Antitumor Cell-Complex Vaccines Employing Genetically Modified Tumor Cells and Fibroblasts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
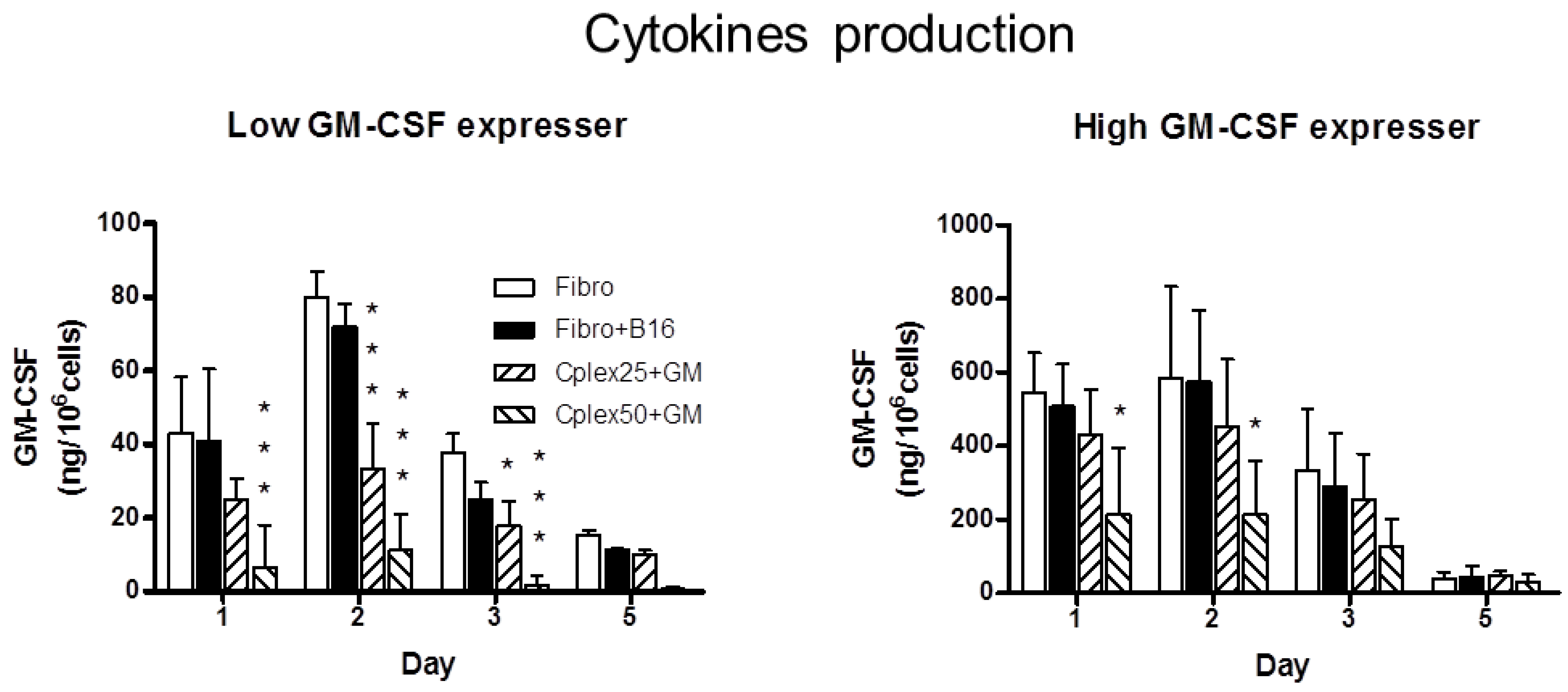
2.1. Cytokine Production

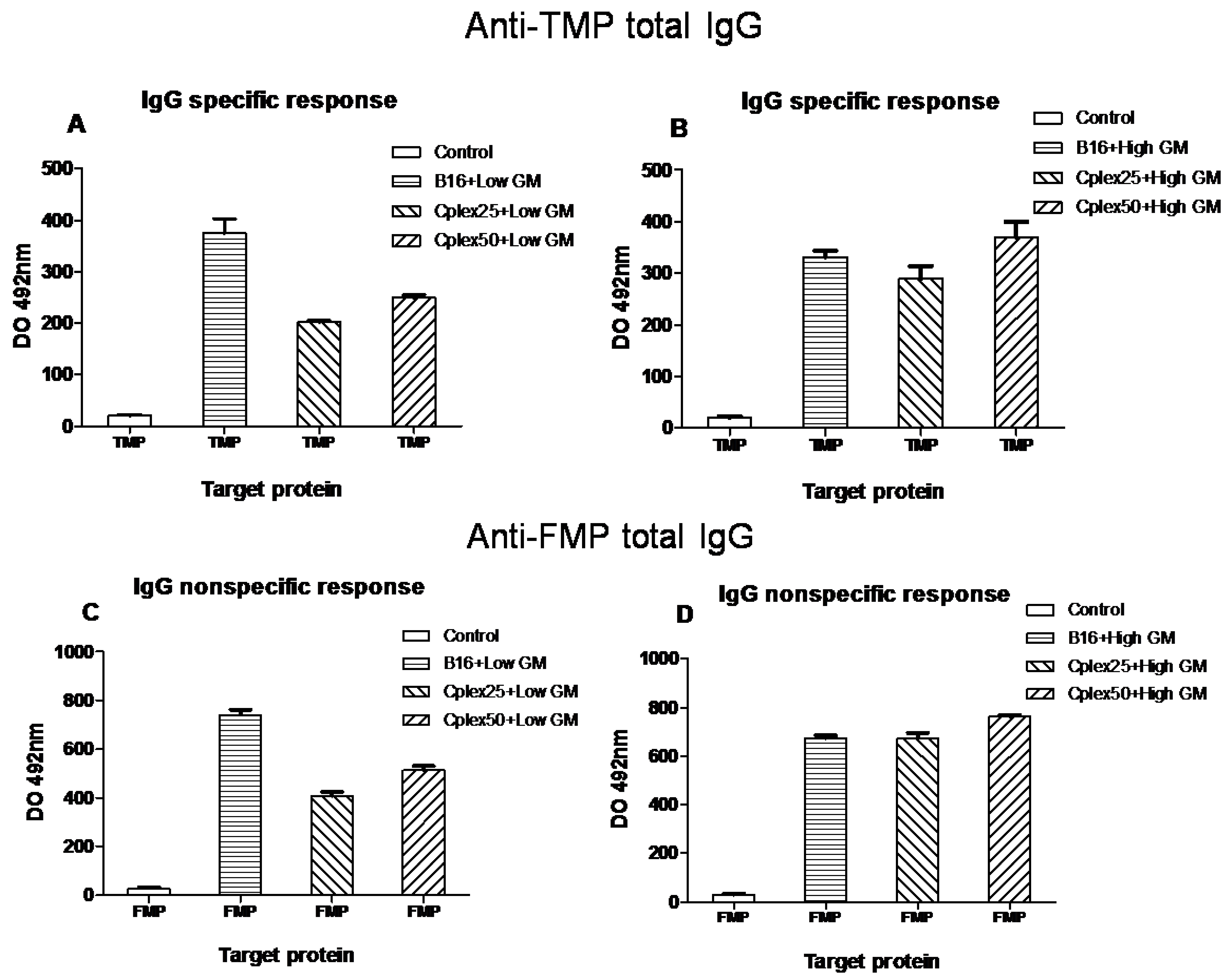
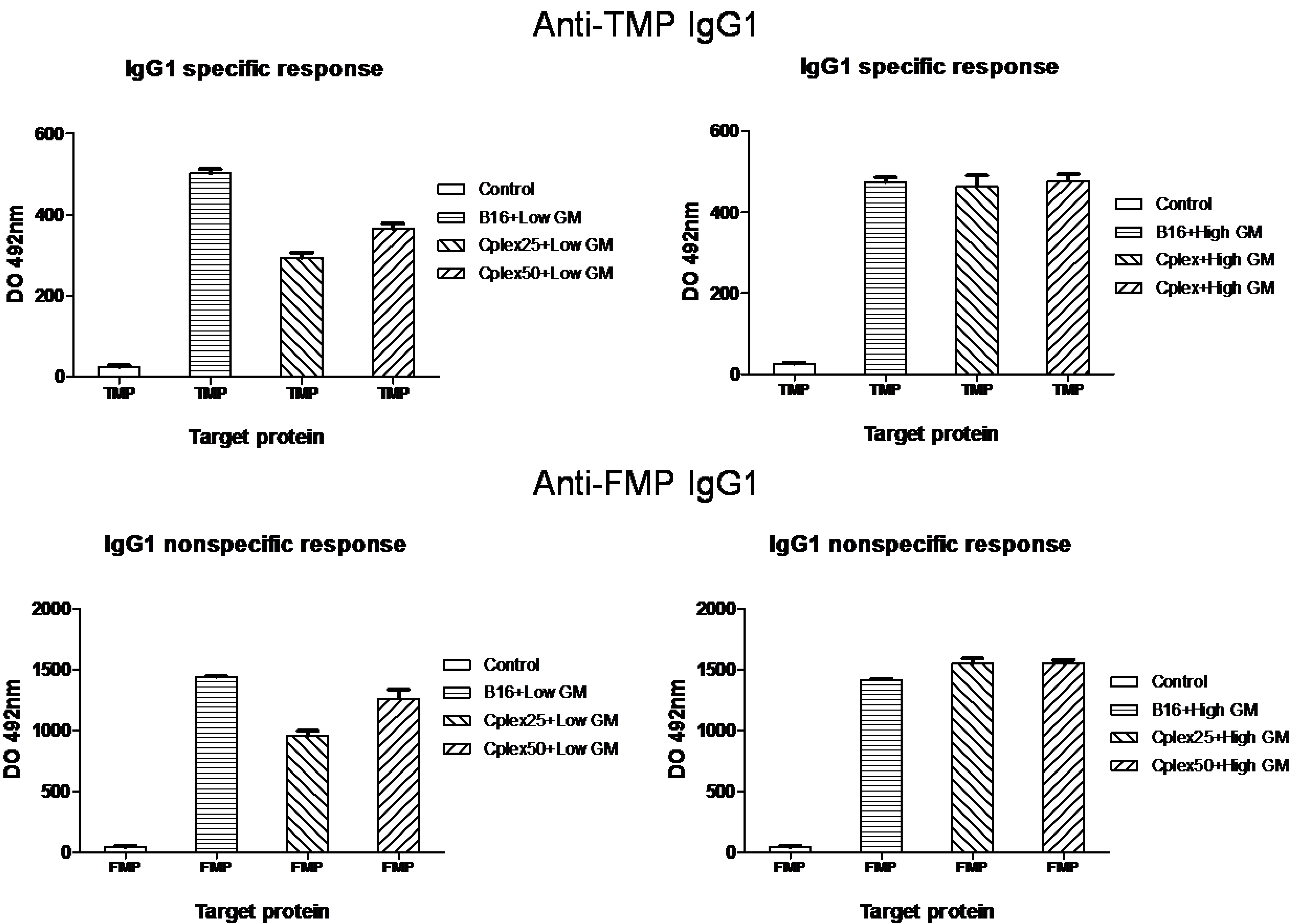
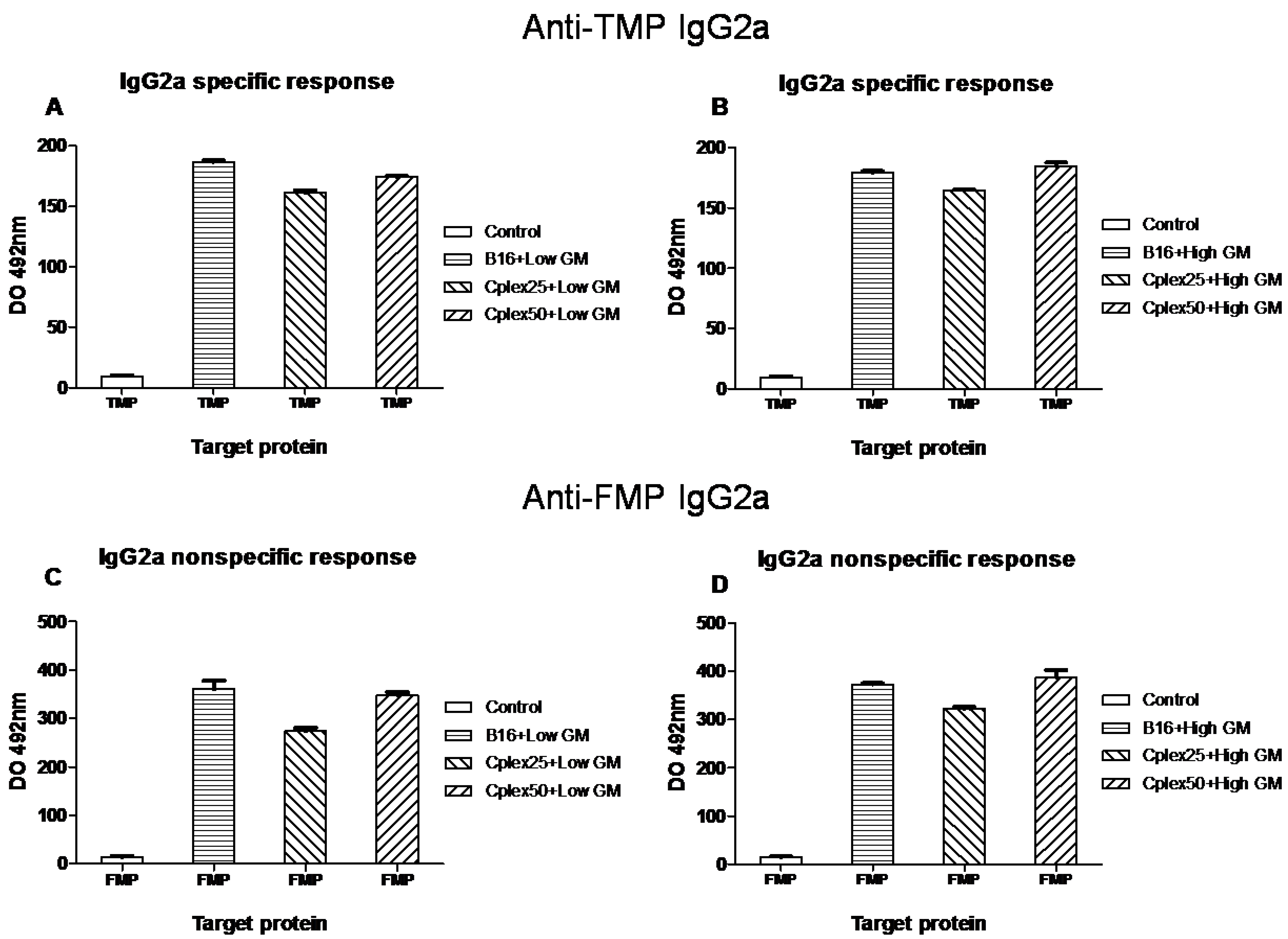
2.2. Specific Antitumor IgG Production



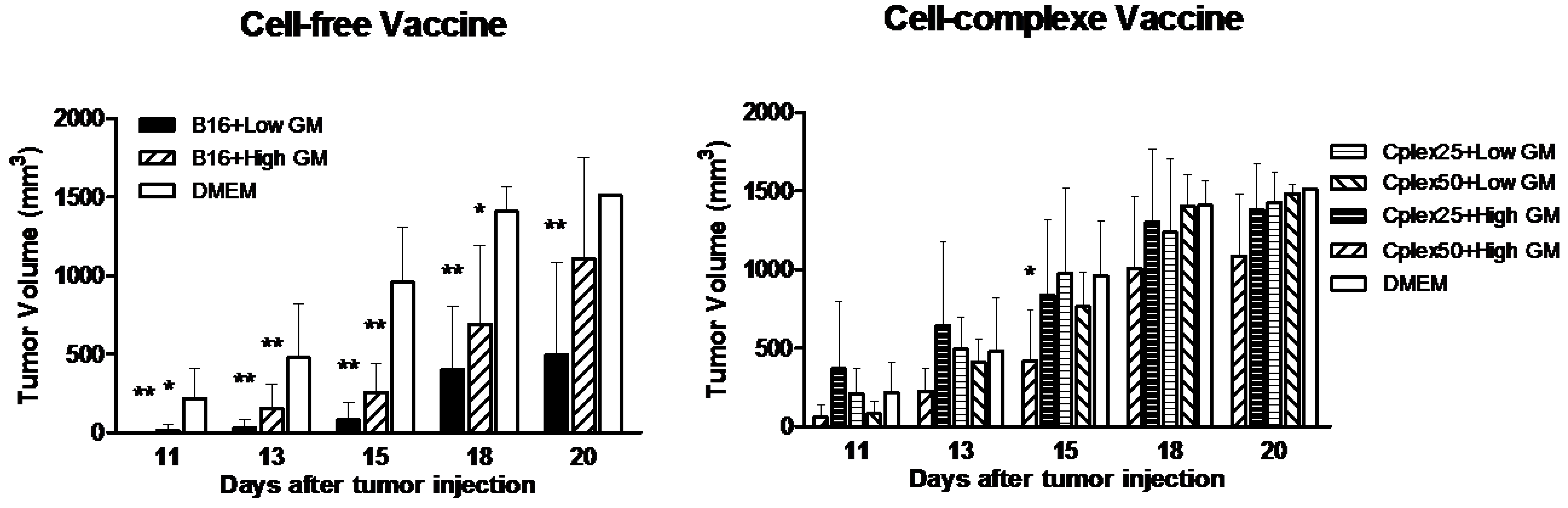
2.3. Tumor Volume

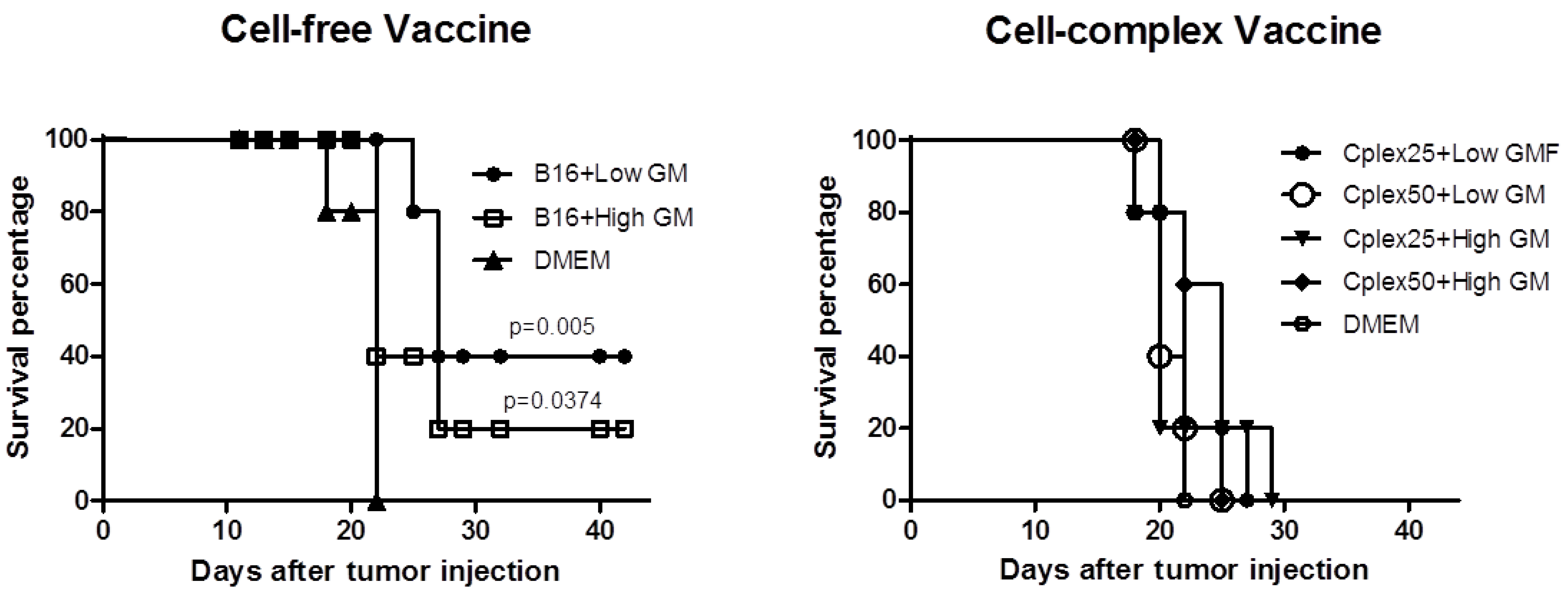
2.4. Animal Survival

3. Experimental Section
3.1. Plasmids
3.2. Cells and Transfection Procedure
3.3. Cytokine Expression ELISA Assay
3.4. Specific Anti-TMP and Anti-FMP IgG ELISA Assay
3.5. Cell Complexes Formation
3.6. Vaccination Design
3.7. Tumor Growth Measurement and Survival
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Dunussi-Joannopoulos, K.; Dranoff, G.; Weinstein, H.J.; Ferrara, J.L.; Bierer, B.E.; Croop, J.M. Gene immunotherapy in murine acute myeloid leukemia: Granulocyte-macrophage colony-stimulating factor tumor cell vaccines elicit more potent antitumor immunity compared with B7 family and other cytokine vaccines. Blood 1998, 91, 222–230. [Google Scholar]
- Small, E.J.; Sacks, N.; Nemunaitis, J.; Urba, W.J.; Dula, E.; Centeno, A.S.; Nelson, W.G.; Ando, D.; Howard, C.; Borellini, F.; et al. Granulocyte macrophage colony-stimulating factor-secreting allogeneic cellular immunotherapy for hormone-refractory prostate cancer. Clin. Cancer Res. 2007, 13, 3883–3891. [Google Scholar] [CrossRef]
