Deoxynivalenol and Oxidative Stress Indicators in Winter Wheat Inoculated with Fusarium graminearum

Abstract
:1. Introduction
2. Results
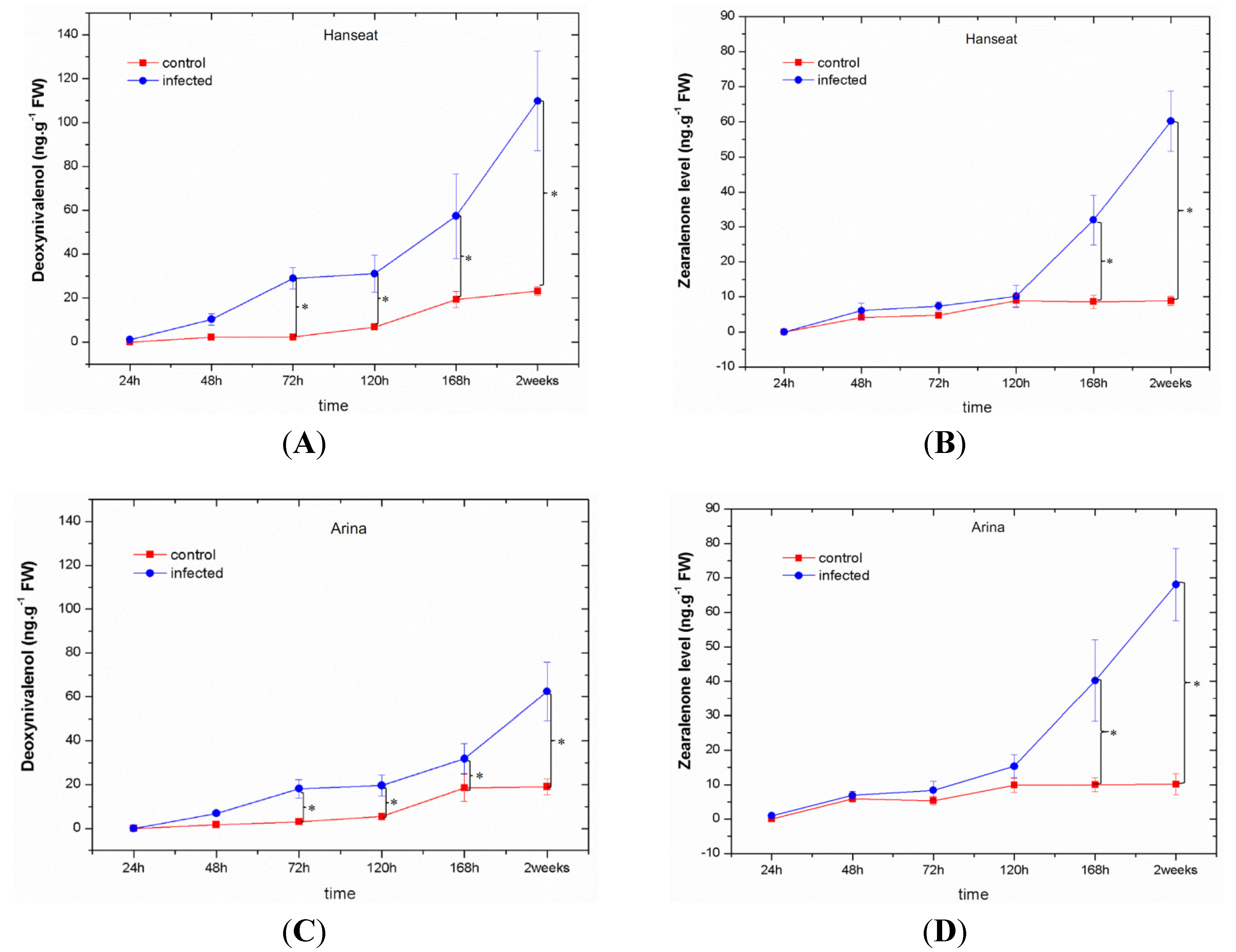
2.1. Mycotoxin Contents

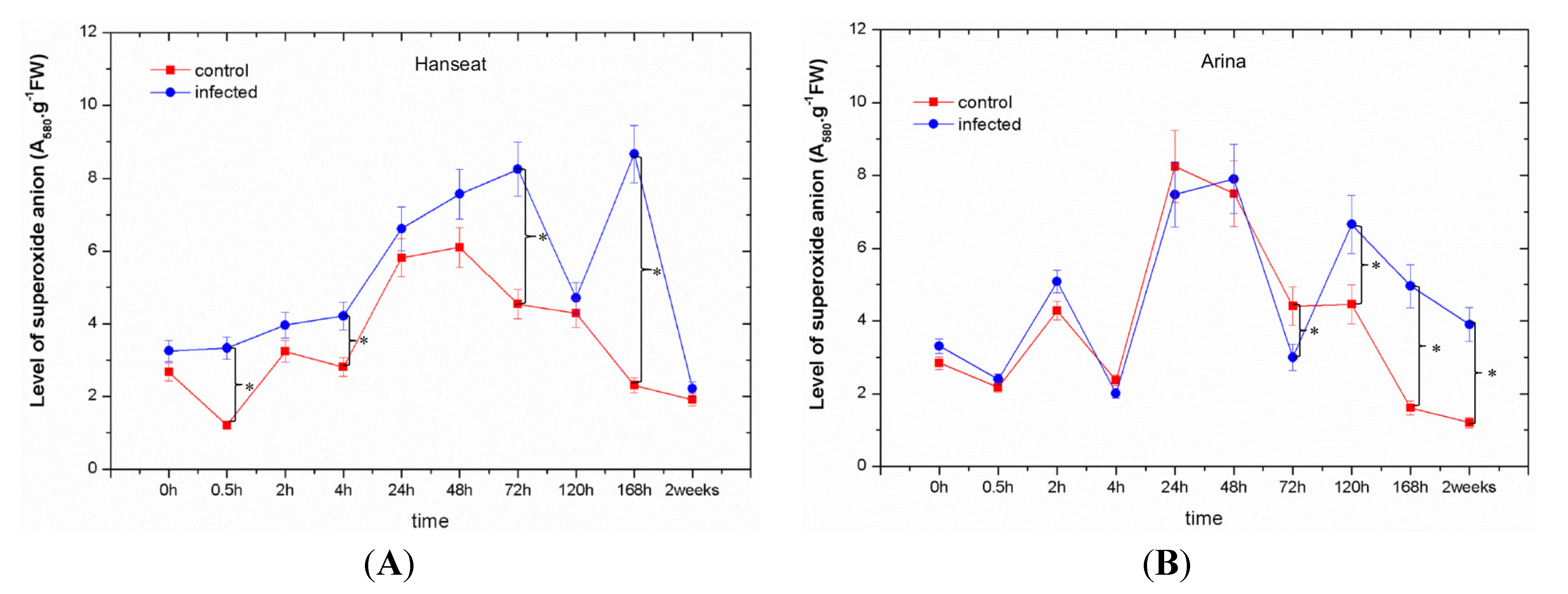
2.2. Generation of Superoxide Anion

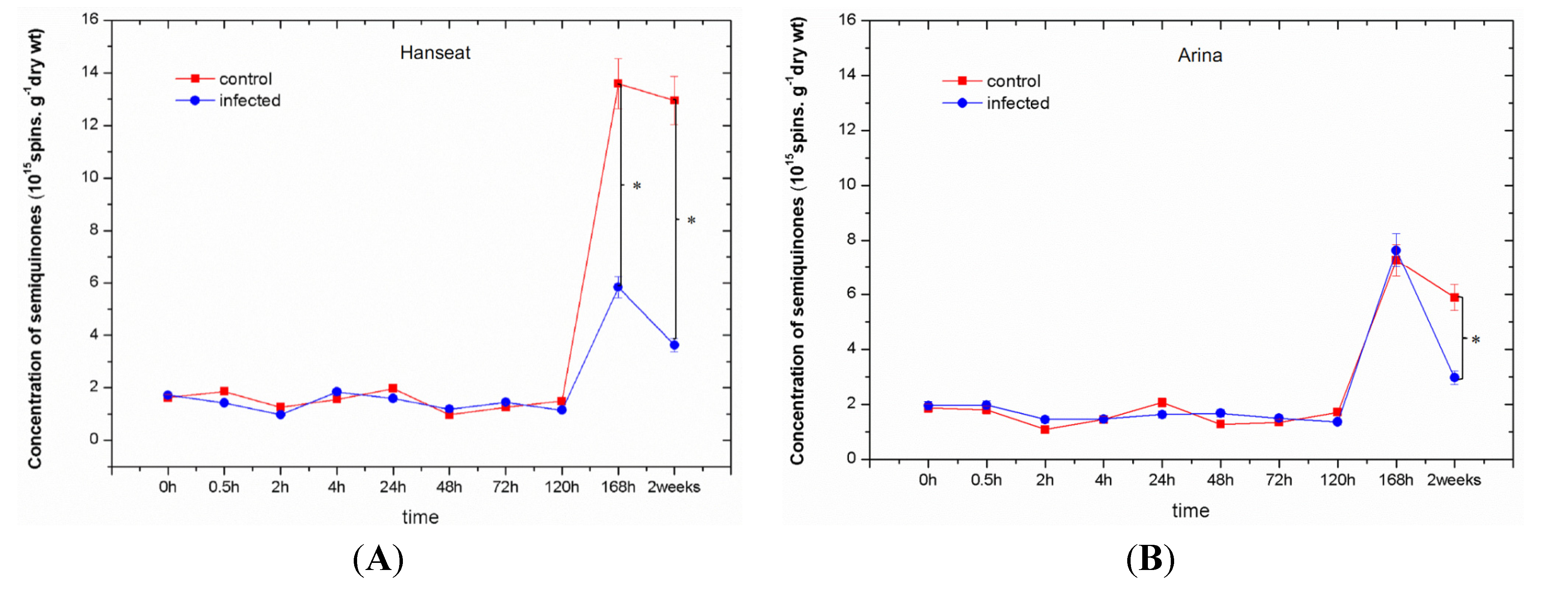
2.3. Generation of Semiquinone Radicals

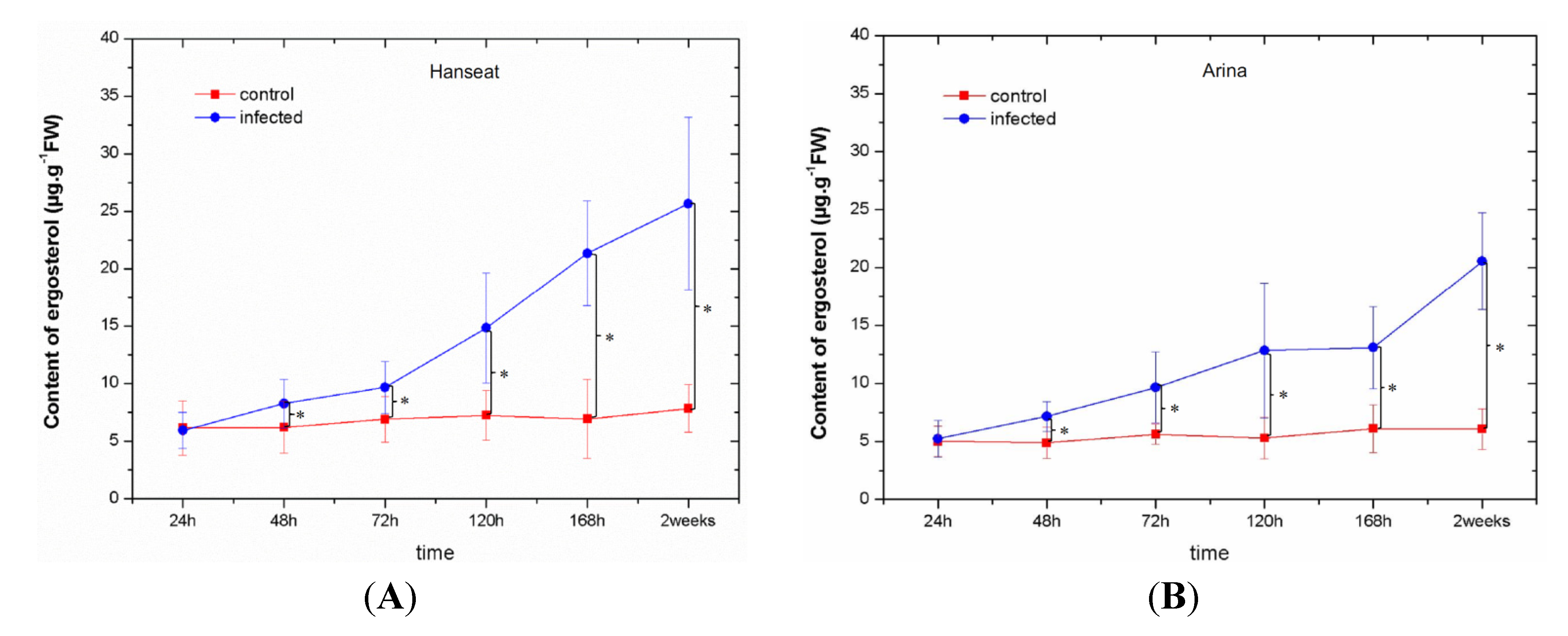
2.4. Analysis of Disease Symptoms and the Level of Ergosterol

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time after inoculation | Disease development | |||
|---|---|---|---|---|
| Arina - Resistant | Hanseat - Susceptible | |||
| Control | Infected | Control | Infected | |
| 24 h | − | − | − | − |
