Staphylococcal Bicomponent Pore-Forming Toxins: Targets for Prophylaxis and Immunotherapy

Abstract
:1. Introduction
2. Staphylococcal Bicomponent Pore-Forming Toxins (BCPFTs)
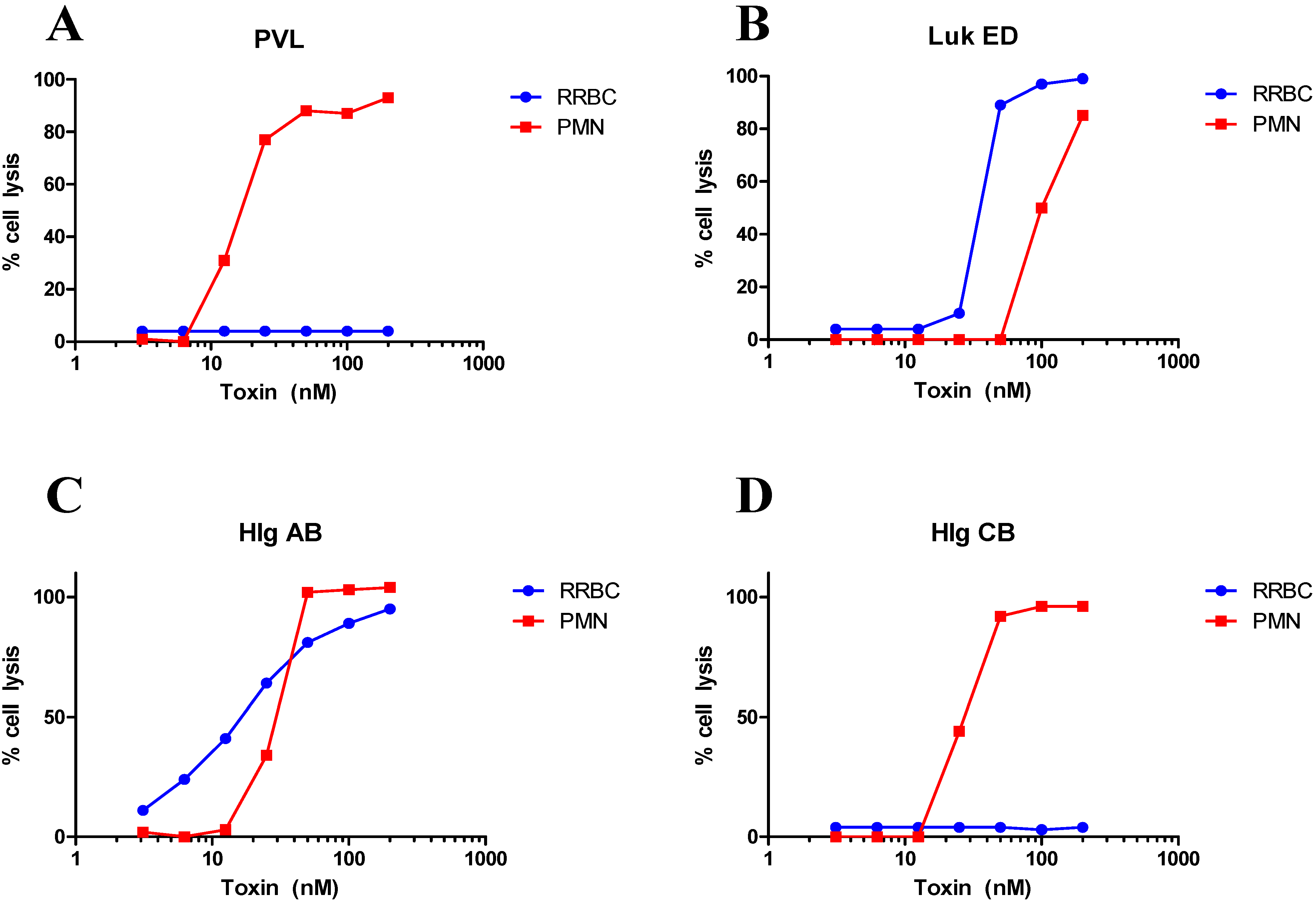
3. Tropism and Species Specificity

{kind=link}
{kind=link}
{kind=link}
| Cells and Species | Toxin | Effect | References |
|---|---|---|---|
| Human PMN | PVL | Receptor binding | [36,38] |
| Lysis | [36,38,65,66] | ||
| Ca++ influx | [36,67] | ||
| Proinflammatory response | [38,66] | ||
| LukED | Lysis | [63,68,69] and Figure 1 | |
| Receptor binding | [68] | ||
| Ca++ influx | [69,70] | ||
| Proinflammatory response | [69] | ||
| LukAB (LukGH) | Receptor binding | [71] | |
| Lysis | [61,66,72,73,74] | ||
| HlgAB/CB | Lysis | [63] and Figure 1 | |
| Murine PMN | PVL | No effect | [65,66] |
| Proinflammatory response | [26,75] | ||
| LukED | Lysis | [34,60] | |
| LukAB | No effect | [71] | |
| Lysis | [72] | ||
| Rabbit PMN | PVL | Lysis | [19,66] |
| Proinflammatory response | [66] | ||
| LukAB | Lysis | [72] | |
| Human Monocytes and Macrophages | PVL | Receptor binding | [36,38] |
| Proinflammatory response | [76] | ||
| LukAB LukED | Lysis | [61] | |
| Lysis | [34,68] | ||
| Human T cells | LukED | Lysis | [34] |
| Human Natural Killer cells | LukED | Lysis | [68] |
| Murine Monocytes and Macrophages | PVL | Proinflammatory response | [75] |
| LukED | Lysis | [60] | |
| Human DC | LukAB | Lysis | [61] |
| LukED | Lysis | [34] | |
| Human RBC | Hlg | Lysis | [28] |
| Rabbit RBC | Hlg | Lysis | [11,39,77,78] |
| LukED | No effect | [79] | |
| Lysis | [80] and Figure 1 |
4. Cellular Receptors
5. Nine Subunits; How Many Toxins?


6. Cell Lysis versus Pro-inflammatory Activity
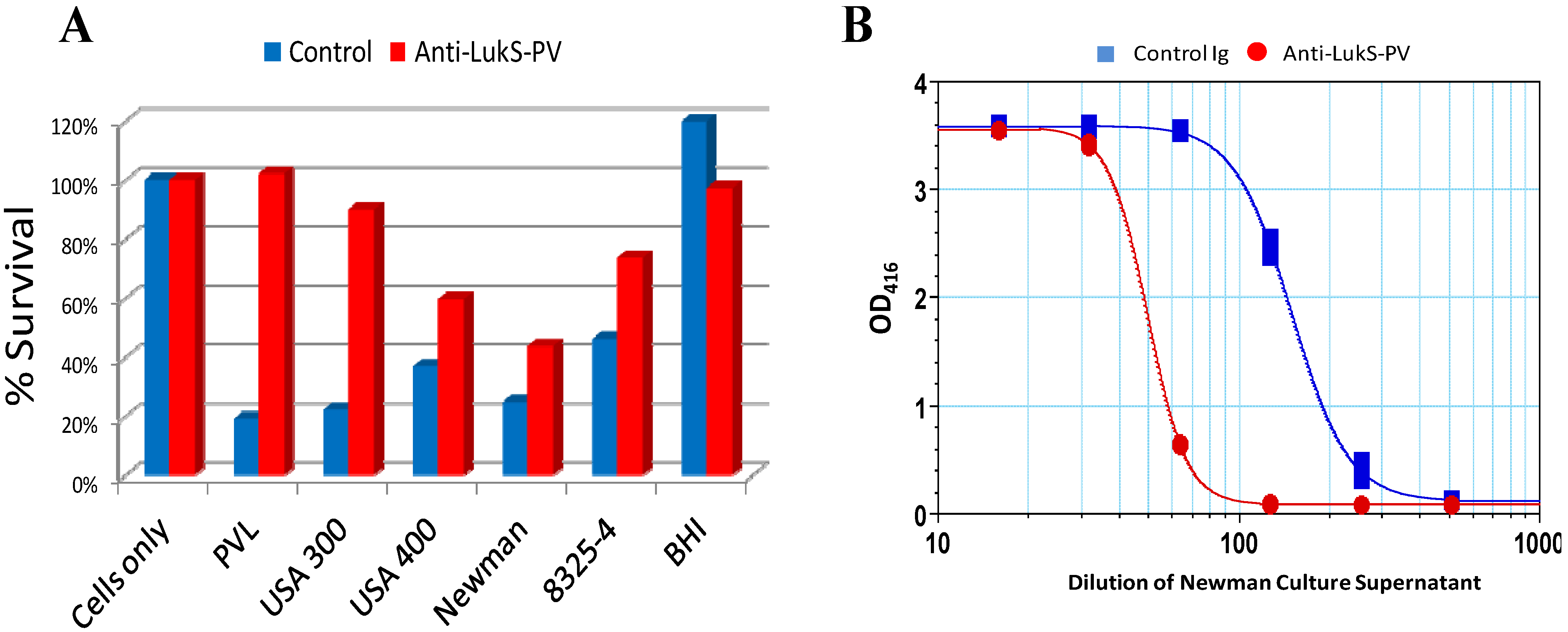
7. Leukotoxins as Targets for Prophylaxis and Immunotherapy
| Class | Subunit | S-Subunit | F-Subunit | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| LukS-PV | Hlg C | Hlg A | Luk E | Luk A | LukF-PV | Hlg B | Luk D | Luk B | ||
| S-Subunit | LukS-PV | 100 | 82 | 68 | 71 | 33 | 30 | 28 | 31 | 27 |
| Hlg C | 100 | 70 | 70 | 34 | 30 | 28 | 30 | 27 | ||
| Hlg A | 100 | 70 | 35 | 29 | 29 | 31 | 28 | |||
| Luk E | 100 | 36 | 28 | 29 | 30 | 27 | ||||
| Luk A | 100 | 28 | 28 | 30 | 31 | |||||
| F-Subunit | LukF-PV | 100 | 72 | 82 | 39 | |||||
| Hlg B | 100 | 76 | 40 | |||||||
| Luk D | 100 | 39 | ||||||||
| Luk B | 100 | |||||||||

8. Concluding Remarks
Conflicts of Interest
References