- Maio, M.; Fonsatti, E.; Lamaj, E.; Altomonte, M.; Cattarossi, I.; Santantonio, C.; Melani, C.; Belli, F.; Arienti, F.; Colombo, M.P.; et al. Vaccination of stage IV patients with allogeneic IL-4- or IL-2-gene-transduced melanoma cells generates functional antibodies against vaccinating and autologous melanoma cells. Cancer Immunol. Immunother. 2002, 51, 9–14. [Google Scholar]
- Salgia, R.; Lynch, T.; Skarin, A.; Lucca, J.; Lynch, C.; Jung, K.; Hodi, F.S.; Jaklitsch, M.; Mentzer, S.; Swanson, S.; et al. Vaccination with irradiated autologous tumor cells engineered to secrete granulocyte-macrophage colony-stimulating factor augments antitumor immunity in some patients with metastatic non-small-cell lung carcinoma. J. Clin. Oncol. 2003, 21, 624–630. [Google Scholar] [CrossRef]
- Soiffer, R.; Hodi, F.S.; Haluska, F.; Jung, K.; Gillessen, S.; Singer, S.; Tanabe, K.; Duda, R.; Mentzer, S.; Jaklitsch, M.; et al. Vaccination with irradiated, autologous melanoma cells engineered to secrete granulocyte-macrophage colony-stimulating factor by adenoviral-mediated gene transfer augments antitumor immunity in patients with metastatic melanoma. J. Clin. Oncol. 2003, 21, 3343–3350. [Google Scholar] [CrossRef]