| 48 h | − | − | − | + |
| 72 h | − | ++ | − | ++ |
| 120 h | − | ++ | − | ++++ |
| 168 h | − | +++ | − | ++++ |
| 2 weeks | − | ++++ | − | +++++ |

3. Discussion
4. Experimental Section
4.1. Plant Material and Growth Conditions
4.2. Fusarium Strain and Inoculum Preparation
4.3. Standards, Chemicals, and Reagents
4.4. Extraction and Purification Procedure for Mycotoxins
4.5. HPLC Analysis of Mycotoxins
- Waters 2996 Photodiode Array Detector with a Nova Pak C-18 column (300 mm × 3.9 mm) for DON (λmax = 224 nm) analysis,
- Waters 2475 Multi λ Fluorescence Detector (λex = 274 nm, λem = 440 nm) and a Waters 2996 Photodiode Array Detector with a Nova Pak C-18 column (150 mm ×3.9 mm) for ZON analysis.
4.6. Ergosterol Extraction, Purification, and HPLC Analysis
4.7. Determination of Superoxide Anion Radical Content
4.8. Determination of Semiquinone Radicals
4.9. Statistical Analysis
5. Conclusions
- The susceptible cultivar was characterized by a much greater accumulation of deoxynivalenol than the resistant cultivar.
- In the susceptible cultivar a marked post-infection increase in O2•− level was found up to 120 h after inoculation.
- The level of O2•− generation in infested ears of both wheat cultivars was generally greater than in the control.
- An earlier reduction in the level of O2•− generation with the time after inoculation was observed in the resistant rather than in the susceptible cultivar.
- The concentration of semiquinone radicals, detected by EPR, increased at later culture times; however, in infested ears of wheat it was generally lower than in the control.
- The resistant cultivar was characterized by a lower level of semiquinone radicals than the sensitive cultivar especially at the late phase after inoculation. It may probably indicate their incorporation into polymers, such as lignins, and strengthening of the cell wall.
- Development of disease was inhibited in the resistant cultivar and ergosterol content was lower than in the sensitive cultivar.
- Production of the mycotoxin DON and the level of generation of superoxide anion (O2•−) in the resistant cultivar at the late phase after inoculation was lower than in the sensitive cultivar.