- Nizet, V. Understanding how leading bacterial pathogens subvert innate immunity to reveal novel therapeutic targets. J. Allergy Clin. Immunol. 2007, 120, 13–22. [Google Scholar] [CrossRef]
- Kotzin, B.L.; Leung, D.Y.; Kappler, J.; Marrack, P. Superantigens and their potential role in human disease. Adv. Immunol. 1993, 54, 99–166. [Google Scholar] [CrossRef]
- Meyer, R.D.; Monday, S.R.; Bohach, G.A.; Schlievert, S.M. Prolonged course of toxic shock syndrome associated with methicillin-resistant staphylococcus aureus enterotoxins G and I. Int. J. Infect. Dis. 2001, 5, 163–166. [Google Scholar] [CrossRef]
- Schuberth, H.J.; Krueger, C.; Zerbe, H.; Bleckmann, E.; Leibold, W. Characterization of leukocytotoxic and superantigen-like factors produced by staphylococcus aureus isolates from milk of cows with mastitis. Vet. Microbiol. 2001, 82, 187–189. [Google Scholar] [CrossRef]
- Tristan, A.; Ferry, T.; Durand, G.; Dauwalder, O.; Bes, M.; Lina, G.; Vandenesch, F.; Etienne, J. Virulence determinants in community and hospital meticillin-resistant staphylococcus aureus. J. Hosp. Infect. 2007, 65 (Suppl. 2), 105–109. [Google Scholar]
- King, M.D.; Humphrey, B.J.; Wang, Y.F.; Kourbatova, E.V.; Ray, S.M.; Blumberg, H.M. Emergence of community-acquired methicillin-resistant staphylococcus aureus usa 300 clone as the predominant cause of skin and soft-tissue infections. Ann. Intern. Med. 2006, 144, 309–317. [Google Scholar] [CrossRef]
- Moran, G.J.; Krishnadasan, A.; Gorwitz, R.J.; Fosheim, G.E.; McDougal, L.K.; Carey, R.B.; Talan, D.A. Methicillin-esistant S. aureus infections among patients in the emergency department. N. Engl. J. Med. 2006, 355, 666–674. [Google Scholar] [CrossRef]
- Nichol, K.A.; Adam, H.J.; Hussain, Z.; Mulvey, M.R.; McCracken, M.; Mataseje, L.F.; Thompson, K.; Kost, S.; Lagace-Wiens, P.R.; Hoban, D.J.; et al. Comparison of community-associated and health care-associated methicillin-resistant staphylococcus aureus in Canada: Results of the canward 2007–2009 study. Diagn. Microbiol. Infect. Dis. 2011, 69, 320–325. [Google Scholar] [CrossRef]
- Diep, B.A.; Otto, M. The role of virulence determinants in community-associated mrsa pathogenesis. Trends Microbiol. 2008, 16, 361–369. [Google Scholar] [CrossRef]
- Diep, B.A.; Gill, S.R.; Chang, R.F.; Phan, T.H.; Chen, J.H.; Davidson, M.G.; Lin, F.; Lin, J.; Carleton, H.A.; Mongodin, E.F.; Sensabaugh, G.F.; Perdreau-Remington, F. Complete genome sequence of usa300, an epidemic clone of community-acquired meticillin-resistant staphylococcus aureus. Lancet 2006, 367, 731–739. [Google Scholar] [CrossRef]