- Moret-Tatay, I.; Diaz, J.; Marco, F.M.; Crespo, A.; Aliño, S.F. Complete tumor prevention by engineered tumor cell vaccines employing nonviral vectors. Cancer Gene Ther. 2003, 10, 887–897. [Google Scholar]
- Nemunaitis, J.; Sterman, D.; Jablons, D.; Smith, J.W., II; Fox, B.; Maples, P.; Hamilton, S.; Borellini, F.; Lin, A.; Morali, S.; et al. Granulocyte-macrophage colony-stimulating factor gene-modified autologous tumor vaccines in non-small-cell lung cancer. J. Natl. Cancer Inst. 2004, 96, 326–331. [Google Scholar] [CrossRef]
- Herrero, M.J.; Botella, R.; Dasí, F.; Agás, R.; Sánchez, M.; Aliño, S.F. Antigens and cytokine genes in antitumor vaccines: The importance of the temporal delivery sequence in antitumor signals. Ann. N.Y. Acad. Sci. 2006, 1091, 412–424. [Google Scholar]
- Olivares, J.; Kumar, P.; Yu, Y.; Maples, P.B.; Senzer, N.; Bedell, C.; Barve, M.; Tong, A.; Pappen, B.O.; Kuhn, J.; et al. Phase I trial of TGF-{beta}2 antisense GM-CSF gene-modified autologous tumor cell (TAG) vaccine. Clin. Cancer Res. 2011, 17, 183–192. [Google Scholar] [CrossRef]
- Agarwalla, P.; Barnard, Z.; Fecci, P.; Dranoff, G.; Curry, W.T., Jr. Sequential immunotherapy by vaccination with GM-CSF-expressing glioma cells and CTLA-4 blockade effectively treats established murine intracranial tumors. J. Immunother. 2012, 35, 385–389. [Google Scholar] [CrossRef]
