Differences in the Regulation of Ochratoxin A by the HOG Pathway in Penicillium and Aspergillus in Response to High Osmolar Environments

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
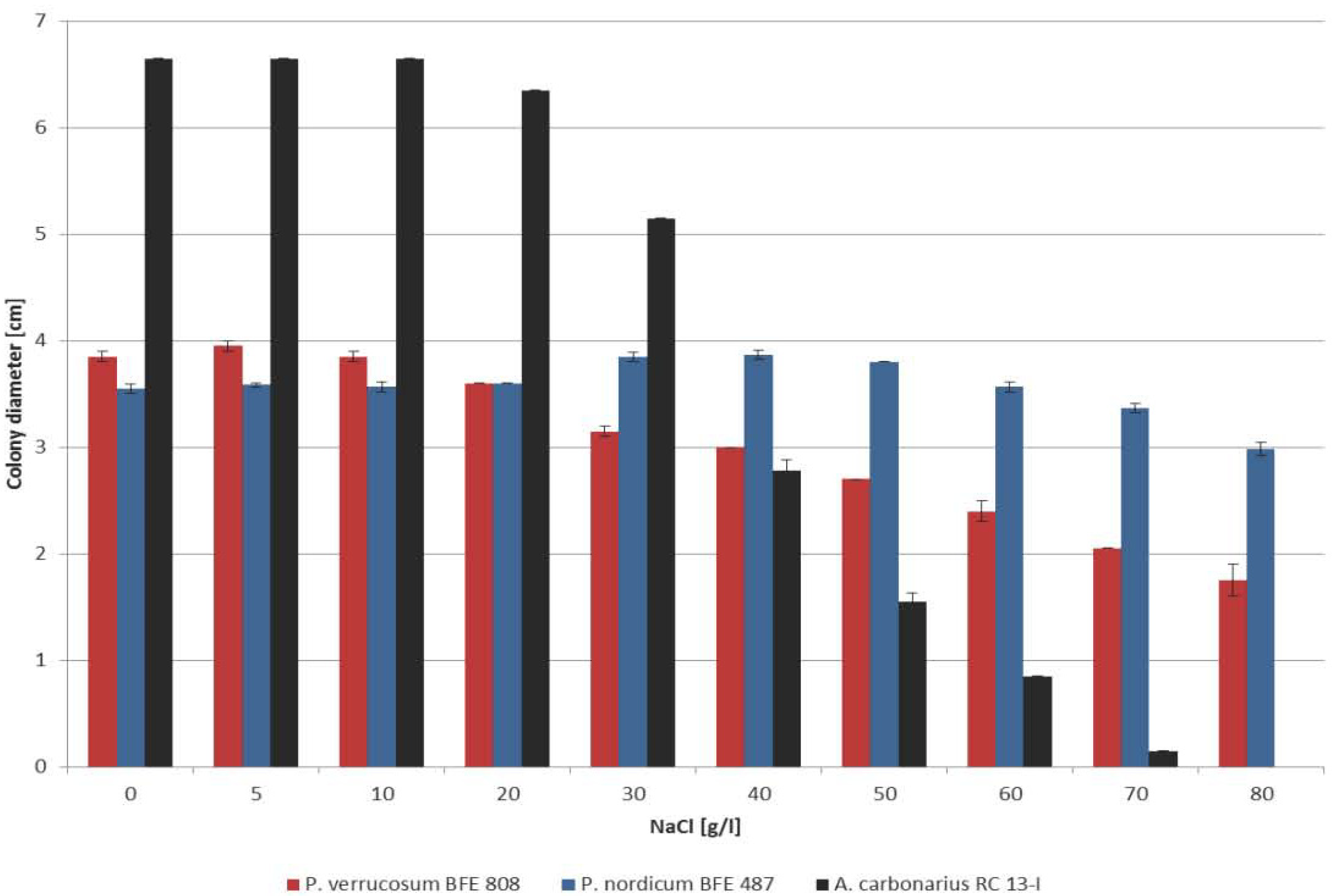
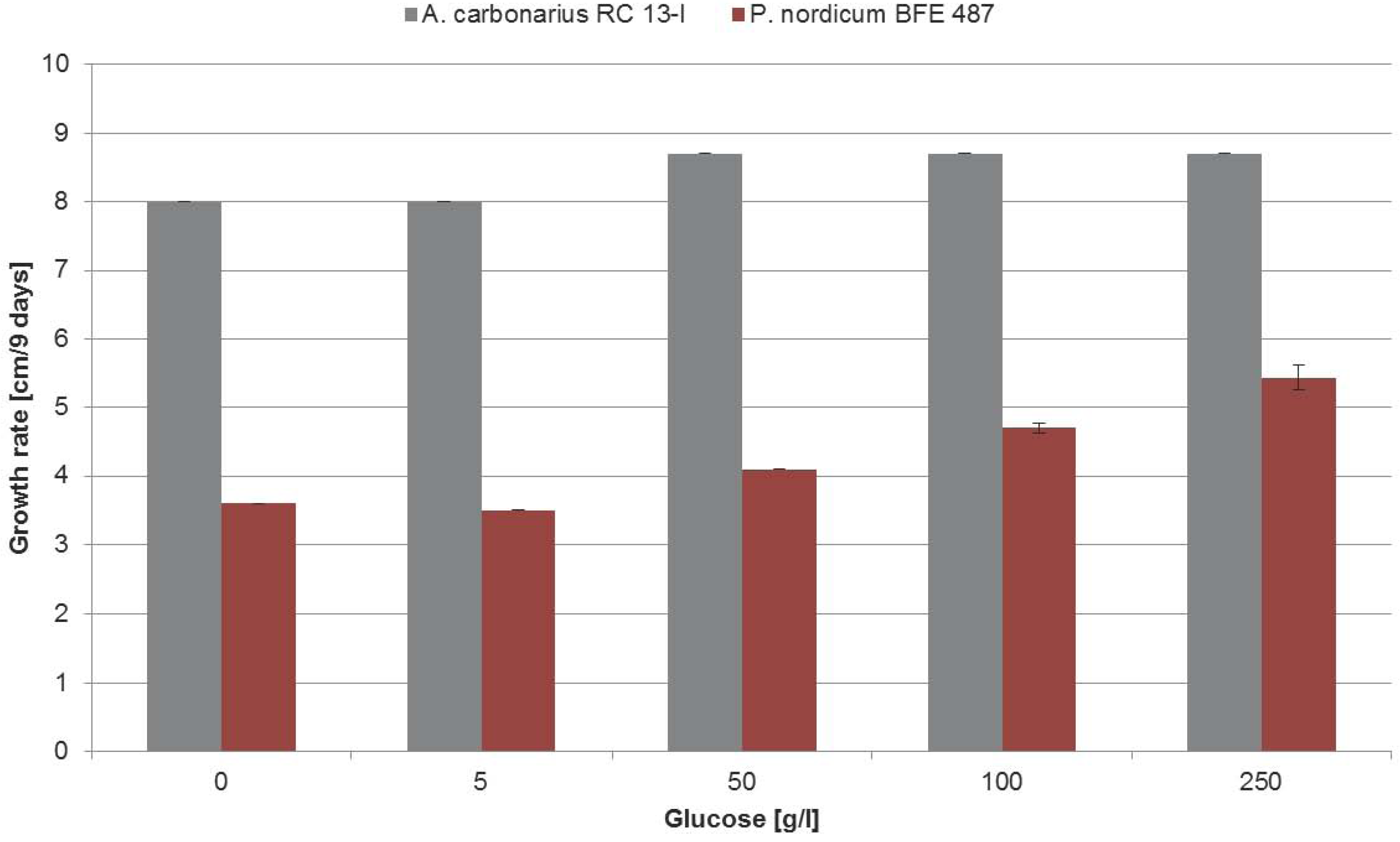
2.1. Growth Behavior of P. nordicum, P. verrucosum and A. carbonarius on YES Medium with Increasing Concentrations of NaCl

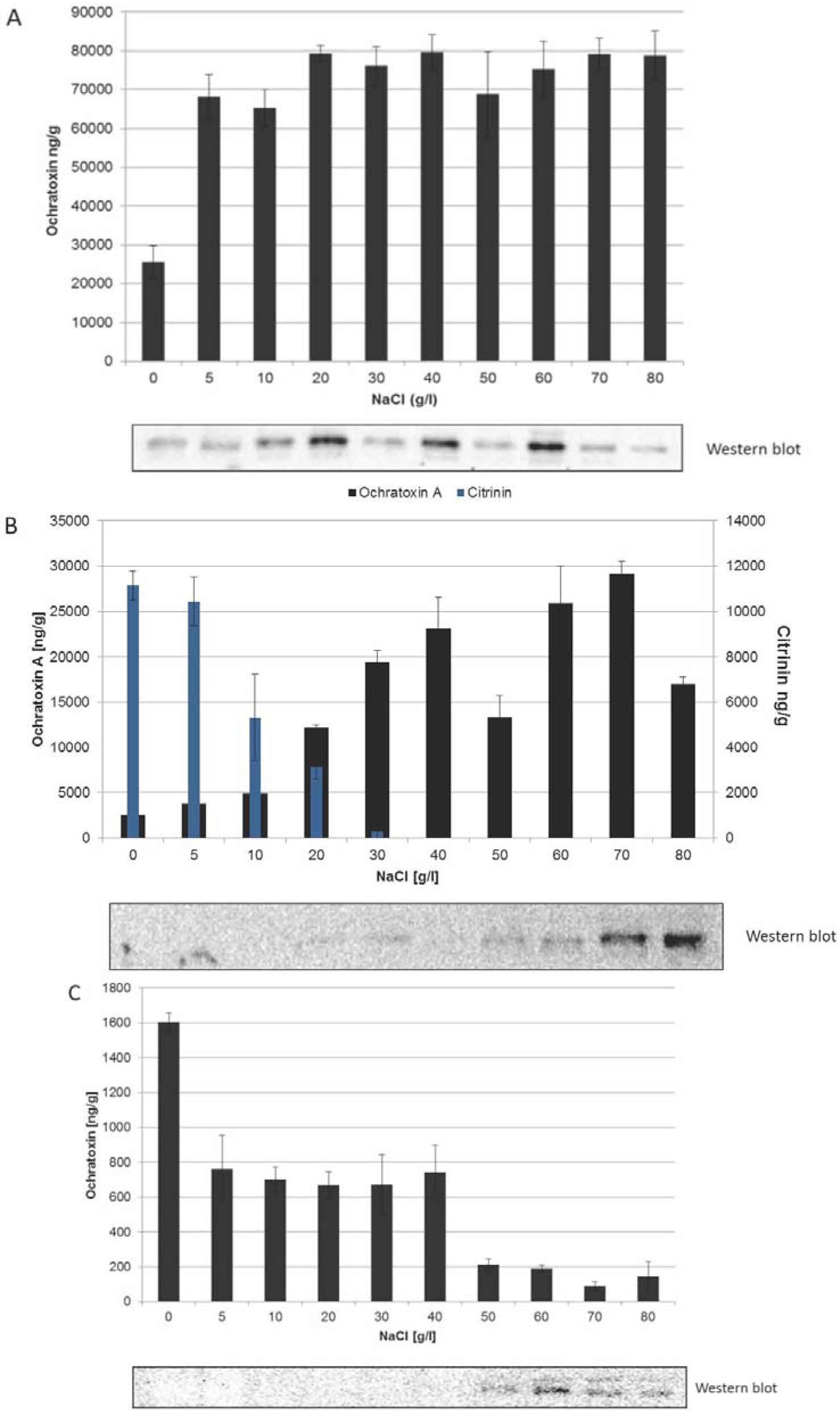
2.2. Different Correlations between HOG Phosphorylation and the Onset of Ochratoxin A Biosynthesis in the Three Species
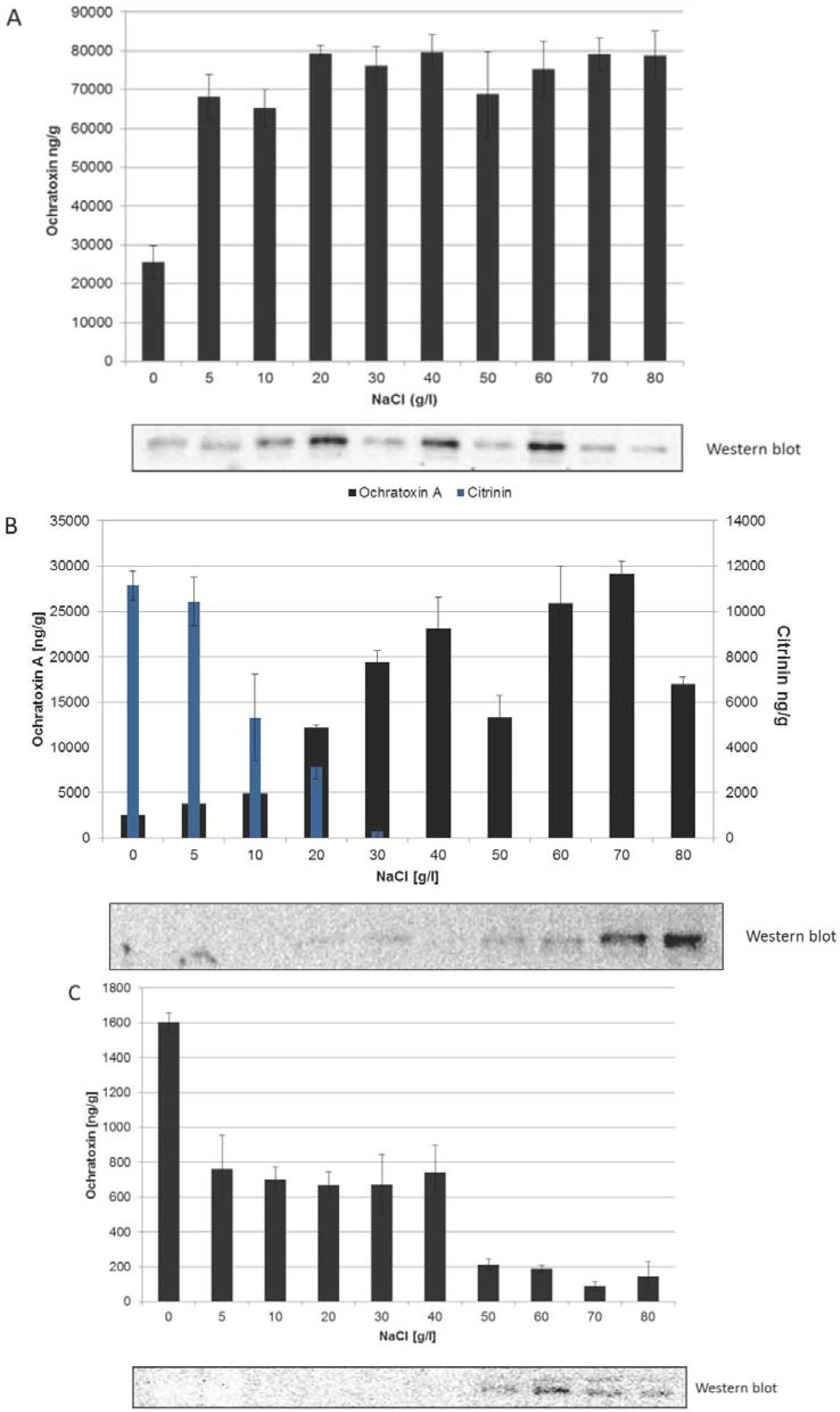
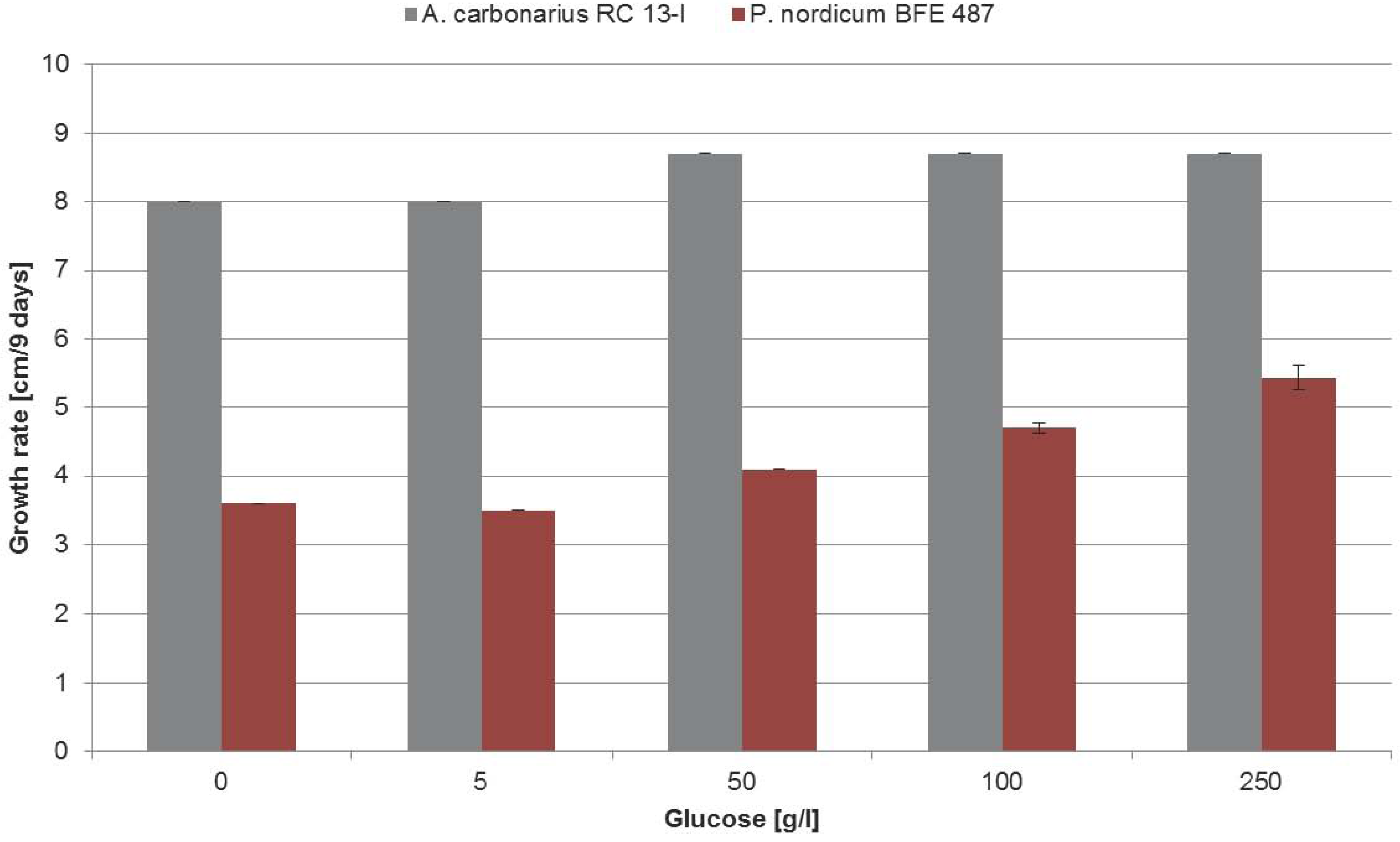
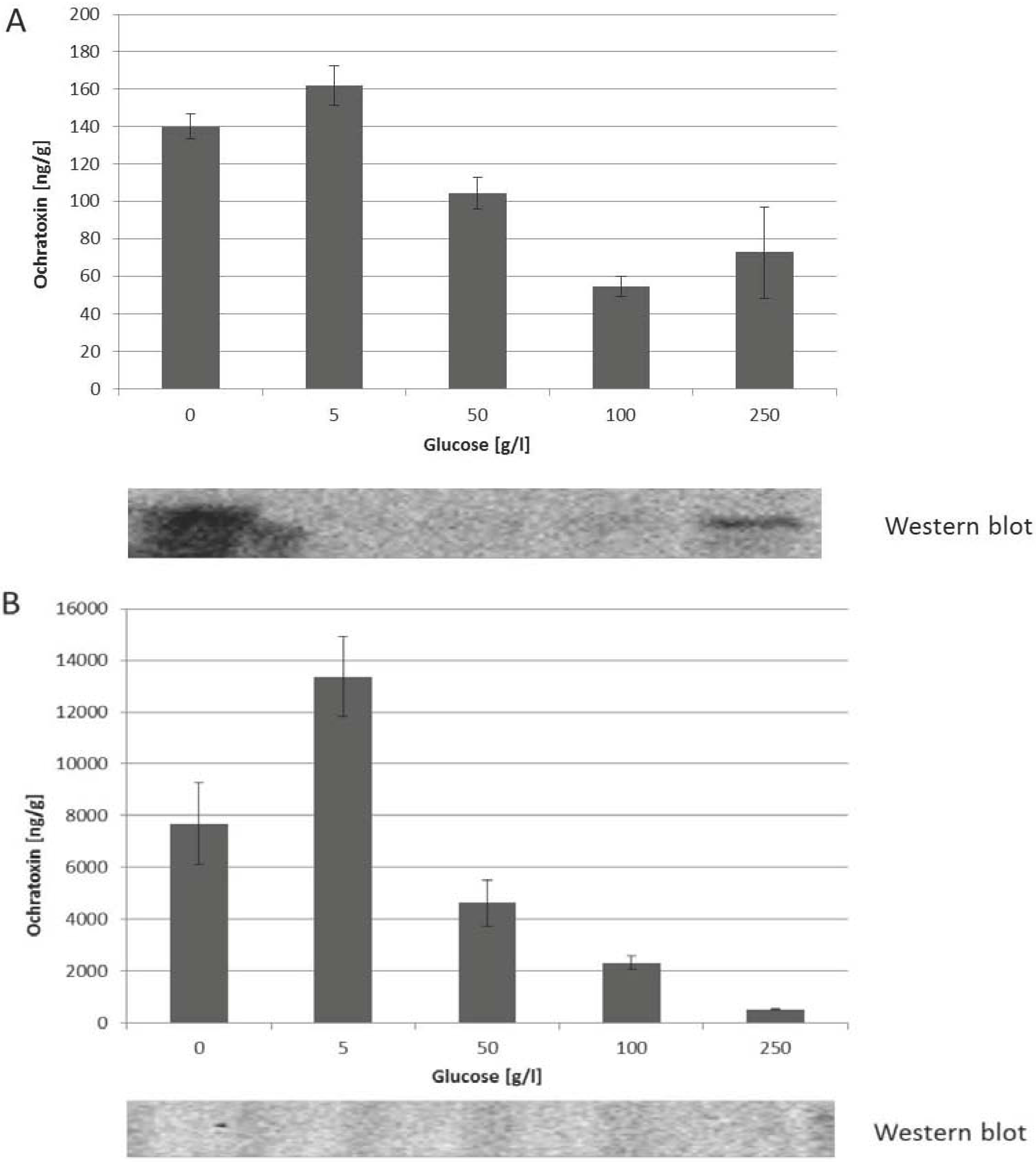
2.3. Influence of Increasing Amounts of Glucose on Growth, Ochratoxin A Biosynthesis and HOG Phosphorylation by P. nordicum and A. carbonarius

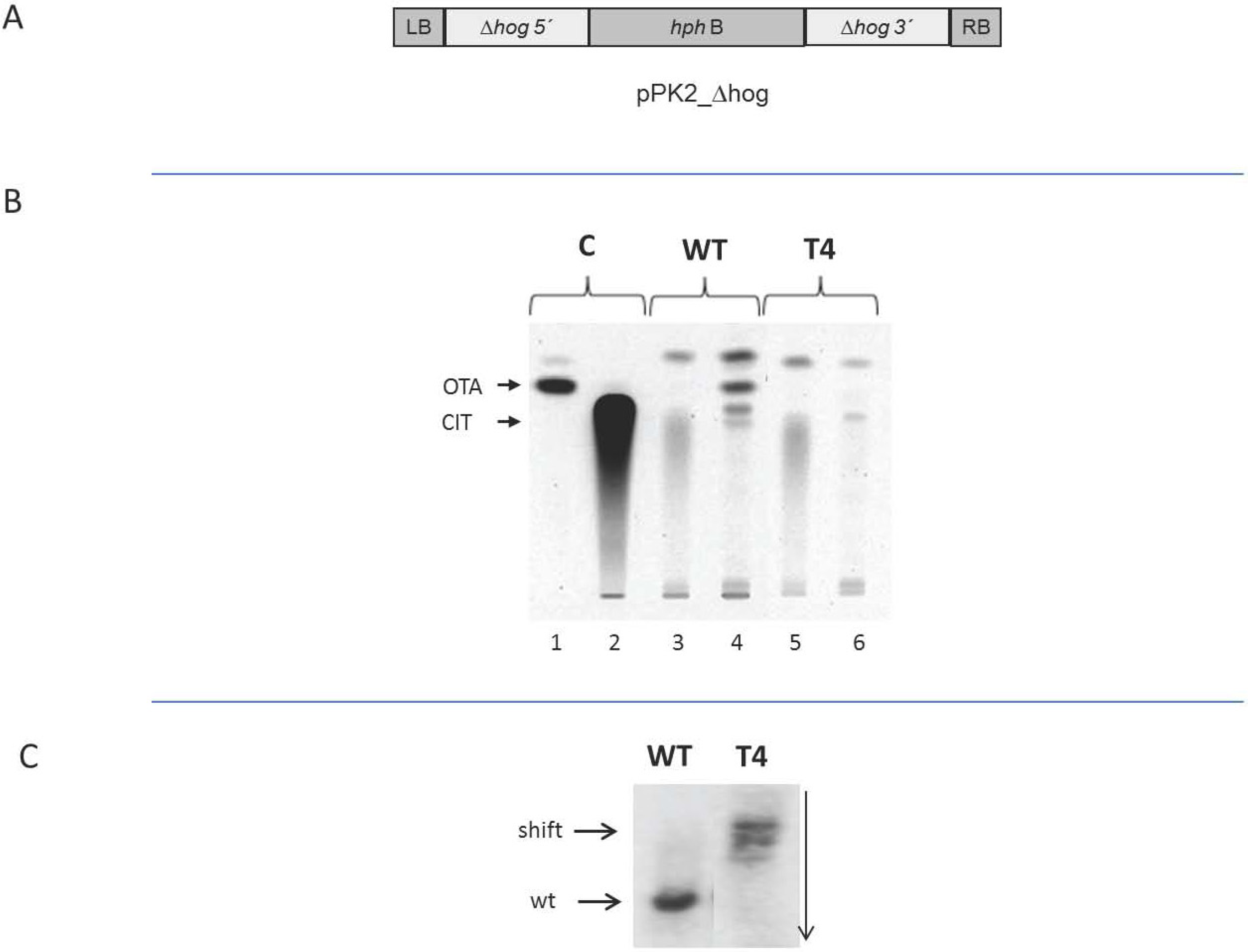
2.4. Dependence of the Biosynthesis of Ochratoxin A on Functional HOG
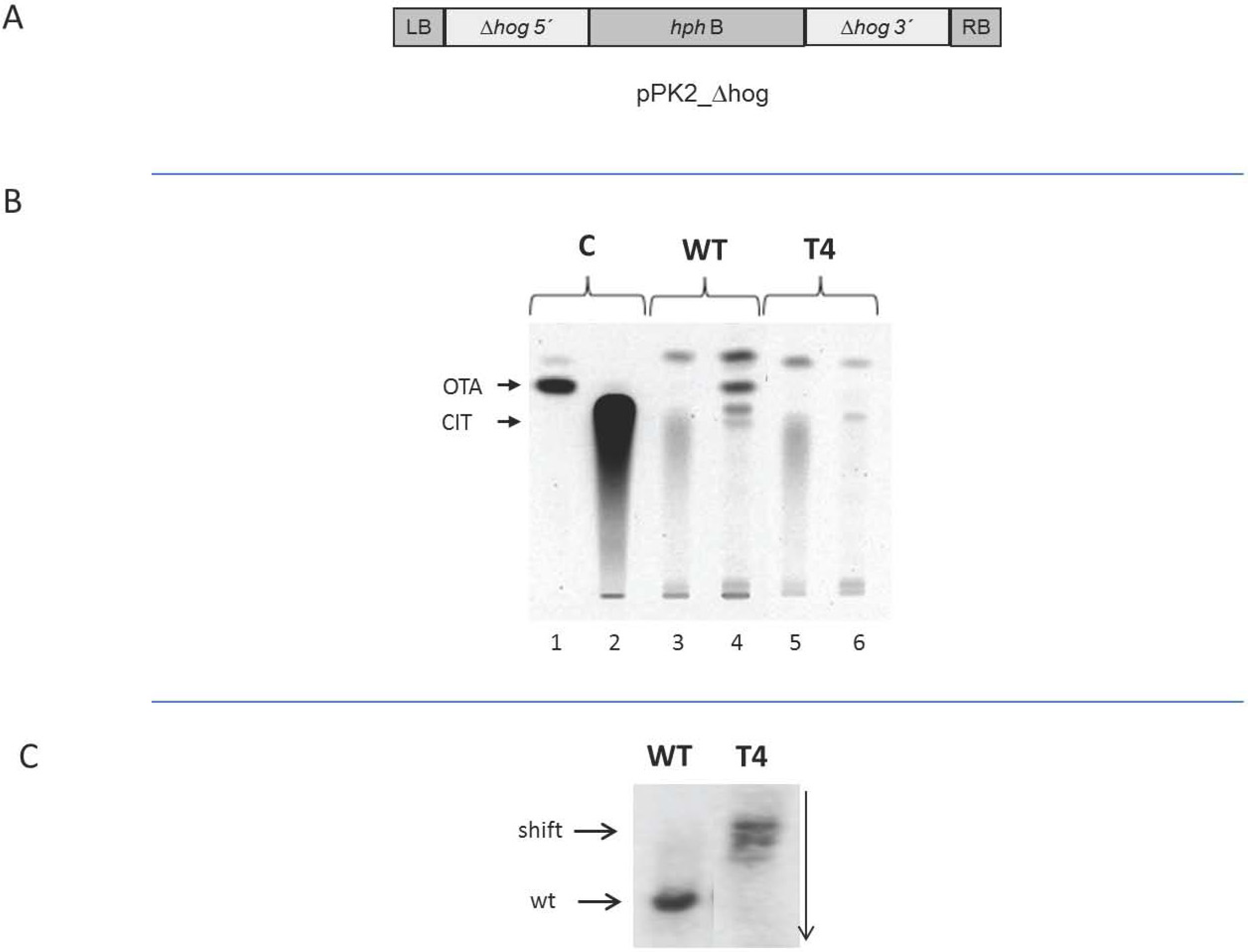
3. Discussion
4. Experimental Section
4.1. Strains and Growth Conditions
4.2. Growth Assessment
4.3. Determination of Ochratoxin A and Citrinin by Thin Layer (TLC) and High Pressure Liquid (HPLC) Chromatography
4.4. Generation and Treatment of Data