- Kaneko, J.; Kimura, T.; Narita, S.; Tomita, T.; Kamio, Y. Complete nucleotide sequence and molecular characterization of the temperate staphylococcal bacteriophage phipvl carrying panton-valentine leukocidin genes. Gene 1998, 215, 57–67. [Google Scholar] [CrossRef]
- Narita, S.; Kaneko, J.; Chiba, J.; Piemont, Y.; Jarraud, S.; Etienne, J.; Kamio, Y. Phage conversion of panton-valentine leukocidin in staphylococcus aureus: Molecular analysis of a pvl-converting phage, PhiSLT. Gene 2001, 268, 195–206. [Google Scholar] [CrossRef]
- Panton, P.N.; Valentine, F.C. Staphylococcal toxin. Lancet 1932, 222, 506–508. [Google Scholar] [CrossRef]
- Naimi, T.S.; LeDell, K.H.; Como-Sabetti, K.; Borchardt, S.M.; Boxrud, D.J.; Etienne, J.; Johnson, S.K.; Vandenesch, F.; Fridkin, S.; O’Boyle, C.; et al. Comparison of community-and health care-associated methicillin-resistant staphylococcus aureus infection. JAMA 2003, 290, 2976–2984. [Google Scholar] [CrossRef]
- Lina, G.; Piemont, Y.; Godail-Gamot, F.; Bes, M.; Peter, M.O.; Gauduchon, V.; Vandenesch, F.; Etienne, J. Involvement of panton-valentine leukocidin-producing staphylococcus aureus in primary skin infections and pneumonia. Clin. Infect. Dis. 1999, 29, 1128–1132. [Google Scholar] [CrossRef]
- Prevost, G.; Couppie, P.; Prevost, P.; Gayet, S.; Petiau, P.; Cribier, B.; Monteil, H.; Piemont, Y. Epidemiological data on staphylococcus aureus strains producing synergohymenotropic toxins. J. Med. Microbiol. 1995, 42, 237–245. [Google Scholar] [CrossRef]
- Gillet, Y.; Issartel, B.; Vanhems, P.; Fournet, J.C.; Lina, G.; Bes, M.; Vandenesch, F.; Piemont, Y.; Brousse, N.; Floret, D.; et al. Association between staphylococcus aureus strains carrying gene for panton-valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet 2002, 359, 753–759. [Google Scholar] [CrossRef]
- Vandenesch, F.; Naimi, T.; Enright, M.C.; Lina, G.; Nimmo, G.R.; Heffernan, H.; Liassine, N.; Bes, M.; Greenland, T.; Reverdy, M.E.; et al. Community-acquired methicillin-resistant staphylococcus aureus carrying panton-valentine leukocidin genes: worldwide emergence. Emerg. Infect. Dis. 2003, 9, 978–984. [Google Scholar] [CrossRef]
- Diep, B.A.; Chan, L.; Tattevin, P.; Kajikawa, O.; Martin, T.R.; Basuino, L.; Mai, T.T.; Marbach, H.; Braughton, K.R.; Whitney, A.R.; et al. Polymorphonuclear leukocytes mediate staphylococcus aureus panton-valentine leukocidin-induced lung inflammation and injury. Proc. Natl. Acad. Sci. USA 2010, 107, 5587–5592. [Google Scholar] [CrossRef]
