IgG1) | 1 | - | - | - |
| Hamad et al. (1994) | [112] | SEB | 2 (IgG1) | 2 (IgG1 and IgG2b) | - | - | - | |
| Pang et al. (2000) | [113] | SEB | 1 (IgG1) | - | - | - | - | |
| Tilahun et al. (2010) | [114] | SEB | 21 (IgG1) | - | - | - | - | |
| Larkin et al. (2010) | [115] | SEB | 4 | 6 | - | - | mouse (BALB/c) | |
| Drozdowski et al. (2010) | [116] | SEB | 3 (IgG1) | - | - | - | mouse (BALB/c) | |
| Varshnev et al. (2011) | [117] | SEB | 3 (IgM, IgG1, IgA) | 8 (62.5% IgG1, 37.5% IgG2a) | - | - | mouse (BALB/c) |
Figure 1.
Distribution of mAbs classified as protective, indifferent, or disease-enhancing. (A) mAb studies divided into two groups. Group A consists of studies that describe multiple categories of mAbs (among protective, indifferent, and disease-enhancing). Group B consists of studies that describe only a single category of mAbs. n = 97 studies; (B) Distribution of mAbs with different activities from Group A studies. n = 2559 mAbs from 54 studies; (C) Distribution of mAbs with different activities from Group B studies. n = 111 mAbs from 43 studies; (D) Distribution of studies on protective, indifferent, or disease-enhancing mAbs.
Figure 1.
Distribution of mAbs classified as protective, indifferent, or disease-enhancing. (A) mAb studies divided into two groups. Group A consists of studies that describe multiple categories of mAbs (among protective, indifferent, and disease-enhancing). Group B consists of studies that describe only a single category of mAbs. n = 97 studies; (B) Distribution of mAbs with different activities from Group A studies. n = 2559 mAbs from 54 studies; (C) Distribution of mAbs with different activities from Group B studies. n = 111 mAbs from 43 studies; (D) Distribution of studies on protective, indifferent, or disease-enhancing mAbs.
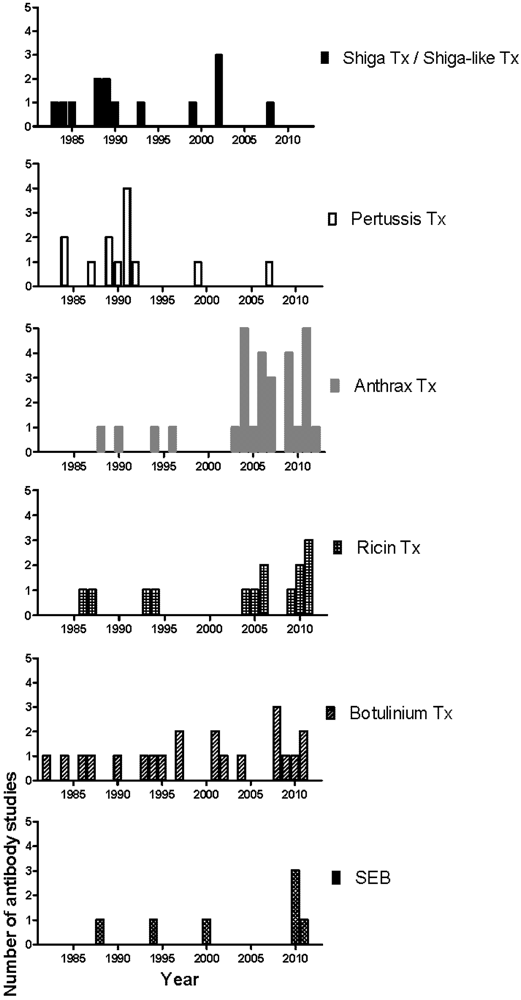
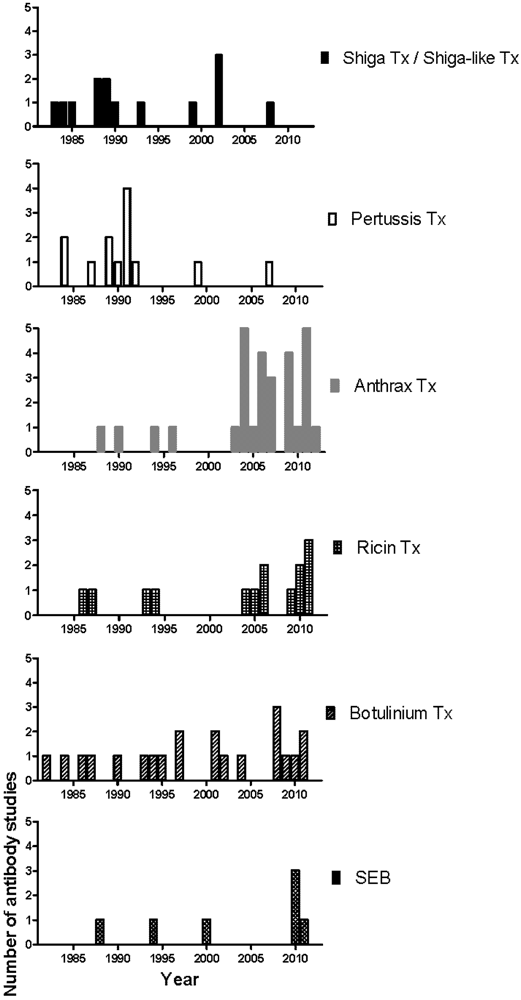
The first phase of mAb development against most toxins began in the 80s and slowed down in the early 90s, a result from the wide application of the hybridoma technology and the successful isolation of toxins and their components (Figure 2). The second phase began after the increase of research spending on biological warfare toxins and the approval of the Project BioShield Act in 2004 that specifically aimed “to provide protections and countermeasures against chemical, radiological, or nuclear agents that may be in a terrorist attack against the United States by giving the National Institutes of Health (NIH) contracting flexibility, infrastructure improvements, and expediting the scientific peer review process, and streamlining the Food and Drug Administration (FDA) approval process of countermeasures” [118]. Consequently, the number of mAb studies on National Institute of Allergy and Infectious Diseases (NIAID) biodefense category A and B priority pathogens and their toxins, such as anthrax toxin, ricin toxin and Staphylococcus enterotoxin B (SEB) increased significantly (Figure 2) [118]. Many of these mAbs were further chimerized and humanized (data not shown). In 2009, Human Genome Sciences delivered 20,000 doses of Raxibacumab, a human IgG1 mAb as a treatment of inhalation anthrax, to the US Strategic National Stockpile, and an additional 45,000 doses were ordered later in the same year [119,120]. In contrast, for diseases with a less obvious threat profile and better vaccine/treatment such as those mediated by Shiga toxin, Shiga-like toxin, and pertussis toxin, the mAb development has slowed down since the late 90s (Figure 2). Progress in the generation of prophylactic and therapeutics mAbs against biodefense pathogens has recently been reviewed [121].
Figure 2.
Chronological distribution of mAb studies for individual toxin.