4.5. Protein Extraction
4.6. SDS PAGE and Western Blot
4.7. A. tumefaciens Mediated Transformation (ATMT) to Inactivate the hog Gene
4.8. Southern Blotting
5. Conclusions
Acknowledgments
Abbreviations
| YES | yeast extract sucrose medium |
| MEA | malt extract agar |
| HOG signal pathway | high osmolarity glycerol signal pathway |
| MAPK | mitogen activated protein kinase |
| ATMT | Agrobacterium tumefaciens mediated transformation |
| TLC | thin layer chromatography |
Conflict of Interest
References
- Pardo, E.; Marin, S.; Ramos, A.J.; Sanchis, V. Occurrence of ochratoxingenic fungi and ochratoxin A in green coffee from different origins. Food Sci. Technol. 2004, 10, 45–49. [Google Scholar] [CrossRef]
- Palumbo, J.D.; O’Keefe, T.L.; Vasquez, S.J.; Mahoney, N.E. Isolation and identification of ochratoxin A producing Aspergillus section Nigri strains from Californian raisins. Lett. Appl. Microbiol. 2011, 52, 330–336. [Google Scholar] [CrossRef]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage; Aspen Publishers: Gaithersburg, MD, USA, 1999. [Google Scholar]
- Leong, S.I.L.; Hocking, A.D.; Scott, E.S. Effect of temperature and water activity on growth and ochratoxin A production by Australian Aspergillus carbonarius and A. niger isolates on a simulated grape juice medium. Int. J. Food Microbiol. 2006, 110, 209–216. [Google Scholar] [CrossRef]
- Battilani, P.; Giorni, P.; Pietri, A. Epidemiology of toxin-producing fungi and ochratoxin A occurrence in grape. Eur. J. Plant Pathol. 2003, 109, 715–722. [Google Scholar] [CrossRef]
- Perrone, G.; Mulè, G.; Susca, A.; Battilani, P.; Pietri, A.; Logrieco, A. Ochratoxin A Production and amplified fragment length polymorphism analysis of Aspergillus carbonarius, Aspergillus tubingensis, and Aspergillus niger strains Isolated from grapes in Italy. Appl. Environ. Microbiol. 2006, 72, 680–685. [Google Scholar] [CrossRef]
- Lund, F.; Frisvad, J.C. Penicillium verrucosum in wheat and barley indicates presence of ochratoxin A. J. Appl. Microbiol. 2003, 95, 1117–1123. [Google Scholar] [CrossRef]
- Larsen, T.O.; Svendsen, A.; Smedsgaard, J. Biochemical characterization of ochratoxin A-producing strains of the genus Penicillium. Appl. Environ. Microbiol. 2001, 67, 3630–3635. [Google Scholar] [CrossRef]
- Sonjak, S.; Licen, M.; Frisvad, J.C.; Gunde-Cimerman, N. Salting of dry-cured meat—A potential cause of contamintation with the ochratoxin A-producing species Penicillium nordicum. Food Microbiol. 2011, 28, 1111–1116. [Google Scholar] [CrossRef]