- Zhang, X.; Shi, X.; Li, J.; Hu, Z.; Zhou, D.; Gao, J.; Tan, W. A novel therapeutic vaccine of mouse GM-CSF surface modified MB49 cells against metastatic bladder cancer. J. Urol. 2012, 187, 1071–1079. [Google Scholar]
- Dessureault, S.; Noyes, D.; Lee, D.; Dunn, M.; Janssen, W.; Cantor, A.; Sotomayor, E.; Messina, J.; Antonia, S.J. A phase-I trial using a universal GM-CSF-producing and CD40L-expressing bystander cell line (GM.CD40L) in the formulation of autologous tumor cell-based vaccines for cancer patients with stage IV disease. Ann. Surg. Oncol. 2007, 14, 869–884. [Google Scholar] [CrossRef]
- Brusic, A.; Hainz, U.; Wadleigh, M.; Neuberg, D.; Su, M.; Canning, C.M.; Deangelo, D.J.; Stone, R.M.; Lee, J.S.; Mulligan, R.C.; et al. Detecting T-cell reactivity to whole cell vaccines: Proof of concept analysis of T-cell response to K562 cell antigens in CML patients. Oncoimmunology 2012, 1, 1095–1103. [Google Scholar] [CrossRef]
- Creelan, B.C.; Antonia, S.; Noyes, D.; Hunter, T.B.; Simon, G.R.; Bepler, G.; Williams, C.C.; Tanvetyanon, T.; Haura, E.B.; Schell, M.J.; et al. Phase II trial of a GM-CSF-producing and CD40L-expressing bystander cell line combined with an allogeneic tumor cell-based vaccine for refractory lung adenocarcinoma. J. Immunother. 2013, 36, 442–450. [Google Scholar] [CrossRef]