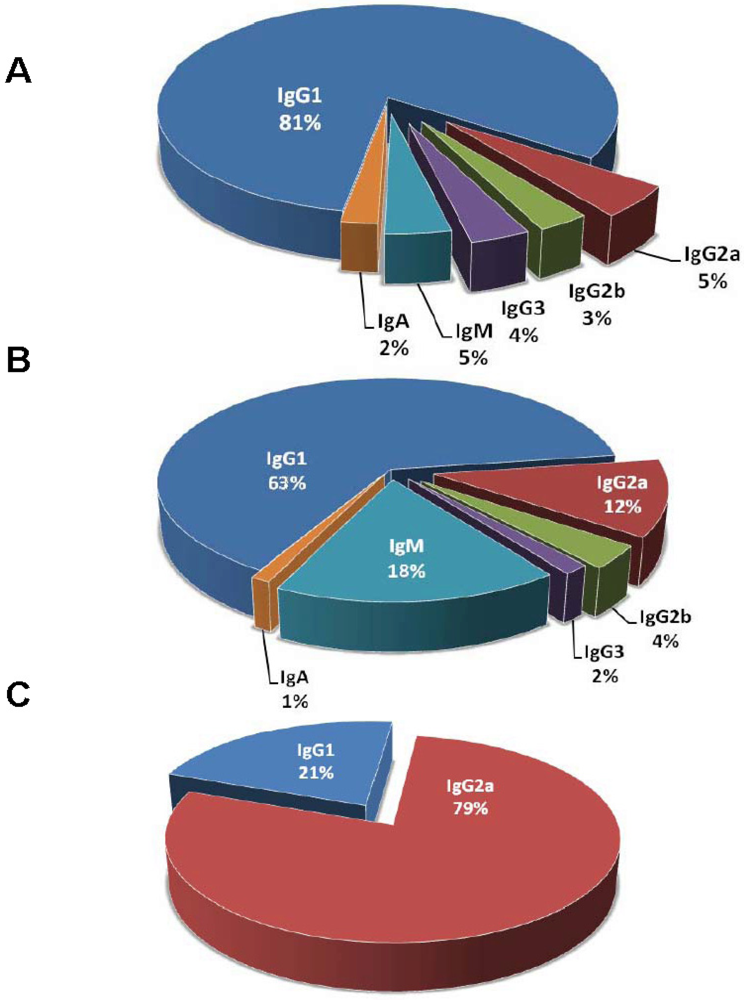
Isotype function is important in designing antibody therapeutics and the alteration of isotype subclass can increase protective efficacy [122,123]. The reported percentage of IgG for protective and indifferent mAbs listed in Table 1 is 93% and 81%, respectively (Figure 3). There are about 5 fold more IgM for indifferent mAbs than protective mAbs. Interestingly, 44 of the 45 listed indifferent IgM are raised against either anthrax toxin or Shiga/Shiga-like toxin. A possibility for the relative scarcity of IgM comparing to IgG raised against toxins is that protein antigens trigger T-cell dependent responses resulting in B cell memory, and consequently, IgG becomes the primary isotype of an immunoglobulin to protein antigens. IgG1 is the predominant IgG subclass (Figure 3) [124], but the choice of immunization adjuvants can skew the immune system to produce more IgG of other subclasses [125,126]. Alternatively, the scarcity of IgM may have represented a bias for IgG in preserving hybridomas or lack of screening for IgM in the hybridoma development. IgM as a pentamer with large molecular weight is more difficult to purify, which makes it less attractive as therapeutics candidate. IgA plays important roles in mucosal immunity including the respiratory and gastrointestinal tracts, with primary functions of intracellular neutralization and immune exclusion [9]. However, protective IgA mAbs against toxins are rare that they are only reported in a ricin toxin study [82]. It should be noted that the nomenclature of IgG subclasses between human and mouse is different [127]. Although there is no exact correspondence in function between murine and human isotypes, human IgG1 is generally thought to be analogous to murine IgG2a or IgG2c depending on the mouse strains while human IgG2, IgG3, and IgG4 are considered analogous to murine IgG3, IgG2b, and IgG1, respectively. Although many murine mAbs have been used therapeutically in humans, advances in antibody engineering technologies allow the construction of chimeric and humanized antibodies which have the advantage of being less immunogenic and confer human constant region function [128].
Figure 3.
Distribution of mAb isotypes. (A) Isotype distribution of protective mAbs. n = 222 mAbs from 67 studies; (B) Isotype distribution of indifferent mAbs. n = 252 mAbs from 30 studies; (C) Isotype distribution of disease-enhancing mAbs. n = 28 mAbs from 3 studies. Only mAbs with defined isotypes are counted.
Figure 3.
Distribution of mAb isotypes. (A) Isotype distribution of protective mAbs. n = 222 mAbs from 67 studies; (B) Isotype distribution of indifferent mAbs. n = 252 mAbs from 30 studies; (C) Isotype distribution of disease-enhancing mAbs. n = 28 mAbs from 3 studies. Only mAbs with defined isotypes are counted.

3. Caveats of Protection Studies
Several caveats are observed from the mAb studies listed in Table 1, which may render the results difficult to interpret and compare. Some of them are discussed below, and the aims are to highlight the diversity of methods and experimental parameters in mAb works, and help guiding the design of future protection studies.
3.1. Definition of Protection
It is sometimes difficult to define a mAb as protective, since protection may not be a clear cut phenomenon. It should also be noted that “neutralization” is a term that is generally reserved for interference with toxin action in an in vitro assays, while “protection” is a favorable outcome observed from in vivo studies. Pragmatically, protection can be defined as any gain of survival, viability, or fitness comparing to the control intoxicated condition. While any arbitrary percentage of increased survival can be defined as protection, the studies reviewed usually used a value ranging from 30% to 100%. The large range can then be divided into finer categories as “protective”, “slightly protective”, and “minimally protective”, indicating that the “protection” of mAbs is also relative to each other. Too little mAb cannot neutralize enough toxin to give protection, while too much mAb may result in prozone-like effects that reverse the protection observed with smaller amounts of mAb [129,130]. Consequently, most studies describe the mAb concentration being used to better standardize the protective efficacy of mAbs in the field, even though properties of an immunoglobulin such as binding affinity [131], avidity [132], degree of glycosylation [133], and effect of Fc region [123] are not as easily and commonly compared.
3.2. Toxin and Bacterium Inoculum
The toxin concentration in an assay directly affects the readout of mAb efficacy. If too much toxin is used, the fine resolution of slightly protective mAbs may be lost, and vice versa. To detect the disease or toxin enhancing effect of a given mAb, a toxin concentration that results in relatively higher percentage of viability should be used. Typically, a toxin concentration that gives about 50% viability is a reasonable starting point to screen for both mAb-mediated protection and disease enhancement. To compare the efficacy between mAbs in different studies, it is important to consider the toxin and bacterial inoculum. For example, a fully protective mAb in one study may not have higher efficacy than a slightly protective mAb in another study that uses 1.5-fold more toxin in the system. In fact, the concentration of toxin can vary by as much as 5-fold in different studies [18,19]. However, this problem can be ameliorated by the inclusion of standardization that correlates toxin concentration and toxicity [18], since this allows comparison of results across studies. Another advantage of this measure is to minimize the uncertainty due to toxin shelf life and batch-to-batch difference. Side-by-side experiments can be conducted for the comparison of mAbs to reduce the effect of toxin quality on the neutralization assay.