- Butinar, L.; Frisvad, J.C.; Gunde-Cimerman, N. Hypersaline waters—A potential source of foodborne toxigenic Aspergilli and Penicillia. FEMS Microbiol. Ecol. 2011, 77, 186–199. [Google Scholar] [CrossRef]
- Peintner, U.; Geiger, J.; Pöder, R. The mycobiota of speck, a traditional tyrolean smoked and cured ham. J. Food Protect. 2000, 63, 1399–1403. [Google Scholar]
- Heperkan, D.; Dazkir, G.S.; Kansu, D.Z.; Güler, F.K. Influence of temperature on citrinin accumulation by Penicillium citrinum and Penicillium verrucosum in black table olives. Toxin Rev. 2009, 28, 180–186. [Google Scholar] [CrossRef]
- Tokusoglu, Ö; Bozoglu, F. Citrinin risk in black and green table olives: Simultaneous determination with ochratoxin-A by optimized extration and IAC-HPLC-FD. Ital. J. Food Sci. 2010, 22, 284–291. [Google Scholar]
- Schmidt-Heydt, M.; Rüfer, C.E.; Raupp, F.; Bruchmann, A.; Perrone, G.; Geisen, R. Influence of light on food relevant fungi with emphasis on ochratoxin producing species. Int. J. Food Microbiol. 2011, 145, 229–237. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Graf, E.; Stoll, D.; Geisen, R. The biosynthesis of ochratoxin A by Penicillium as one mechanism for adaptation to NaCl rich foods. Food Microbiol. 2012, 29, 233–241. [Google Scholar] [CrossRef]