Acknowledgments
Conflicts of Interest
References
- Molodchenkova, O.O.; Adamovskaya, V.G.; Yu, A.L.; Gontarenko, O.V.; Sokolov, V.M. Maize response to salicylic acid and Fusarium moniliforme. Appl. Biochem. Microbiol. 2002, 38, 381–385. [Google Scholar] [CrossRef]
- Thatcher, L.F.; Manners, J.M.; Kazan, K. Fusarium oxysporum hijacks COI1-mediated jasmonate signaling to promote disease development in Arabidopsis. Plant J. 2009, 588, 927–939. [Google Scholar] [CrossRef]
- Berger, S.; Benediktyova, Z.; Matous, K.; Bonfig, K.B.; Mueller, M.J.; Nedbal, L.; Roitsch, T. Visualiztion of dynamics of plant–pathogen interaction by novel combination of chlorophyll fluorescence imaging and statistical analysis: Differential effects of virulent and avirulent strains of P. syringae and of oxylipins on A. thaliana. J. Exp. Bot. 2007, 58, 797–806. [Google Scholar]
- Wojtaszek, P. Mechanisms for the generation of reactive oxygen species in plant defence response. Acta Physiol. Plant 1997, 19, 581–589. [Google Scholar] [CrossRef]
- Hückelhoven, R.; Kogel, K.H. Reactive oxygen intermediates in plant–microbe interactions: Who is who in powdery mildew resistance? Planta 2003, 216, 891–902. [Google Scholar]
- Morkunas, I.; Bednarski, W. Fusarium oxysporum—Induced oxidative stress and antioxidative defenses of yellow lupine embryo axes with different sugar levels. J. Plant Physiol. 2008, 165, 262–277. [Google Scholar] [CrossRef]
- Doke, N.; Miura, Y.; Sanchez, L.M.; Park, H.-J.; Noritake, T.; Yoshioka, H.; Kawakita, K. The oxidative burst protects plants against pathogen attack: Mechanism and role as an emergency signal for plant bio-defense-a review. Gene 1996, 179, 45–51. [Google Scholar] [CrossRef]
- Govrin, E.M.; Levine, A. The hypersensitive response facilitates plant infection by the necrotrophic pathogen Botrytis cinerea. Curr. Biol. 2000, 10, 751–757. [Google Scholar] [CrossRef]
- Apell, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Hammerschmidt, R. Phenols and plant–pathogen interactions: The saga continues. Physiol. Mol. Plant Pathol. 2005, 66, 77–78. [Google Scholar] [CrossRef]
- Morkunas, I.; Bednarski, W.; Kopyra, M. Defense strategies of pea embryo axes with different levels of sucrose to Fusarium oxysporum and Ascochyta pisi. Physiol. Mol. Plant Pathol. 2008, 72, 167–178. [Google Scholar] [CrossRef]
- Kotchoni, S.O.; Gachomo, E.W. The reactive oxygen species network pathways: An essential prerequisite for perception of pathogen attack and the acquired disease resistance in plants. J. Biosci. 2006, 31, 389–404. [Google Scholar] [CrossRef]
- Mayer, A.M.; Staples, R.C.; Gil-ad, N.L. Mechanisms of survival necrotrophic fungal plant pathogens in hosts expressing the hypersensitive response. Phytochemistry 2001, 58, 33–41. [Google Scholar] [CrossRef]
- Choquer, M.; Fournier, E.; Kunz, C.; Levis, C.; Pradier, J.-M.; Simon, A.; Viaud, M. Botrytis cinerea virulence factors: New insights into a necrotrophic and polyphageous pathogen. FEMS Microbiol. Lett. 2007, 277, 1–10. [Google Scholar] [CrossRef]
- Reverberi, M.; Zjalic, S.; Ricelli, A.; Punelli, F.; Camera, E.; Fabbri, C.; Picardo, M.; Fanelli, C.; Fabbri, A. Modulation of antioxidant defense in Aspergillus parasiticus is involved in aflatoxin biosynthesis: A role for the ApyapA gene. Eukaryot. Cell 2008, 7, 988–1000. [Google Scholar] [CrossRef]
- Boutigny, A.L.; Richard-Forget, F.; Barreau, C. Natural mechanisms for cereal resistance to Fusarium mycotoxins accumulation. Rev. Eur. J. Plant Pathol. 2008, 121, 411–423. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef]