- Diep, B.A.; Palazzolo-Ballance, A.M.; Tattevin, P.; Basuino, L.; Braughton, K.R.; Whitney, A.R.; Chen, L.; Kreiswirth, B.N.; Otto, M.; DeLeo, F.R.; et al. Contribution of panton-valentine leukocidin in community-associated methicillin-resistant staphylococcus aureus pathogenesis. PLoS ONE 2008, 3, e3198. [Google Scholar] [CrossRef]
- Karauzum, H.; Adhikari, R.P.; Sarwar, J.; Devi, V.S.; Abaandou, L.; Haudenschild, C.; Mahmoudieh, M.; Boroun, A.R.; Vu, H.; Nguyen, T.; et al. Structurally designed attenuated subunit vaccines for S. Aureus Luks-PV and Lukf-PV confer protection in a mouse bacteremia model. PLoS ONE 2013, 8, e65384. [Google Scholar] [CrossRef]
- Zaidi, T.; Yoong, P.; Pier, G.B. Staphylococcus aureus corneal infections: Effect of the panton-valentine leukocidin (Pvl) and antibody to Pvl on virulence and pathology. Invest. Ophthalmol. Vis. Sci. 2013, 54, 4430–4438. [Google Scholar] [CrossRef]
- Lipinska, U.; Hermans, K.; Meulemans, L.; Dumitrescu, O.; Badiou, C.; Duchateau, L.; Haesebrouck, F.; Etienne, J.; Lina, G. Panton-Valentine Leukocidin Does Play a Role in the Early Stage of Staphylococcus Aureus Skin Infections: A Rabbit Model. PLoS ONE 2011, 6, e22864. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.J.; Kobayashi, S.D.; Ayeras, A.A.; Ashraf, M.; Graves, S.F.; Ragasa, W.; Humbird, T.; Greaver, J.L.; Cantu, C.; Swain, J.L.; et al. Lack of a major role of staphylococcus aureus panton-valentine leukocidin in lower respiratory tract infection in nonhuman primates. Am. J. Pathol. 2010, 176, 1346–1354. [Google Scholar] [CrossRef]
- Voyich, J.M.; Otto, M.; Mathema, B.; Braughton, K.R.; Whitney, A.R.; Welty, D.; Long, R.D.; Dorward, D.W.; Gardner, D.J.; Lina, G.; et al. Is panton-valentine leukocidin the major virulence determinant in community-associated methicillin-resistant staphylococcus aureus disease? J. Infect. Dis. 2006, 194, 1761–1770. [Google Scholar] [CrossRef]
- Yoong, P.; Pier, G.B. Antibody-mediated enhancement of community-acquired methicillin-resistant staphylococcus aureus infection. Proc. Natl. Acad. Sci. USA 2010, 107, 2241–2246. [Google Scholar] [CrossRef]
- Yoong, P.; Pier, G.B. Immune-activating properties of panton-valentine leukocidin improve the outcome in a model of methicillin-resistant staphylococcus aureus pneumonia. Infect. Immun. 2012, 80, 2894–2904. [Google Scholar] [CrossRef]