- Ochiai, N.; Tokai, T.; Nishiuchi, T.; Takahashi-Ando, N.; Fujimura, M.; Kimura, M. Involvement of the osmosensor histidine kinase and osmotic stress-activated protein kinases in the regulation of secondary metabolism in Fusarium graminearum. Biochem. Biophys. Res. Commun. 2007, 363, 639–644. [Google Scholar] [CrossRef]
- Graf, E.; Schmidt-Heydt, M.; Geisen, R. HOG MAP kinase regulation of alternariol biosynthesis in Alternaria alternata is important for substrate colonization. Int. J. Food Microbiol. 2012, 157, 353–359. [Google Scholar] [CrossRef]
- Covert, S.F.; Kapoor, P.; Lee, M.-H.; Briley, A.; Nairn, C.J. Agrobacterium tumefaciens-mediated transformation of Fusarium circinatum. Mycol. Res. 2001, 105, 259–264. [Google Scholar] [CrossRef]
- Bragulat, M.R.; Abarca, M.L.; Cabanes, J. Low occurrence of patulin- and citrinin-producing species isolated from grapes. Lett. Appl. Microbiol. 2008, 47, 286–289. [Google Scholar] [CrossRef]
- Stormer, F.C.; Sandven, P.; Huitfeldt, S.; Eduard, W.; Skogstad, A. Does the mycotoxin citrinin function as a sun protectant in conidia from Penicillium verrucosum? Mycopathologia 1998, 142, 43–47. [Google Scholar]