3.3. Screening Methods
Based on the need and availability, mAb studies employed different screening methods to test for mAb efficacy. Different methods can influence the efficacy readout, which makes the comparison of studies, even for the same toxin, challenging. For example, ADP-ribosylating, hemagglutinating, T-cell mitogenic, CHO cell-clustering, histamine-sensitizing, islet-activating, and leukocytosis-promoting activities have been used to define the efficacy of a mAb against pertussis toxin [134]. Similarly, mAb activity against anthrax toxin can be accessed by inhibition of anthrax toxin receptor and capillary morphogenesis protein (CMG2) binding, inhibition of pore formation, and inhibition of lethal toxin-mediated killing of macrophage-like cells [135]. Further complicating comparisons, a mAb may mediate protection in some but not all of the assays. The use of different cell lines can also complicate the comparisons of results from in vitro studies. Consequently, in vivo studies may be a better measure of mAb efficacy, and numerous studies have shown that mAbs reported to be neutralizing in vitro can fail to protect in animal models. 74.2% of neutralizing mAbs measured with in vitro assays also protect in animal model (Table 1). Yet, different species and strains have been used as animal models (Table 1), a common practice that contributes to the difficulty in comparing mAb across studies, since the genetic background affects the susceptibility to toxin [136]. The choice of animal model may be very different to the animal from which the antibody was made, with the examples of the testing of mAbs generated from chimpanzees in a rat model [62,70]. Furthermore, a single dose instead of a dose response is usually tested in most animal studies, an omission that could be very important if the former does not capture the optimum protection window for a particular toxin. In general, each of the screening assays plays a special role in revealing mAb efficacy. While in vitro studies can be used as the primary screening of potential mAb candidate and reveal the action of mechanism for mAb-mediated neutralization, the result of in vivo studies is still considered as the primary evidence for mAb efficacy. However, it is noteworthy that in vitro neutralization tests rely on interfering with mechanism that is part of the toxigenic process and that positive results may or may not translate into in vivo effects. Furthermore, mAbs whose function requires host factors such as cytokine, cell surface receptor, and complement to mediate a protective effect will fail to neutralize toxin or virus in a system where these components are lacking but could still be protective in animal model.
3.4. Interactions of Multiple Monoclonal Antibodies
Since mAbs became available, most investigators have assumed that the effect of antibody can be defined by the immunoglobulin in isolation—mAbs are screened alone and characterized as protective, indifferent, and disease-enhancing. While this approach simplifies the study of antibody-mediated immunity, it neglects the fact that naturally-occurring antibody responses are complex polyclonal mixtures with diverse specificities and properties. The outcome of the interaction between multiple antibodies and toxin is complex and not easily predictable. In this regard, the combination of multiple protective mAbs that target different epitopes on the same antigen can synergize protection [43,137,138]. Mixing of mAbs that target anthrax toxin components, protective antigen and lethal factor, respectively, augments protection against toxin challenge [58]. A mixture of two non-protective mAbs rescued mice from Staphylococcal enterotoxin B-induced lethal shock by a mechanism whereby the binding of one mAb promotes structural change that allows the second one to neutralize the toxin [117]. The fact that antibody-mediated immunity is often protective, contrasts with the observation that most toxin-binding mAbs described in the literature are non-protective (Figure 1B).
4. Conclusions
The properties of mAb that include high specificity, purity, and ability to enhance host immune system make immunoglobulins attractive candidates for the next generation of therapeutics. While mAb development is primarily focused on the candidates with the greatest medicinal potential, one should be cautious that the mAbs categorized as non-protective on single antibody screen, which comprise the majority of naturally-produced antibodies (Figure 1B), could be false-negative results. In other words, it is possible that those antibodies are biologically active when they function in combination with other antibodies. Combination testing could reveal new properties of these so called non-protective or indifferent antibodies with the caveat that evaluating antibodies in combinations would increase considerable cost and complexity to any screen for useful mAbs. However, the promising outcomes from mAbs cocktail against viral diseases and synergy between mAb and anti-infective drugs [9] suggest that this approach may unveil the potential of previously neglected mAbs. Perhaps the next generation technology will shift the focus of antibody development from single antibody screening to combination testing. Finally, continuous advancement of mAb engineering in the areas of mAb bispecificity, glycosylation, and Fc region modification will lay a solid foundation for the future mAb therapeutic era against toxin-mediated diseases.
Acknowledgements
This work was supported by grants from the Department of Defense (Proposal Log #07164001; Award No. W81XWH08-01-0011), from the Northeast Biodefense Center (5U54AI05715807 to Lipkin, W.I.). In addition, Arturo Casadevall is also supported by HL059842-3, A1033774, A1052733, and AI033142.
Conflict of Interest
The authors declare no conflict of interest.
References
- Brock, T.D. Milestones in Microbiology: 1556 to 1940; ASM Press: Washington, DC, USA, 1998. [Google Scholar]
- Casadevall, A. The third age of antimicrobial therapy. Clin. Infect. Dis. 2006, 42, 1414–1416. [Google Scholar] [CrossRef]
- Guarner, F.; Malagelada, J.R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Kohler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef]
- Casadevall, A.; Scharff, M.D. Serum therapy revisited: Animal models of infection and development of passive antibody therapy. Antimicrob. Agents Chemother. 1994, 38, 1695–1702. [Google Scholar]