- Kaneko, J.; Kamio, Y. Bacterial two-component and hetero-heptameric pore-forming cytolytic toxins: structures, pore-forming mechanism, and organization of the genes. Biosci. Biotechnol. Biochem. 2004, 68, 981–1003. [Google Scholar] [CrossRef]
- Los, F.C.; Randis, T.M.; Aroian, R.V.; Ratner, A.J. Role of pore-forming toxins in bacterial infectious diseases. Microbiol. Mol. Biol. Rev. 2013, 77, 173–207. [Google Scholar] [CrossRef]
- Vandenesch, F.; Lina, G.; Henry, T. Staphylococcus aureus hemolysins, bi-component leukocidins, and cytolytic peptides: A redundant arsenal of membrane-damaging virulence factors? Front. Cell. Infect. Microbiol. 2012, 2, 12:1–12:29. [Google Scholar]
- Murphy, R.A.; Haque, R. Purification and properties of staphylococcal delta-hemolysin. I. Production of delta-hemolysin. J. Bacteriol. 1967, 94, 1327–1333. [Google Scholar]
- Otto, M. Staphylococcus aureus toxin gene hitchhikes on a transferable antibiotic resistance element. Virulence 2010, 1, 49–51. [Google Scholar] [CrossRef]
- Bhakdi, S.; Tranum-Jensen, J. Alpha-toxin of staphylococcus aureus. Microbiol. Rev. 1991, 55, 733–751. [Google Scholar]
- Alonzo, F., 3rd; Kozhaya, L.; Rawlings, S.A.; Reyes-Robles, T.; DuMont, A.L.; Myszka, D.G.; Landau, N.R.; Unutmaz, D.; Torres, V.J. CCR5 is a receptor for staphylococcus aureus leukotoxin ED. Nature 2013, 493, 51–55. [Google Scholar]
- Colin, D.A.; Mazurier, I.; Sire, S.; Finck-Barbancon, V. Interaction of the two components of leukocidin from staphylococcus aureus with human polymorphonuclear leukocyte membranes: sequential binding and subsequent activation. Infect. Immun. 1994, 62, 3184–3188. [Google Scholar]
- Gauduchon, V.; Werner, S.; Prevost, G.; Monteil, H.; Colin, D.A. Flow cytometric determination of panton-valentine leucocidin S component binding. Infect. Immun. 2001, 69, 2390–2395. [Google Scholar] [CrossRef]
- Meyer, F.; Girardot, R.; Piemont, Y.; Prevost, G.; Colin, D.A. Analysis of the specificity of panton-valentine leucocidin and gamma-hemolysin F component binding. Infect. Immun. 2009, 77, 266–273. [Google Scholar] [CrossRef]
- Spaan, A.N.; Henry, T.; van Rooijen, W.J.; Perret, M.; Badiou, C.; Aerts, P.C.; Kemmink, J.; de Haas, C.J.; van Kessel, K.P.; Vandenesch, F.; et al. The staphylococcal toxin panton-valentine leukocidin targets human C5a receptors. Cell Host Microbe 2013, 13, 584–594. [Google Scholar] [CrossRef]
- Ozawa, T.; Kaneko, J.; Kamio, Y. Essential binding of lukf of staphylococcal gamma-hemolysin followed by the binding of H. gamma Ii for the hemolysis of human erythrocytes. Biosci. Biotechnol. Biochem. 1995, 59, 1181–1183. [Google Scholar] [CrossRef]