- Heider, E.M.; Harper, J.K.; Grant, D.M.; Hoffmann, A.; Dugan, F.; Tomer, D.P.; O’Neil, K.O. Exploring unusual antioxidant activity in a benzoic acid derivative: A proposed mechanism for citrinin. Tetrahedron 2012, 62, 1199–1208. [Google Scholar]
- Meister, U. New method of citrinin determination by HPLC after polyamide column clean-up. Eur. Food Res. Technol. 2004, 218, 394–399. [Google Scholar] [CrossRef]
- Molinié, A.; Faucet, V.; Castegnaro, M.; Pfohl-Leszkowicz, A. Pfohl-Leszkowicz, A. Analysis of some breakfeast cereals on the French market for their contents of ochatoxin A, citrinin and fumonisin B1: Development of a method for simultaneous extraction of ochratoxin A and citrinin. Food Chem. 2005, 92, 391–400. [Google Scholar]
- Samapundo, S.; Deschuyffeleer, N.; van Laere, D.; de Leyn, I.; Devlieghere, F. Effect of NaCl reduction and replacement on the growth of fungi important to the spoilage of bread. Food Microbiol. 2010, 27, 749–756. [Google Scholar] [CrossRef]
- Ayodele, S.M.; Ojoghoro, O.J. Salt tress effects on the vegetative growth of Pleurotus tuberregium (FR) sing. J. Biol. Sci. 2007, 7, 1278–1281. [Google Scholar] [CrossRef]