- Khazaeli, M.B.; Conry, R.M.; LoBuglio, A.F. Human immune response to monoclonal antibodies. J. Immunother. Emphasis. Tumor. Immunol. 1994, 15, 42–52. [Google Scholar] [CrossRef]
- Lang, A.B.; Cryz, S.J., Jr.; Schurch, U.; Ganss, M.T.; Bruderer, U. Immunotherapy with human monoclonal antibodies. Fragment a specificity of polyclonal and monoclonal antibodies is crucial for full protection against tetanus toxin. J. Immunol. 1993, 151, 466–472. [Google Scholar]
- Saylor, C.; Dadachova, E.; Casadevall, A. Monoclonal antibody-based therapies for microbial diseases. Vaccine 2009, 27, S38–S46. [Google Scholar]
- Berry, J.D.; Gaudet, R.G. Antibodies in infectious diseases: Polyclonals, monoclonals and niche biotechnology. N. Biotechnol. 2011, 28, 489–501. [Google Scholar]
- Laffly, E.; Danjou, L.; Condemine, F.; Vidal, D.; Drouet, E.; Lefranc, M.P.; Bottex, C.; Thullier, P. Selection of a macaque fab with framework regions like those in humans, high affinity, and ability to neutralize the protective antigen (pa) of bacillus anthracis by binding to the segment of pa between residues 686 and 694. Antimicrob. Agents Chemother. 2005, 49, 3414–3420. [Google Scholar]
- Pelat, T.; Hust, M.; Laffly, E.; Condemine, F.; Bottex, C.; Vidal, D.; Lefranc, M.P.; Dubel, S.; Thullier, P. High-affinity, human antibody-like antibody fragment (single-chain variable fragment) neutralizing the lethal factor (lf) of bacillus anthracis by inhibiting protective antigen-lf complex formation. Antimicrob. Agents Chemother. 2007, 51, 2758–2764. [Google Scholar] [CrossRef]
- Chahboun, S.; Hust, M.; Liu, Y.; Pelat, T.; Miethe, S.; Helmsing, S.; Jones, R.G.; Sesardic, D.; Thullier, P. Isolation of a nanomolar scfv inhibiting the endopeptidase activity of botulinum toxin a, by single-round panning of an immune phage-displayed library of macaque origin. BMC Biotechnol. 2011, 11, 113. [Google Scholar] [CrossRef]
- Peiris, J.S.; Porterfield, J.S. Antibody-mediated enhancement of flavivirus replication in macrophage-like cell lines. Nature 1979, 282, 509–511. [Google Scholar] [CrossRef]
- Takeda, A.; Tuazon, C.U.; Ennis, F.A. Antibody-enhanced infection by hiv-1 via fc receptor-mediated entry. Science 1988, 242, 580–583. [Google Scholar]
- Dejnirattisai, W.; Jumnainsong, A.; Onsirisakul, N.; Fitton, P.; Vasanawathana, S.; Limpitikul, W.; Puttikhunt, C.; Edwards, C.; Duangchinda, T.; Supasa, S.; et al. Cross-reacting antibodies enhance dengue virus infection in humans. Science 2010, 328, 745–748. [Google Scholar]
- Takada, A.; Watanabe, S.; Okazaki, K.; Kida, H.; Kawaoka, Y. Infectivity-enhancing antibodies to ebola virus glycoprotein. J. Virol. 2001, 75, 2324–2330. [Google Scholar] [CrossRef]
- Belova, E.V.; Dubilei, S.A.; Kravchenko, T.B.; Kolesnikov, A.V.; Zakharova, M.; Shemiakin, I.G. Monoclonal antibodies to B. Anthracis protective antigen are capable to neutralize and to enhance the anthrax lethal toxin action in vitro. Mol. Gen. Mikrobiol. Virusol. 2004, 21–26. [Google Scholar]
- Mohamed, N.; Li, J.; Ferreira, C.S.; Little, S.F.; Friedlander, A.M.; Spitalny, G.L.; Casey, L.S. Enhancement of anthrax lethal toxin cytotoxicity: A subset of monoclonal antibodies against protective antigen increases lethal toxin-mediated killing of murine macrophages. Infect. Immun. 2004, 72, 3276–3283. [Google Scholar]
- Little, S.F.; Webster, W.M.; Fisher, D.E. Monoclonal antibodies directed against protective antigen of bacillus anthracis enhance lethal toxin activity in vivo. FEMS Immunol. Med. Microbiol. 2011, 62, 11–22. [Google Scholar] [CrossRef]
- Colombatti, M.; Pezzini, A.; Colombatti, A. Monoclonal antibodies against ricin: Effects on toxin function. Hybridoma 1986, 5, 9–19. [Google Scholar] [CrossRef]
- Maddaloni, M.; Cooke, C.; Wilkinson, R.; Stout, A.V.; Eng, L.; Pincus, S.H. Immunological characteristics associated with the protective efficacy of antibodies to ricin. J. Immunol. 2004, 172, 6221–6228. [Google Scholar]
- Reichert, J.M. Antibody-based therapeutics to watch in 2011. MAbs 2011, 3, 76–99. [Google Scholar] [CrossRef]
- The Impact-RSV Study Group. Palivizumab, a humanized respiratory syncytial virus monoclonal antibody, reduces hospitalization from respiratory syncytial virus infection in high-risk infants. The impact-rsv study group. Pediatrics 1998, 102, 531–537. [CrossRef]
- Storey, S. Respiratory syncytial virus market. Nat. Rev. Drug. Discov. 2010, 9, 15–16. [Google Scholar] [CrossRef]
- Chames, P.; Van Regenmortel, M.; Weiss, E.; Baty, D. Therapeutic antibodies: Successes, limitations and hopes for the future. Br. J. Pharmacol. 2009, 157, 220–233. [Google Scholar] [CrossRef]
- Griffin, D.E.; Gentry, M.K.; Brown, J.E. Isolation and characterization of monoclonal antibodies to shiga toxin. Infect. Immun. 1983, 41, 430–433. [Google Scholar]
- Donohue-Rolfe, A.; Keusch, G.T.; Edson, C.; Thorley-Lawson, D.; Jacewicz, M. Pathogenesis of shigella diarrhea. IX. Simplified high yield purification of shigella toxin and characterization of subunit composition and function by the use of subunit-specific monoclonal and polyclonal antibodies. J. Exp. Med. 1984, 160, 1767–1781. [Google Scholar] [CrossRef]
- Strockbine, N.A.; Marques, L.R.; Holmes, R.K.; O'Brien, A.D. Characterization of monoclonal antibodies against shiga-like toxin from escherichia coli. Infect. Immun. 1985, 50, 695–700. [Google Scholar]
- Downes, F.P.; Barrett, T.J.; Green, J.H.; Aloisio, C.H.; Spika, J.S.; Strockbine, N.A.; Wachsmuth, I.K. Affinity purification and characterization of shiga-like toxin ii and production of toxin-specific monoclonal antibodies. Infect. Immun. 1988, 56, 1926–1933. [Google Scholar]
- Perera, L.P.; Marques, L.R.; O’Brien, A.D. Isolation and characterization of monoclonal antibodies to shiga-like toxin ii of enterohemorrhagic escherichia coli and use of the monoclonal antibodies in a colony enzyme-linked immunosorbent assay. J. Clin. Microbiol. 1988, 26, 2127–2131. [Google Scholar]
- Donohue-Rolfe, A.; Acheson, D.W.; Kane, A.V.; Keusch, G.T. Purification of shiga toxin and shiga-like toxins i and ii by receptor analog affinity chromatography with immobilized p1 glycoprotein and production of cross-reactive monoclonal antibodies. Infect. Immun. 1989, 57, 3888–3893. [Google Scholar]
- Padhye, V.V.; Zhao, T.; Doyle, M.P. Production and characterisation of monoclonal antibodies to verotoxins 1 and 2 from escherichia coli of serotype o 157:H7. J. Med. Microbiol. 1989, 30, 219–226. [Google Scholar] [CrossRef]