- Ferrer, E.; Juan-Gracia, A.; Font, G.; Ruiz, M.J. Reactive oxygen species induced by beauvericin, patulin and zearalenone in CHO-K1 cells. Toxicol. In Vitro 2009, 23, 1504–1509. [Google Scholar]
- Wiśniewska, H.; Stępień, Ł.; Waśkiewicz, A.; Beszterda, M.; Góral, T.; Belter, J. Toxigenic Fusarium species infecting wheat heads in Poland. Cent. Eur. J. Biol. 2014, 9, 163–172. [Google Scholar] [CrossRef]
- Goliński, P.; Waśkiewicz, A.; Wiśniewska, H.; Kiecana, I.; Mielniczuk, E.; Gromadzka, K.; Kostecki, M.; Bocianowski, J.; Rymaniak, E. Reaction of winter wheat (Triticum aestivum L.) cultivars to infection with Fusarium spp.: Mycotoxin contamination in grain and chaff. Food Addit. Contam. 2010, 27, 1015–1024. [Google Scholar] [CrossRef]
- Audenaert, K.; van Broeck, R.; Bekaert, B.; de Witte, F.; Heremans, B.; Messens, K.; Höfte, M.; Haesaert, G. Fusarium head blight (FHB) in Flanders: Population diversity, inter-species associations and DON contamination in commercial winter wheat varieties. Eur. J. Plant Pathol. 2009, 125, 445–458. [Google Scholar] [CrossRef]
- Spanic, V.; Lemmens, M.; Drezner, G. Morphological and molecular identification of Fusarium species associated with head blight on wheat in East Croatia. Eur. J. Plant Pathol. 2010, 128, 511–516. [Google Scholar] [CrossRef]
- Jestoi, M.; Paavanen-Huhtala, S.; Parikka, S.; Yli-Mattila, T. In vitro and in vivo mycotoxin production of Fusarium species isolated from Finnish grains. Arch. Phytopathol. Plant Protect. 2008, 41, 545–558. [Google Scholar] [CrossRef]
- Mishra, S.; Ansari, K.M.; Dwivedi, P.D.; Pandey, H.P.; Das, M. Occurrence of deoxynivalenol in cereals and exposure risk assessment in Indian population. Food Control 2013, 30, 549–555. [Google Scholar] [CrossRef]
- Wegulo, S.N. Factors influencing deoxynivalenol accumulation in small grain cereals. Toxins 2012, 4, 1157–1180. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Molto, J.C.; Manes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Brown, N.A.; Antoniw, J.; Hammond-Kosack, K.E. The predicted secretome of the plant pathogenic fungus Fusarium graminearum: A refined comparative analysis. PLoS ONE 2012, 7, e33731. [Google Scholar]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P.; Desjardins, A.E. Tri6 encodes an unusual zinc finger protein involved in regulation of trichothecene biosynthesis in Fusarium sporotrichioides. Appl. Environ. Microbiol. 1995, 61, 1923–1930. [Google Scholar]
- Cuzick, A.; Urban, M.; Hammond-Kosack, K. Fusarium graminearum gene deletion mutants map1 and tri5 reveal similarities and differences in the pathogenicity requirements to cause disease on Arabidopsis and wheat floral tissue. New Phytol. 2008, 177, 990–1000. [Google Scholar] [CrossRef]
- Kimura, M.; Kaneko, I.; Komiyama, M.; Takatsuki, A.; Koshino, H.; Yoneyama, K.; Yamaguchi, I. Trichothecene 3-O-acetyltransferase protects both the producing organism and transformed yeast from related mycotoxins. Cloning and characterization of Tri101. J. Biol. Chem. 1998, 273, 1654–1661. [Google Scholar]
- Bin-Umer, M.A.; McLaughlin, J.E.; Basu, D.; McCormick, S.; Tumer, N.E. Trichothecene mycotoxins inhibit mitochondrial translation—Implication for the mechanism of toxicity. Toxins 2011, 3, 1484–1501. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol. 2006, 141, 373–437. [Google Scholar] [CrossRef]
- Torres, M.A. ROS in biotic interactions. Physiol. Plant 2010, 138, 414–429. [Google Scholar] [CrossRef]
- Chen, S.-X.; Schopfer, P. Hydroxyl-radical production in physiological reactions: A novel function of peroxidase. Eur. J. Biochem. 1999, 260, 726–735. [Google Scholar] [CrossRef]