- Rispail, N.; Soanes, D.M.; Ant, C.; Czajkowski, R.; Grünler, A.; Huguet, R.; Perez-Nadales, E.; Poli, A.; Sartorel, E.; Valiante, V.; et al. Comparative genomics of MAP kinase and calcium-calcineurin signalling compontents in plant and human pathogenic fungi. Fungal Genet. Biol. 2009, 46, 287–298. [Google Scholar] [CrossRef]
- Kohut, G.; Ádám, A.L.; Fazekas, B.; Hornok, L. N-starvation stress induced FUM gene expression and fumonisin production is mediated via the HOG-type MAPK pathway in Fusarium proliferatum. Int. J. Food Microbiol. 2009, 130, 65–69. [Google Scholar] [CrossRef]
- Li, L.; Shao, Y.; Li, Q.; Yang, S.; Chen, F. Identification of Mga1, a G-protein alpha-subunit gene involved in regulating citrinin and pigment production in Monascus ruber M7. FEMS Microbiol. Lett. 2010, 308, 108–114. [Google Scholar]
- Gunde-Cimerman, N.; Sonjak, S.; Zalar, P.; Frisvad, J.C.; Diderichsen, B.; Plemenitas, A. Extremophilic fungi in arctic ice: A relationship between adaptation to low temperature and water activity. Phys. Chem. Earth 2003, 28, 1273–1278. [Google Scholar] [CrossRef]