- Islam, M.S.; Stimson, W.H. Production and characterization of monoclonal antibodies with therapeutic potential against shiga toxin. J. Clin. Lab. Immunol. 1990, 33, 11–16. [Google Scholar]
- Qadri, F.; Azim, T.; Hossain, A.; Islam, D.; Mondol, G.; Faruque, S.M.; Albert, M.J. A monoclonal antibody to shigella dysenteriae serotype 13 cross-reacting with shiga toxin. FEMS Microbiol. Lett. 1993, 107, 343–347. [Google Scholar] [CrossRef]
- Nakao, H.; Kiyokawa, N.; Fujimoto, J.; Yamasaki, S.; Takeda, T. Monoclonal antibody to shiga toxin 2 which blocks receptor binding and neutralizes cytotoxicity. Infect. Immun. 1999, 67, 5717–5722. [Google Scholar]
- Mukherjee, J.; Chios, K.; Fishwild, D.; Hudson, D.; O'Donnell, S.; Rich, S.M.; Donohue-Rolfe, A.; Tzipori, S. Production and characterization of protective human antibodies against shiga toxin 1. Infect. Immun. 2002, 70, 5896–5899. [Google Scholar]
- Mukherjee, J.; Chios, K.; Fishwild, D.; Hudson, D.; O’Donnell, S.; Rich, S.M.; Donohue-Rolfe, A.; Tzipori, S. Human stx2-specific monoclonal antibodies prevent systemic complications of escherichia coli o157:H7 infection. Infect. Immun. 2002, 70, 612–619. [Google Scholar] [CrossRef]
- Nakao, H.; Kataoka, C.; Kiyokawa, N.; Fujimoto, J.; Yamasaki, S.; Takeda, T. Monoclonal antibody to shiga toxin 1, which blocks receptor binding and neutralizes cytotoxicity. Microbiol. Immunol. 2002, 46, 777–780. [Google Scholar]
- Tanikawa, T.; Ishikawa, T.; Maekawa, T.; Kuronane, K.; Imai, Y. Characterization of monoclonal immunoglobulin a and g against shiga toxin binding subunits produced by intranasal immunization. Scand J. Immunol. 2008, 68, 414–422. [Google Scholar] [CrossRef]
- Sato, H.; Ito, A.; Chiba, J.; Sato, Y. Monoclonal antibody against pertussis toxin: Effect on toxin activity and pertussis infections. Infect. Immun. 1984, 46, 422–428. [Google Scholar]
- Frank, D.W.; Parker, C.D. Interaction of monoclonal antibodies with pertussis toxin and its subunits. Infect. Immun. 1984, 46, 195–201. [Google Scholar]
- Sato, H.; Sato, Y.; Ito, A.; Ohishi, I. Effect of monoclonal antibody to pertussis toxin on toxin activity. Infect. Immun. 1987, 55, 909–915. [Google Scholar]
- Kenimer, J.G.; Kim, K.J.; Probst, P.G.; Manclark, C.R.; Burstyn, D.G.; Cowell, J.L. Monoclonal antibodies to pertussis toxin: Utilization as probes of toxin function. Hybridoma 1989, 8, 37–51. [Google Scholar] [CrossRef]
- Lang, A.B.; Ganss, M.T.; Cryz, S.J., Jr. , Monoclonal antibodies that define neutralizing epitopes of pertussis toxin: Conformational dependence and epitope mapping. Infect. Immun. 1989, 57, 2660–2665. [Google Scholar]
- Sato, H.; Sato, Y. Protective activities in mice of monoclonal antibodies against pertussis toxin. Infect. Immun. 1990, 58, 3369–3374. [Google Scholar]
- Halperin, S.A.; Issekutz, T.B.; Kasina, A. Modulation of bordetella pertussis infection with monoclonal antibodies to pertussis toxin. J. Infect. Dis. 1991, 163, 355–361. [Google Scholar] [CrossRef]
- Kenimer, J.G.; Probst, P.G.; Karpas, A.B.; Burns, D.L.; Kaslow, H.R. Monoclonal antibodies against the enzymatic subunit of both pertussis and cholera toxins. Dev. Biol. Stand 1991, 73, 133–141. [Google Scholar]
- Sato, H.; Sato, Y.; Ohishi, I. Comparison of pertussis toxin (pt)-neutralizing activities and mouse-protective activities of anti-pt mouse monoclonal antibodies. Infect. Immun. 1991, 59, 3832–3835. [Google Scholar]
- Walker, M.J.; Wehland, J.; Timmis, K.N.; Raupach, B.; Schmidt, M.A. Characterization of murine monoclonal antibodies that recognize defined epitopes of pertussis toxin and neutralize its toxic effect on chinese hamster ovary cells. Infect. Immun. 1991, 59, 4249–4251. [Google Scholar]
- Zaccolo, M.; Roggero, S.; Armellini, D.; Pegoraro, L.; Rappuoli, R.; Malavasi, F. Generation of human monoclonal antibodies that confer protection against pertussis toxin. Infect. Immun. 1992, 60, 1258–1260. [Google Scholar]
- Lee, S.J.; Gray, M.C.; Guo, L.; Sebo, P.; Hewlett, E.L. Epitope mapping of monoclonal antibodies against bordetella pertussis adenylate cyclase toxin. Infect. Immun. 1999, 67, 2090–2095. [Google Scholar]
- Pootong, A.; Budhirakkul, P.; Tongtawe, P.; Tapchaisri, P.; Chongsa-nguan, M.; Chaicumpa, W. Monoclonal antibody that neutralizes pertussis toxin activities. Asian Pac. J. Allergy Immunol. 2007, 25, 37–45. [Google Scholar]
- Little, S.F.; Leppla, S.H.; Cora, E. Production and characterization of monoclonal antibodies to the protective antigen component of bacillus anthracis toxin. Infect. Immun. 1988, 56, 1807–1813. [Google Scholar]
- Little, S.F.; Leppla, S.H.; Friedlander, A.M. Production and characterization of monoclonal antibodies against the lethal factor component of bacillus anthracis lethal toxin. Infect. Immun. 1990, 58, 1606–1613. [Google Scholar]
- Little, S.F.; Leppla, S.H.; Burnett, J.W.; Friedlander, A.M. Structure-function analysis of bacillus anthracis edema factor by using monoclonal antibodies. Biochem. Biophys. Res. Commun. 1994, 199, 676–682. [Google Scholar] [CrossRef]
- Little, S.F.; Novak, J.M.; Lowe, J.R.; Leppla, S.H.; Singh, Y.; Klimpel, K.R.; Lidgerding, B.C.; Friedlander, A.M. Characterization of lethal factor binding and cell receptor binding domains of protective antigen of bacillus anthracis using monoclonal antibodies. Microbiology 1996, 142 (Pt 3), 707–715. [Google Scholar]
- Zhao, P.; Liang, X.; Kalbfleisch, J.; Koo, H.M.; Cao, B. Neutralizing monoclonal antibody against anthrax lethal factor inhibits intoxication in a mouse model. Hum. Antibodies 2003, 12, 129–135. [Google Scholar]
- Brossier, F.; Levy, M.; Landier, A.; Lafaye, P.; Mock, M. Functional analysis of bacillus anthracis protective antigen by using neutralizing monoclonal antibodies. Infect. Immun. 2004, 72, 6313–6317. [Google Scholar] [CrossRef]
- Kozel, T.R.; Murphy, W.J.; Brandt, S.; Blazar, B.R.; Lovchik, J.A.; Thorkildson, P.; Percival, A.; Lyons, C.R. Mabs to bacillus anthracis capsular antigen for immunoprotection in anthrax and detection of antigenemia. Proc. Nat. Acad. Sci. USA 2004, 101, 5042–5047. [Google Scholar]
- Sawada-Hirai, R.; Jiang, I.; Wang, F.; Sun, S.M.; Nedellec, R.; Ruther, P.; Alvarez, A.; Millis, D.; Morrow, P.R.; Kang, A.S. Human anti-anthrax protective antigen neutralizing monoclonal antibodies derived from donors vaccinated with anthrax vaccine adsorbed. J. Immune. Based Ther. Vaccines 2004, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Lim, N.K.; Kim, J.H.; Oh, M.S.; Lee, S.; Kim, S.Y.; Kim, K.S.; Kang, H.J.; Hong, H.J.; Inn, K.S. An anthrax lethal factor-neutralizing monoclonal antibody protects rats before and after challenge with anthrax toxin. Infect. Immun. 2005, 73, 6547–6551. [Google Scholar]