- Sugawara, N.; Tomita, T.; Sato, T.; Kamio, Y. Assembly of staphylococcus aureus leukocidin into a pore-forming ring-shaped oligomer on human polymorphonuclear leukocytes and rabbit erythrocytes. Biosci. Biotechnol. Biochem. 1999, 63, 884–891. [Google Scholar] [CrossRef]
- Song, L.; Hobaugh, M.R.; Shustak, C.; Cheley, S.; Bayley, H.; Gouaux, J.E. Structure ofstaphylococcal alpha-hemolysin, a heptameric transmembrane pore. Science 1996, 274, 1859–1866. [Google Scholar] [CrossRef]
- Olson, R.; Nariya, H.; Yokota, K.; Kamio, Y.; Gouaux, E. Crystal structure of staphylococcal lukf delineates conformational changes accompanying formation of a transmembrane channel. Nat. Struct. Biol. 1999, 6, 134–140. [Google Scholar] [CrossRef]
- Pedelacq, J.D.; Maveyraud, L.; Prevost, G.; Baba-Moussa, L.; Gonzalez, A.; Courcelle, E.; Shepard, W.; Monteil, H.; Samama, J.P.; Mourey, L. The structure of a staphylococcus aureus leucocidin component (Lukf-PV) reveals the fold of the water-soluble species of a family of transmembrane pore-forming toxins. Structure 1999, 7, 277–287. [Google Scholar] [CrossRef]
- Aman, M.J.; Karauzum, H.; Bowden, M.G.; Nguyen, T.L. Structural model of the pre-pore ring-like structure of panton-valentine leukocidin: Providing dimensionality to biophysical and mutational data. J. Biomol. Struct. Dyn. 2010, 28, 1–12. [Google Scholar]
- Miles, G.; Movileanu, L.; Bayley, H. Subunit composition of a bicomponent toxin: Staphylococcal leukocidin forms an octameric transmembrane pore. Protein. Sci. 2002, 11, 894–902. [Google Scholar] [CrossRef]
- Jayasinghe, L.; Bayley, H. The Leukocidin Pore: Evidence for an octamer with four lukf subunits and four luks subunits alternating around a central axis. Protein. Sci. 2005, 14, 2550–2561. [Google Scholar] [CrossRef]
- Spaan, A.N.; Surewaard, B.G.; Nijland, R.; van Strijp, J.A. Neutrophils versus staphylococcus aureus: A biological tug of war. Annu. Rev. Microbiol. 2013, 67, 629–650. [Google Scholar] [CrossRef]
- Zou, D.; Kaneko, J.; Narita, S.; Kamio, Y. Prophage, phipv83-pro, carrying panton-valentine leukocidin genes, on the staphylococcus aureus P83 chromosome: comparative analysis of the genome structures of Phipv83-Pro, Phipvl, Phi11, and other phages. Biosci. Biotechnol. Biochem. 2000, 64, 2631–2643. [Google Scholar] [CrossRef]
- Younis, A.; Krifucks, O.; Fleminger, G.; Heller, E.D.; Gollop, N.; Saran, A.; Leitner, G. Staphylococcus aureus leucocidin, a virulence factor in bovine mastitis. J. Dairy Res. 2005, 72, 188–194. [Google Scholar] [CrossRef]