- Veelken, H.; Mackensen, A.; Lahn, M.; Kohler, G.; Becker, D.; Franke, B.; Brennscheidt, U.; Kulmburg, P.; Rosenthal, F.M.; Keller, H.; et al. A phase-I clinical study of autologous tumor cells plus interleukin-2-gene-transfected allogeneic fibroblasts as a vaccine in patients with cancer. Int. J. Cancer 1997, 70, 269–277. [Google Scholar] [CrossRef]
- Sobol, R.E.; Shawler, D.L.; Carson, C.; Van Beveren, C.; Mercola, D.; Fakhrai, H.; Garrett, M.A.; Barone, R.; Goldfarb, P.; Bartholomew, R.M.; et al. Interleukin 2 gene therapy of colorectal carcinoma with autologous irradiated tumor cells and genetically engineered fibroblasts: A Phase I study. Clin. Cancer Res. 1999, 5, 2359–2365. [Google Scholar]
- Kang, W.K.; Park, C.; Yoon, H.L.; Kim, W.S.; Yoon, S.S.; Lee, M.H.; Park, K.; Kim, K.; Jeong, H.S.; Kim, J.A.; et al. Interleukin 12 gene therapy of cancer by peritumoral injection of transduced autologous fibroblasts: Outcome of a phase I study. Hum. Gene Ther. 2001, 12, 671–684. [Google Scholar] [CrossRef]
- Okada, H.; Lieberman, F.S.; Edington, H.D.; Witham, T.F.; Wargo, M.J.; Cai, Q.; Elder, E.H.; Whiteside, T.L.; Schold, S.C., Jr.; Pollack, I.F. Autologous glioma cell vaccine admixed with interleukin-4 gene transfected fibroblasts in the treatment of recurrent glioblastoma: preliminary observations in a patient with a favorable response to therapy. J. Neurooncol. 2003, 64, 13–20. [Google Scholar]