- Jiménez-Martín, E.; Zuzuarregui, A.; Gomar-Alba, M.; Guteérrez, D.; Gil, C.; del Olmo, M. Molecular response of Saccharomyces cerevisiae wine and laboratory strains to high sugar stress conditions. Int. J. Food Microbiol. 2011, 145, 211–220. [Google Scholar] [CrossRef]
- Sambrook, J.; Russel, D.W. Molecular Cloning, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbour, NY, USA, 2001. [Google Scholar]
- Southern, E. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 1975, 98, 503–517. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stoll, D.; Schmidt-Heydt, M.; Geisen, R. Differences in the Regulation of Ochratoxin A by the HOG Pathway in Penicillium and Aspergillus in Response to High Osmolar Environments. Toxins 2013, 5, 1282-1298. https://doi.org/10.3390/toxins5071282
Stoll D, Schmidt-Heydt M, Geisen R. Differences in the Regulation of Ochratoxin A by the HOG Pathway in Penicillium and Aspergillus in Response to High Osmolar Environments. Toxins. 2013; 5(7):1282-1298. https://doi.org/10.3390/toxins5071282
Chicago/Turabian StyleStoll, Dominic, Markus Schmidt-Heydt, and Rolf Geisen. 2013. "Differences in the Regulation of Ochratoxin A by the HOG Pathway in Penicillium and Aspergillus in Response to High Osmolar Environments" Toxins 5, no. 7: 1282-1298. https://doi.org/10.3390/toxins5071282