- Bradley, D.J.; Kjellbom, P.; Lamb, C.J. Elicitor- and wound-induced oxidative cross-linking of a proline-rich plant cell wall protein: A novel, rapid defense response. Cell 1992, 70, 21–30. [Google Scholar] [CrossRef]
- Hückelhoven, R. Cell wall-associated mechanisms of disease resistance and susceptibility. Annu. Rev. Phytopathol. 2007, 45, 101–127. [Google Scholar] [CrossRef]
- Gupta, R.K.; Banerjee, A.; Pathak, S.; Sharma, C.; Singh, N. Induction of mitochondrial-mediated apoptosis by Morinda citrifolia (noni) in human cervical cancer cells. Asian Pac. J. Cancer Prev. 2013, 14, 237–242. [Google Scholar] [CrossRef]
- Tiessen, C.; Fehr, M.; Schwarz, C.; Baechler, S.; Domnanich, K.; Böttler, U.; Pahlke, G.; Marko, D. Modulation of the cellular redox status by the Alternaria toxins alternariol and alternariol monomethyl ether. Toxicol. Lett. 2013, 216, 23–30. [Google Scholar] [CrossRef]
- Arunachalam, C.; Doohan, F.M. Trichothecene toxicity in eukaryotes: Cellular and molecular mechanisms in plants and animals. Toxicol. Lett. 2013, 21, 149–158. [Google Scholar] [CrossRef]
- Gilchrist, D.G. Mycotoxins reveal connections between plants and animals in apoptosis and ceramide signaling. Cell Death Differ. 1997, 4, 689–698. [Google Scholar]
- Dobosz, B.; Drzewiecka, K.; Waskiewicz, A.; Irzykowska, L.; Bocianowski, J.; Karolewski, Z.; Kostecki, M.; Kruczynski, Z.; Krzyminiewski, R.; Weber, Z.; et al. Free radicals, salicylic acid and mycotoxins in Asparagus after inoculation with Fusarium proliferatum and F. oxysporum. Appl. Magn. Reson. 2011, 41, 19–30. [Google Scholar] [CrossRef]
- Nishiuchi, T.; Masuda, D.; Nakashita, H.; Ichimura, K.; Shinozaki, K.; Yoshida, S.; Kimura, M.; Yamaguchi, I.; Yamaguchi, K. Fusarium phytotoxin trichothecenes have an elicitor-like activity in Arabidopsis thaliana, but the activity differed significantly among their molecular species. Mol. Plant Microbe Interact. 2006, 198, 512–520. [Google Scholar]
- Masuda, D.; Ishida, M.; Yamaguchi, K.; Yamaguchi, I.; Kimura, M.; Nishiuchi, T. Phytotoxic effects of trichothecenes on the growth and morphology of Arabidopsis thaliana. J. Exp. Bot. 2007, 588, 1617–1626. [Google Scholar]
- Desmond, O.J.; Manners, J.M.; Stephens, A.E.; Maclean, D.J.; Schenk, P.M.; Gardiner, D.M.; Munn, A.L.; Kazan, K. The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide production, programmed cell death and defence responses in wheat. Mol. Plant Pathol. 2008, 9, 435–445. [Google Scholar] [CrossRef]
- Parsi, Z.; Gorecki, T. Determination of ergosterol as an indicator of fungal biomass in various samples using non-discriminating flash pyrolysis. J. Chromatogr. A 2006, 1130, 145–150. [Google Scholar]
- Hippelein, M.; Rügamer, M. Ergosterol as an indicator of mould growth on building materials. Int. J. Hyg. Environ. Health 2004, 207, 379–385. [Google Scholar] [CrossRef]
- Bhosle, S.R.; Sandhya, G.; Sonawane, H.B.; Vaidya, J.G. Ergosterol content of several wood decaying fungi using a modified method. Int. J.Pharm. Life Sci. 2011, 2, 916–918. [Google Scholar]
- Bankole, S.A.; Adenusi, A.A.; Lawal, O.S.; Adesanya, O.O. Occurrence of aflatoxin B1 in food products derivable from “egusi” melon seeds consumed in southwestern Nigeria. Food Control 2010, 7, 974–976. [Google Scholar]
- Janardhana, G.R.; Raveesha, K.A.; Shetty, H.S. Mycotoxin contamination of maize grains grown in Karnataka (India). Food Chem. Toxicol. 1999, 37, 863–868. [Google Scholar] [CrossRef]