- Barker, S.E.; Grosse, S.M.; Siapati, E.K.; Kritz, A.; Kinnon, C.; Thrasher, A.J.; Hart, S.L. Immunotherapy for neuroblastoma using syngeneic fibroblasts transfected with IL-2 and IL-12. Br. J. Cancer 2007, 97, 210–217. [Google Scholar]
- Okada, H.; Lieberman, F.S.; Walter, K.A.; Lunsford, L.D.; Kondziolka, D.S.; Bejjani, G.K.; Hamilton, R.L.; Torres-Trejo, A.; Kalinski, P.; Cai, Q.; et al. Autologous glioma cell vaccine admixed with interleukin-4 gene transfected fibroblasts in the treatment of patients with malignant gliomas. J. Transl. Med. 2007, 5, 67. [Google Scholar] [CrossRef] [Green Version]
- Palumbo, R.N.; Zhing, X.; Wang, C. Polymer-mediated DNA vaccine delivery via bystander cells requires a proper balance between transfection efficiency and cytoxity. J. Control. Release 2012, 157, 86–93. [Google Scholar] [CrossRef]
- Godbey, W.T.; Wu, K.K.; Mikos, A.G. Poly(ethylenimine) and its role in gene delivery. J. Control. Release 1999, 60, 149–160. [Google Scholar] [CrossRef]
- Godbey, W.T.; Barry, M.A.; Saggau, P.; Wu, K.K.; Mikos, A.G. Poly(ethylenimine)-mediated transfection: A new paradigm for gene delivery. J. Biome Mater. Res. 2000, 51, 321–328. [Google Scholar] [CrossRef]
- Godbey, W.T.; Mikos, A.G. Recent progress in gene delivery using non-viral transfer complexes. J. Control. Release 2001, 72, 115–125. [Google Scholar] [CrossRef]