- Chen, Z.; Moayeri, M.; Zhou, Y.H.; Leppla, S.; Emerson, S.; Sebrell, A.; Yu, F.; Svitel, J.; Schuck, P.; St Claire, M.; et al. Efficient neutralization of anthrax toxin by chimpanzee monoclonal antibodies against protective antigen. J. Infect. Dis. 2006, 193, 625–633. [Google Scholar] [CrossRef]
- Gubbins, M.J.; Berry, J.D.; Corbett, C.R.; Mogridge, J.; Yuan, X.Y.; Schmidt, L.; Nicolas, B.; Kabani, A.; Tsang, R.S. Production and characterization of neutralizing monoclonal antibodies that recognize an epitope in domain 2 of bacillus anthracis protective antigen. FEMS Immunol. Med. Microbiol. 2006, 47, 436–443. [Google Scholar] [CrossRef]
- Rivera, J.; Nakouzi, A.; Abboud, N.; Revskaya, E.; Goldman, D.; Collier, R.J.; Dadachova, E.; Casadevall, A. A monoclonal antibody to bacillus anthracis protective antigen defines a neutralizing epitope in domain 1. Infect. Immun. 2006, 74, 4149–4156. [Google Scholar]
- Vitale, L.; Blanset, D.; Lowy, I.; O’Neill, T.; Goldstein, J.; Little, S.F.; Andrews, G.P.; Dorough, G.; Taylor, R.K.; Keler, T. Prophylaxis and therapy of inhalational anthrax by a novel monoclonal antibody to protective antigen that mimics vaccine-induced immunity. Infect. Immun. 2006, 74, 5840–5847. [Google Scholar]
- Albrecht, M.T.; Li, H.; Williamson, E.D.; LeButt, C.S.; Flick-Smith, H.C.; Quinn, C.P.; Westra, H.; Galloway, D.; Mateczun, A.; Goldman, S.; et al. Human monoclonal antibodies against anthrax lethal factor and protective antigen act independently to protect against bacillus anthracis infection and enhance endogenous immunity to anthrax. Infect. Immun. 2007, 75, 5425–5433. [Google Scholar]
- Kozel, T.R.; Thorkildson, P.; Brandt, S.; Welch, W.H.; Lovchik, J.A.; AuCoin, D.P.; Vilai, J.; Lyons, C.R. Protective and immunochemical activities of monoclonal antibodies reactive with the bacillus anthracis polypeptide capsule. Infect. Immun. 2007, 75, 152–163. [Google Scholar]
- Staats, H.F.; Alam, S.M.; Scearce, R.M.; Kirwan, S.M.; Zhang, J.X.; Gwinn, W.M.; Haynes, B.F. In vitro and in vivo characterization of anthrax anti-protective antigen and anti-lethal factor monoclonal antibodies after passive transfer in a mouse lethal toxin challenge model to define correlates of immunity. Infect. Immun. 2007, 75, 5443–5452. [Google Scholar] [CrossRef]
- Abboud, N.; De Jesus, M.; Nakouzi, A.; Cordero, R.J.; Pujato, M.; Fiser, A.; Rivera, J.; Casadevall, A. Identification of linear epitopes in bacillus anthracis protective antigen bound by neutralizing antibodies. J. Biol. Chem. 2009, 284, 25077–25086. [Google Scholar]
- Chen, Z.; Moayeri, M.; Crown, D.; Emerson, S.; Gorshkova, I.; Schuck, P.; Leppla, S.H.; Purcell, R.H. Novel chimpanzee/human monoclonal antibodies that neutralize anthrax lethal factor, and evidence for possible synergy with anti-protective antigen antibody. Infect. Immun. 2009, 77, 3902–3908. [Google Scholar]
- Kelly-Cirino, C.D.; Mantis, N.J. Neutralizing monoclonal antibodies directed against defined linear epitopes on domain 4 of anthrax protective antigen. Infect. Immun. 2009, 77, 4859–4867. [Google Scholar] [CrossRef]
- Rosenfeld, R.; Marcus, H.; Ben-Arie, E.; Lachmi, B.E.; Mechaly, A.; Reuveny, S.; Gat, O.; Mazor, O.; Ordentlich, A. Isolation and chimerization of a highly neutralizing antibody conferring passive protection against lethal bacillus anthracis infection. PLoS One 2009, 4, e6351. [Google Scholar]
- Winterroth, L.; Rivera, J.; Nakouzi, A.S.; Dadachova, E.; Casadevall, A. Neutralizing monoclonal antibody to edema toxin and its effect on murine anthrax. Infect. Immun. 2010, 78, 2890–2898. [Google Scholar] [CrossRef]
- Chen, Z.; Schneerson, R.; Lovchik, J.; Lyons, C.R.; Zhao, H.; Dai, Z.; Kubler-Kielb, J.; Leppla, S.H.; Purcell, R.H. Pre- and postexposure protection against virulent anthrax infection in mice by humanized monoclonal antibodies to bacillus anthracis capsule. Proc. Nat. Acad. Sci. USA 2011, 108, 739–744. [Google Scholar]
- Kulshreshtha, P.; Bhatnagar, R. Inhibition of anthrax toxins with a bispecific monoclonal antibody that cross reacts with edema factor as well as lethal factor of bacillus anthracis. Mol. Immunol. 2011, 48, 1958–1965. [Google Scholar] [CrossRef]
- Leysath, C.E.; Chen, K.H.; Moayeri, M.; Crown, D.; Fattah, R.; Chen, Z.; Das, S.R.; Purcell, R.H.; Leppla, S.H. Mouse monoclonal antibodies to anthrax edema factor protect against infection. Infect. Immun. 2011, 79, 4609–4616. [Google Scholar]
- vor dem Esche, U.; Huber, M.; Zgaga-Griesz, A.; Grunow, R.; Beyer, W.; Hahn, U.; Bessler, W.G. Passive vaccination with a human monoclonal antibody: Generation of antibodies and studies for efficacy in bacillus anthracis infections. Immunobiology 2011, 216, 847–853. [Google Scholar] [CrossRef]
- Colombatti, M.; Johnson, V.G.; Skopicki, H.A.; Fendley, B.; Lewis, M.S.; Youle, R.J. Identification and characterization of a monoclonal antibody recognizing a galactose-binding domain of the toxin ricin. J. Immunol. 1987, 138, 3339–3344. [Google Scholar]
- Chanh, T.C.; Romanowski, M.J.; Hewetson, J.F. Monoclonal antibody prophylaxis against the in vivo toxicity of ricin in mice. Immunol. Invest. 1993, 22, 63–72. [Google Scholar] [CrossRef]
- Lemley, P.V.; Amanatides, P.; Wright, D.C. Identification and characterization of a monoclonal antibody that neutralizes ricin toxicity in vitro and in vivo. Hybridoma 1994, 13, 417–421. [Google Scholar] [CrossRef]
- Dertzbaugh, M.T.; Rossi, C.A.; Paddle, B.M.; Hale, M.; Poretski, M.; Alderton, M.R. Monoclonal antibodies to ricin: In vitro inhibition of toxicity and utility as diagnostic reagents. Hybridoma (Larchmt) 2005, 24, 236–243. [Google Scholar] [CrossRef]
- Mantis, N.J.; McGuinness, C.R.; Sonuyi, O.; Edwards, G.; Farrant, S.A. Immunoglobulin a antibodies against ricin a and b subunits protect epithelial cells from ricin intoxication. Infect. Immun. 2006, 74, 3455–3462. [Google Scholar]
- McGuinness, C.R.; Mantis, N.J. Characterization of a novel high-affinity monoclonal immunoglobulin g antibody against the ricin b subunit. Infect. Immun. 2006, 74, 3463–3470. [Google Scholar] [CrossRef]
- Pelat, T.; Hust, M.; Hale, M.; Lefranc, M.P.; Dubel, S.; Thullier, P. Isolation of a human-like antibody fragment (scfv) that neutralizes ricin biological activity. BMC Biotechnol 2009, 9, 60. [Google Scholar] [CrossRef]