- Fromageau, A.; Cunha, P.; Gilbert, F.B.; Rainard, P. Purified staphylococcus aureus leukotoxin lukm/f' does not trigger inflammation in the bovine mammary gland. Microb. Pathog. 2011, 51, 396–401. [Google Scholar] [CrossRef]
- Labandeira-Rey, M.; Couzon, F.; Boisset, S.; Brown, E.L.; Bes, M.; Benito, Y.; Barbu, E.M.; Vazquez, V.; Hook, M.; et al. Staphylococcus aureus panton-valentine leukocidin causes necrotizing pneumonia. Science 2007, 315, 1130–1133. [Google Scholar] [CrossRef]
- Shallcross, L.J.; Fragaszy, E.; Johnson, A.M.; Hayward, A.C. The role of the panton-valentine leucocidin toxin in staphylococcal disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 13, 43–54. [Google Scholar]
- Wannet, W.J.; Spalburg, E.; Heck, M.E.; Pluister, G.N.; Tiemersma, E.; Willems, R.J.; Huijsdens, X.W.; de Neeling, A.J.; Etienne, J. Emergence of virulent methicillin-resistant staphylococcus aureus strains carrying panton-valentine leucocidin genes in the Netherlands. J. Clin. Microbiol. 2005, 43, 3341–3345. [Google Scholar] [CrossRef]
- Rasmussen, G.; Monecke, S.; Ehricht, R.; Soderquist, B. Prevalence of clonal complexes and virulence genes among commensal and invasive staphylococcus aureus isolates in Sweden. PLoS ONE 2013, 8, e77477. [Google Scholar]
- von Eiff, C.; Friedrich, A.W.; Peters, G.; Becker, K. Prevalence of genes encoding for members of the staphylococcal leukotoxin family among clinical isolates of staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 2004, 49, 157–162. [Google Scholar] [CrossRef]
- Baba-Moussa, L.; Sina, H.; Scheftel, J.M.; Moreau, B.; Sainte-Marie, D.; Kotchoni, S.O.; Prevost, G.; Couppie, P. Staphylococcal panton-valentine leucocidin as a major virulence factor associated to furuncles. PLoS ONE 2011, 6, e25716. [Google Scholar] [CrossRef]
- Gravet, A.; Couppie, P.; Meunier, O.; Clyti, E.; Moreau, B.; Pradinaud, R.; Monteil, H.; Prevost, G. Staphylococcus aureus isolated in cases of impetigo produces both epidermolysin A or B and luke-lukd in 78% of 131 Retrospective and Prospective Cases. J. Clin. Microbiol. 2001, 39, 4349–4356. [Google Scholar] [CrossRef]
- Gravet, A.; Rondeau, M.; Harf-Monteil, C.; Grunenberger, F.; Monteil, H.; Scheftel, J.M.; Prevost, G. Predominant staphylococcus aureus isolated from antibiotic-associated diarrhea is clinically relevant and produces enterotoxin a and the bicomponent toxin luke-lukd. J. Clin. Microbiol. 1999, 37, 4012–4019. [Google Scholar]