- Guillem, V.M.; Tormo, M.; Revert, F.; Benet, I.; García-Conde, J.; Crespo, A.; Aliño, S.F. Polyethyleneimine-based immunopolyplex for targeted gene transfer in human lymphoma cell lines. J. Gene Med. 2002, 4, 170–182. [Google Scholar] [CrossRef]
- Guillem, V.M.; Tormo, M.; Moret, I.; Benet, I.; García-Conde, J.; Crespo, A.; Aliño, S.F. Targeted oligonucleotide delivery in human lymphoma cell lines using a polyethyleneimine based immunopolyplex. J. Control. Release 2002, 83, 133–146. [Google Scholar] [CrossRef]
- Nagai, H.; Oniki, S.; Fujiwara, S.; Yoshimoto, T.; Nishigori, C. Antimelanoma immunotherapy: Clinical and preclinical applications of IL-12 family members. Immunotherapy 2010, 2, 697–709. [Google Scholar] [CrossRef]
- Borrello, I.; Pardoll, D. GM-CSF-based cellular vaccines: A review of the clinical experience. Cytokine Growth Factor Rev. 2002, 13, 185–193. [Google Scholar] [CrossRef]
- Miguel, A.; Herrero, M.J.; Sendra, L.; Botella, R.; Algás, R.; Sánchez, M.; Aliño, S.F. Comparative antitumor effect of preventive versus therapeutic vaccines employing B16 melanoma cells genetically modified to express GM-CSF and B7.2 in a murine model. Toxins 2012, 4, 1058–1081. [Google Scholar]
- Miguel, A.; Herrero, M.J.; Sendra, L.; Botella, R.; Algás, R.; Sánchez, M.; Aliño, S.F. Comparative antitumor effect among GM-CSF, IL-12 and GM-CSF+IL-12 genetically modified tumor cell vaccines. Cancer Gene Ther. 2013, 20, 576–581. [Google Scholar]