- Waśkiewicz, A.; Goliński, P.; Karolewski, Z.; Irzykowska, L.; Bocianowski, J.; Kostecki, M.; Weber, Z. Formation of fumonisins and other secondary metabolites by Fusarium oxysporum and F. proliferatum—A comparative study. Food Addit. Contam. 2010, 27, 608–615. [Google Scholar] [CrossRef]
- Waśkiewicz, A.; Wit, M.; Goliński, P.; Chełkowski, J.; Warzecha, R.; Ochodzki, P.; Wakuliński, W. Kinetics of fumonisin B1 formation in maize ears inoculated with Fusarium verticillioides. Food Addit. Contam. 2012, 29, 1752–1761. [Google Scholar] [CrossRef]
- Waśkiewicz, A.; Irzykowska, L.; Bocianowski, J.; Karolewski, Z.; Weber, Z.; Goliński, P. Fusariotoxins in asparagus—their biosynthesis and migration. Food Addit. Contam. 2013, 30, 1332–1338. [Google Scholar] [CrossRef]
- Barbehenn, R.V.; Poopat, U.; Spencer, B. Semiquinone and ascorbyl radicals in the gut fluids of caterpillars measured with EPR spectrometry. Insect Biochem. Mol. Biol. 2003, 33, 125–130. [Google Scholar] [CrossRef]
- Bednarski, W.; Ostrowski, A.; Waplak, S. Low temperature short-range ordering caused by Mn2+ doping of Rb3H(SO4)2. J. Phys. Condens. Matter 2010, 22. [Google Scholar] [CrossRef]
- Morkunas, I.; Formela, M.; Floryszak-Wieczorek, J.; Marczak, Ł.; Narożna, D.; Nowak, W.; Bednarski, W. Cross-talk interactions of exogenous nitric oxide and sucrose modulates phenylpropanoid metabolism in yellow lupine embryo axes infected with Fusarium oxysporum. Plant Sci. 2013, 211, 102–121. [Google Scholar] [CrossRef]
- Mai, V.C.; Bednarski, W.; Borowiak-Sobkowiak, B.; Wilkaniec, B.; Samardakiewicz, S.; Morkunas, I. Oxidative stress in pea seedling leaves in response to Acyrthosiphon pisum infestation. Phytochemistry 2013, 93, 49–62. [Google Scholar] [CrossRef]
- Pearce, R.B.; Edwards, P.P.; Green, T.L.; Anderson, P.A.; Fisher, B.J.; Carpenter, T.A.; Hall, L.D. Immobilized long-lived free radicals at the host–pathogen interface in sycamore (Acer pseudoplatanus L.). Physiol. Mol. Plant Pathol. 1997, 50, 371–390. [Google Scholar] [CrossRef]
- Doke, N. Involvement of superoxide anion generation in the hypersensitive response of potato tuber tissues to infection with an incompatible race of Phytophthora infestans and to the hyphal wall components. Physiol. Mol. Plant Pathol. 1983, 23, 345–355. [Google Scholar] [CrossRef]
- Morkunas, I.; Garnczarska, M.; Bednarski, W.; Ratajczak, W.; Waplak, S. Metabolic and ultrastructural responses of lupine embryo axes to sugar starvation. J. Plant Physiol. 2003, 160, 311–319. [Google Scholar] [CrossRef]
- Morkunas, I.; Bednarski, W.; Kozłowska, M. Response of embryo axes of germinating seeds of yellow lupine to Fusarium oxysporum. Plant Physiol. Biochem. 2004, 42, 493–499. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Waśkiewicz, A.; Morkunas, I.; Bednarski, W.; Mai, V.C.; Formela, M.; Beszterda, M.; Wiśniewska, H.; Goliński, P. Deoxynivalenol and Oxidative Stress Indicators in Winter Wheat Inoculated with Fusarium graminearum. Toxins 2014, 6, 575-591. https://doi.org/10.3390/toxins6020575
Waśkiewicz A, Morkunas I, Bednarski W, Mai VC, Formela M, Beszterda M, Wiśniewska H, Goliński P. Deoxynivalenol and Oxidative Stress Indicators in Winter Wheat Inoculated with Fusarium graminearum. Toxins. 2014; 6(2):575-591. https://doi.org/10.3390/toxins6020575
Chicago/Turabian StyleWaśkiewicz, Agnieszka, Iwona Morkunas, Waldemar Bednarski, Van Chung Mai, Magda Formela, Monika Beszterda, Halina Wiśniewska, and Piotr Goliński. 2014. "Deoxynivalenol and Oxidative Stress Indicators in Winter Wheat Inoculated with Fusarium graminearum" Toxins 6, no. 2: 575-591. https://doi.org/10.3390/toxins6020575