- Neal, L.M.; O'Hara, J.; Brey, R.N., 3rd; Mantis, N.J. A monoclonal immunoglobulin g antibody directed against an immunodominant linear epitope on the ricin a chain confers systemic and mucosal immunity to ricin. Infect. Immun. 2010, 78, 552–561. [Google Scholar] [CrossRef]
- O'Hara, J.M.; Neal, L.M.; McCarthy, E.A.; Kasten-Jolly, J.A.; Brey, R.N.3rd; Mantis, N.J. Folding domains within the ricin toxin a subunit as targets of protective antibodies. Vaccine 2010, 28, 7035–7046. [Google Scholar]
- Dai, J.; Zhao, L.; Yang, H.; Guo, H.; Fan, K.; Wang, H.; Qian, W.; Zhang, D.; Li, B.; Guo, Y. Identification of a novel functional domain of ricin responsible for its potent toxicity. J. Biol. Chem. 2011, 286, 12166–12171. [Google Scholar]
- Prigent, J.; Panigai, L.; Lamourette, P.; Sauvaire, D.; Devilliers, K.; Plaisance, M.; Volland, H.; Creminon, C.; Simon, S. Neutralising antibodies against ricin toxin. PLoS One 2011, 6, e20166. [Google Scholar]
- Yermakova, A.; Mantis, N.J. Protective immunity to ricin toxin conferred by antibodies against the toxin's binding subunit (rtb). Vaccine 2011, 29, 7925–7935. [Google Scholar] [CrossRef]
- Oguma, K.; Agui, T.; Syuto, B.; Kimura, K.; Iida, H.; Kubo, S. Four different monoclonal antibodies against type c1 toxin of clostridium botulinum. Infect. Immun. 1982, 38, 14–20. [Google Scholar]
- Oguma, K.; Murayama, S.; Syuto, B.; Iida, H.; Kubo, S. Analysis of antigenicity of clostridium botulinum type c1 and d toxins by polyclonal and monoclonal antibodies. Infect. Immun. 1984, 43, 584–588. [Google Scholar]
- Kozaki, S.; Kamata, Y.; Nagai, T.; Ogasawara, J.; Sakaguchi, G. The use of monoclonal antibodies to analyze the structure of clostridium botulinum type e derivative toxin. Infect. Immun. 1986, 52, 786–791. [Google Scholar]
- Ferreira, J.L.; Hamdy, M.K.; Herd, Z.L.; McCay, S.G.; Zapatka, F.A. Monoclonal antibody for the detection of clostridium botulinum type a toxin. Mol. Cell Probes. 1987, 1, 337–345. [Google Scholar] [CrossRef]
- Simpson, L.L.; Kamata, Y.; Kozaki, S. Use of monoclonal antibodies as probes for the structure and biological activity of botulinum neurotoxin. J. Pharmacol. Exp. Ther. 1990, 255, 227–232. [Google Scholar]
- Toratani, S.; Sekine, N.; Katada, T.; Yokosawa, H. Production of monoclonal antibodies that inhibit adp-ribosylation of small gtp-binding proteins catalyzed by clostridium botulinum adp-ribosyltransferase c3. FEBS Lett. 1993, 324, 353–357. [Google Scholar] [CrossRef]
- Cenci Di Bello, I.; Poulain, B.; Shone, C.C.; Tauc, L.; Dolly, J.O. Antagonism of the intracellular action of botulinum neurotoxin type a with monoclonal antibodies that map to light-chain epitopes. Eur. J. Biochem. 1994, 219, 161–169. [Google Scholar] [CrossRef]
- Noah, C.W.; Poteet, S.S.; Ramos, N.C.; Perez, J.C.; Huang, S.Y. Production of monoclonal antibodies specific to clostridium botulinum type b neurotoxin. J. AOAC Int. 1995, 78, 381–385. [Google Scholar]
- Amersdorfer, P.; Wong, C.; Chen, S.; Smith, T.; Deshpande, S.; Sheridan, R.; Finnern, R.; Marks, J.D. Molecular characterization of murine humoral immune response to botulinum neurotoxin type a binding domain as assessed by using phage antibody libraries. Infect. Immun. 1997, 65, 3743–3752. [Google Scholar]
- Brown, D.R.; Lloyd, J.P.; Schmidt, J.J. Identification and characterization of a neutralizing monoclonal antibody against botulinum neurotoxin serotype f, following vaccination with active toxin. Hybridoma 1997, 16, 447–456. [Google Scholar] [CrossRef]
- Pless, D.D.; Torres, E.R.; Reinke, E.K.; Bavari, S. High-affinity, protective antibodies to the binding domain of botulinum neurotoxin type a. Infect. Immun. 2001, 69, 570–574. [Google Scholar] [CrossRef]
- Wu, H.C.; Yeh, C.T.; Huang, Y.L.; Tarn, L.J.; Lung, C.C. Characterization of neutralizing antibodies and identification of neutralizing epitope mimics on the clostridium botulinum neurotoxin type a. Appl. Environ. Microbiol. 2001, 67, 3201–3207. [Google Scholar] [CrossRef]
- Kamata, Y.; Hoshi, H.; Choki, H.; Kozaki, S. Characterization of a neutralizing monoclonal antibody against botulinum adp-ribosyltransferase, c3 exoenzyme. J. Vet. Med. Sci. 2002, 64, 767–771. [Google Scholar] [CrossRef]
- Yang, G.H.; Kim, K.S.; Kim, H.W.; Jeong, S.T.; Huh, G.H.; Kim, J.C.; Jung, H.H. Isolation and characterization of a neutralizing antibody specific to internalization domain of clostridium botulinum neurotoxin type b. Toxicon 2004, 44, 19–25. [Google Scholar] [CrossRef]
- Adekar, S.P.; Jones, R.M.; Elias, M.D.; Al-Saleem, F.H.; Root, M.J.; Simpson, L.L.; Dessain, S.K. A human monoclonal antibody that binds serotype a botulinum neurotoxin. Hybridoma (Larchmt) 2008, 27, 11–17. [Google Scholar] [CrossRef]
- Adekar, S.P.; Takahashi, T.; Jones, R.M.; Al-Saleem, F.H.; Ancharski, D.M.; Root, M.J.; Kapadnis, B.P.; Simpson, L.L.; Dessain, S.K. Neutralization of botulinum neurotoxin by a human monoclonal antibody specific for the catalytic light chain. PLoS One 2008, 3, e3023. [Google Scholar]
- Adekar, S.P.; Al-Saleem, F.H.; Elias, M.D.; Rybinski, K.A.; Simpson, L.L.; Dessain, S.K. A natural human igm antibody that neutralizes botulinum neurotoxin in vivo. Hybridoma (Larchmt) 2008, 27, 65–69. [Google Scholar] [CrossRef]
- Zhou, H.; Zhou, B.; Pellett, S.; Johnson, E.A.; Janda, K.D. Selection and characterization of a human monoclonal neutralizing antibody for clostridium botulinum neurotoxin serotype b. Bioorg. Med. Chem. Lett. 2009, 19, 662–664. [Google Scholar]
- Mazuet, C.; Dano, J.; Popoff, M.R.; Creminon, C.; Volland, H. Characterization of botulinum neurotoxin type a neutralizing monoclonal antibodies and influence of their half-lives on therapeutic activity. PLoS One 2010, 5, e12416. [Google Scholar]
- Corbett, C.R.; Ballegeer, E.; Weedmark, K.A.; Elias, M.D.; Al-Saleem, F.H.; Ancharski, D.M.; Simpson, L.L.; Berry, J.D. Epitope characterization of sero-specific monoclonal antibody to clostridium botulinum neurotoxin type a. Hybridoma (Larchmt) 2011, 30, 503–510. [Google Scholar] [CrossRef]
- Montgomery, V.A.; Smith, L.A. Diagnostic and possible therapeutic application of a monoclonal antibody (14g8) directed against botulinum type c neurotoxin. Hybridoma (Larchmt) 2011, 30, 209–216. [Google Scholar] [CrossRef]
- Lin, Y.S.; Largen, M.T.; Newcomb, J.R.; Rogers, T.J. Production and characterisation of monoclonal antibodies specific for staphylococcal enterotoxin b. J. Med. Microbiol. 1988, 27, 263–270. [Google Scholar] [CrossRef]