- Serafini, P.; Carbley, R.; Noonan, K.A.; Tan, G.; Bronte, V.; Borrello, I. High-dose granulocyte-macrophage colony-stimulating factor-producing vaccines impair the immune response through the recruitment of myeloid suppressor cells. Cancer Res. 2004, 64, 6337–6343. [Google Scholar] [CrossRef]
- Guillem, V.M.; Aliño, S.F. Transfection pathways of nonspecific and targeted PEI-polyplexes. Gene Ther. Mol. Biol. 2004, 8, 369–384. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Miguel, A.; Herrero, M.J.; Sendra, L.; Botella, R.; Diaz, A.; Algás, R.; Aliño, S.F. Antitumor Cell-Complex Vaccines Employing Genetically Modified Tumor Cells and Fibroblasts. Toxins 2014, 6, 636-649. https://doi.org/10.3390/toxins6020636
Miguel A, Herrero MJ, Sendra L, Botella R, Diaz A, Algás R, Aliño SF. Antitumor Cell-Complex Vaccines Employing Genetically Modified Tumor Cells and Fibroblasts. Toxins. 2014; 6(2):636-649. https://doi.org/10.3390/toxins6020636
Chicago/Turabian StyleMiguel, Antonio, María José Herrero, Luis Sendra, Rafael Botella, Ana Diaz, Rosa Algás, and Salvador F. Aliño. 2014. "Antitumor Cell-Complex Vaccines Employing Genetically Modified Tumor Cells and Fibroblasts" Toxins 6, no. 2: 636-649. https://doi.org/10.3390/toxins6020636