- Hamad, A.R.; Herman, A.; Marrack, P.; Kappler, J.W. Monoclonal antibodies defining functional sites on the toxin superantigen staphylococcal enterotoxin b. J. Exp. Med. 1994, 180, 615–621. [Google Scholar]
- Pang, L.T.; Kum, W.W.; Chow, A.W. Inhibition of staphylococcal enterotoxin b-induced lymphocyte proliferation and tumor necrosis factor alpha secretion by mab5, an anti-toxic shock syndrome toxin 1 monoclonal antibody. Infect. Immun. 2000, 68, 3261–3268. [Google Scholar] [CrossRef]
- Tilahun, M.E.; Rajagopalan, G.; Shah-Mahoney, N.; Lawlor, R.G.; Tilahun, A.Y.; Xie, C.; Natarajan, K.; Margulies, D.H.; Ratner, D.I.; Osborne, B.A.; et al. Potent neutralization of staphylococcal enterotoxin b by synergistic action of chimeric antibodies. Infect. Immun. 2010, 78, 2801–2811. [Google Scholar]
- Larkin, E.A.; Stiles, B.G.; Ulrich, R.G. Inhibition of toxic shock by human monoclonal antibodies against staphylococcal enterotoxin b. PLoS One 2010, 5, e13253. [Google Scholar]
- Drozdowski, B.; Zhou, Y.; Kline, B.; Spidel, J.; Chan, Y.Y.; Albone, E.; Turchin, H.; Chao, Q.; Henry, M.; Balogach, J.; et al. Generation and characterization of high affinity human monoclonal antibodies that neutralize staphylococcal enterotoxin b. J. Immun. Based Ther. Vaccines 2010, 8, 9. [Google Scholar] [CrossRef]
- Varshney, A.K.; Wang, X.; Cook, E.; Dutta, K.; Scharff, M.D.; Goger, M.J.; Fries, B.C. Generation, characterization, and epitope mapping of neutralizing and protective monoclonal antibodies against staphylococcal enterotoxin b-induced lethal shock. J. Biol. Chem. 2011, 286, 9737–9747. [Google Scholar]
- Reichert, J.M.; Dewitz, M.C. Anti-infective monoclonal antibodies: Perils and promise of development. Nat. Rev. Drug. Discov. 2006, 5, 191–195. [Google Scholar] [CrossRef]
- Nelson, A.L.; Dhimolea, E.; Reichert, J.M. Development trends for human monoclonal antibody therapeutics. Nat. Rev. Drug. Discov. 2010, 9, 767–774. [Google Scholar] [CrossRef]
- Mazumdar, S. Raxibacumab. MAbs 2009, 1, 531–538. [Google Scholar] [CrossRef]
- Froude, J.W.; Stiles, B.; Pelat, T.; Thullier, P. Antibodies for biodefense. MAbs 2011, 3, 517–527. [Google Scholar] [CrossRef]
- Salfeld, J.G. Isotype selection in antibody engineering. Nat. Biotechnol. 2007, 25, 1369–1372. [Google Scholar] [CrossRef]
- Abboud, N.; Chow, S.K.; Saylor, C.; Janda, A.; Ravetch, J.V.; Scharff, M.D.; Casadevall, A. A requirement for fcgammar in antibody-mediated bacterial toxin neutralization. J. Exp. Med. 2010, 207, 2395–2405. [Google Scholar] [CrossRef]
- Jefferis, R. Antibody therapeutics: Isotype and glycoform selection. Expert. Opin. Biol. Ther. 2007, 7, 1401–1413. [Google Scholar] [CrossRef]
- Chow, S.K.; Casadevall, A. Evaluation of cryptococcus neoformans galactoxylomannan-protein conjugate as vaccine candidate against murine cryptococcosis. Vaccine 2011, 29, 1891–1898. [Google Scholar] [CrossRef]
- Sun, H.X.; Xie, Y.; Ye, Y.P. Advances in saponin-based adjuvants. Vaccine 2009, 27, 1787–1796. [Google Scholar] [CrossRef]
- Mestas, J.; Hughes, C.C. Of mice and not men: differences between mouse and human immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar]
- Presta, L.G. Molecular engineering and design of therapeutic antibodies. Curr. Opin. Immunol. 2008, 20, 460–470. [Google Scholar] [CrossRef]
- Goodner, K.; Horsfall, F.L. The protective action of type i antipneumococcus serum in mice: I. The quantitative aspects of the mouse protection test. J. Exp. Med. 1935, 62, 359–374. [Google Scholar] [CrossRef]
- Smith, K.; Crowe, S.R.; Garman, L.; Guthridge, C.J.; Muther, J.J.; McKee, E.; Zheng, N.Y.; Farris, A.D.; Guthridge, J.M.; Wilson, P.C.; et al. Human monoclonal antibodies generated following vaccination with ava provide neutralization by blocking furin cleavage but not by preventing oligomerization. Vaccine 2012, 30, 4276–4283. [Google Scholar]
- Maynard, J.A.; Maassen, C.B.; Leppla, S.H.; Brasky, K.; Patterson, J.L.; Iverson, B.L.; Georgiou, G. Protection against anthrax toxin by recombinant antibody fragments correlates with antigen affinity. Nat. Biotechnol. 2002, 20, 597–601. [Google Scholar] [CrossRef]
- Park, Y.S.; Lee, J.H.; Hung, C.F.; Wu, T.C.; Kim, T.W. Enhancement of antibody responses to bacillus anthracis protective antigen domain iv by use of calreticulin as a chimeric molecular adjuvant. Infect. Immun. 2008, 76, 1952–1959. [Google Scholar] [CrossRef]
- Jefferis, R. Glycosylation as a strategy to improve antibody-based therapeutics. Nat. Rev. Drug. Discov. 2009, 8, 226–234. [Google Scholar] [CrossRef]
- Kimura, A.; Mountzouros, K.T.; Schad, P.A.; Cieplak, W.; Cowell, J.L. Pertussis toxin analog with reduced enzymatic and biological activities is a protective immunogen. Infect. Immun. 1990, 58, 3337–3347. [Google Scholar]
- Zhang, X.; Askins, J.; Fleming, R.; Sturm, B.; Poortman, C.; Viriassov, P.; Peterson, B.; Flynn, M.; Miao, Y.; Zukauskas, D.; et al. Selection of Potent Neutralizing Human Monoclonal Antibodies to Protective Antigen of Bacillus Anthracis. In Proceedings of 43rd Interscience Conference on Antimicrobial Agents and Chemotherapy, Washington, DC, USA, 14–17 September 2003; p. 277.
- Moayeri, M.; Leppla, S.H. Cellular and systemic effects of anthrax lethal toxin and edema toxin. Mol. Aspects. Med. 2009, 30, 439–455. [Google Scholar] [CrossRef]
- Nowakowski, A.; Wang, C.; Powers, D.B.; Amersdorfer, P.; Smith, T.J.; Montgomery, V.A.; Sheridan, R.; Blake, R.; Smith, L.A.; Marks, J.D. Potent neutralization of botulinum neurotoxin by recombinant oligoclonal antibody. Proc. Natl. Acad. Sci. USA 2002, 99, 11346–11350. [Google Scholar]
- Volk, W.A.; Bizzini, B.; Snyder, R.M.; Bernhard, E.; Wagner, R.R. Neutralization of tetanus toxin by distinct monoclonal antibodies binding to multiple epitopes on the toxin molecule. Infect. Immun. 1984, 45, 604–609. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Chow, S.-K.; Casadevall, A. Monoclonal Antibodies and Toxins—A Perspective on Function and Isotype. Toxins 2012, 4, 430-454. https://doi.org/10.3390/toxins4060430
AMA Style
Chow S-K, Casadevall A. Monoclonal Antibodies and Toxins—A Perspective on Function and Isotype. Toxins. 2012; 4(6):430-454. https://doi.org/10.3390/toxins4060430
Chicago/Turabian StyleChow, Siu-Kei, and Arturo Casadevall. 2012. "Monoclonal Antibodies and Toxins—A Perspective on Function and Isotype" Toxins 4, no. 6: 430-454. https://doi.org/10.3390/toxins4060430