- Thomsen, I.P.; Dumont, A.L.; James, D.B.; Yoong, P.; Saville, B.R.; Soper, N.; Torres, V.J.; Creech, C.B. Children with invasive staphylococcus aureus disease exhibit a potently neutralizing antibody response to the cytotoxin lukab. Infect. Immun. 2014, 82, 1234–1242. [Google Scholar] [CrossRef]
- Alonzo, F., 3rd; Benson, M.A.; Chen, J.; Novick, R.P.; Shopsin, B.; Torres, V.J. Staphylococcus aureus leucocidin Ed contributes to systemic infection by targeting neutrophils and promoting bacterial growth in Vivo. Mol. Microbiol. 2012, 83, 423–435. [Google Scholar] [CrossRef]
- Dumont, A.L.; Nygaard, T.K.; Watkins, R.L.; Smith, A.; Kozhaya, L.; Kreiswirth, B.N.; Shopsin, B.; Unutmaz, D.; Voyich, J.M.; Torres, V.J. Characterization of a new cytotoxin that contributes to staphylococcus aureus pathogenesis. Mol. Microbiol. 2011, 79, 814–825. [Google Scholar] [CrossRef]
- Joubert, O.; Viero, G.; Keller, D.; Martinez, E.; Colin, D.A.; Monteil, H.; Mourey, L.; Dalla Serra, M.; Prevost, G. Engineered covalent leucotoxin heterodimers form functional pores: Insights into S-F interactions. Biochem. J. 2006, 396, 381–389. [Google Scholar] [CrossRef]
- Malachowa, N.; Whitney, A.R.; Kobayashi, S.D.; Sturdevant, D.E.; Kennedy, A.D.; Braughton, K.R.; Shabb, D.W.; Diep, B.A.; Chambers, H.F.; Otto, M.; et al. Global changes in staphylococcus aureus gene expression in human blood. PLoS ONE 2011, 6, e18617. [Google Scholar] [CrossRef]
- Malachowa, N.; Deleo, F.R. Staphylococcus aureus survival in human blood. Virulence 2011, 2, 567–569. [Google Scholar] [CrossRef]
- Hongo, I.; Baba, T.; Oishi, K.; Morimoto, Y.; Ito, T.; Hiramatsu, K. Phenol-soluble modulin alpha 3 enhances the human neutrophil lysis mediated by panton-valentine leukocidin. J. Infect. Dis. 2009, 200, 715–723. [Google Scholar] [CrossRef]
- Loffler, B.; Hussain, M.; Grundmeier, M.; Bruck, M.; Holzinger, D.; Varga, G.; Roth, J.; Kahl, B.C.; Proctor, R.A.; Peters, G. Staphylococcus aureus panton-valentine leukocidin is a very potent cytotoxic factor for human neutrophils. PLoS Pathog. 2010, 6, e1000715. [Google Scholar] [CrossRef]
- Zhang, D.; Shooshtarizadeh, P.; Laventie, B.J.; Colin, D.A.; Chich, J.F.; Vidic, J.; de Barry, J.; Chasserot-Golaz, S.; Delalande, F.; Van Dorsselaer, A.; et al. Two chromogranin a-derived peptides induce calcium entry in human neutrophils by calmodulin-regulated calcium independent phospholipase A2. PLoS ONE 2009, 4, e4501. [Google Scholar] [CrossRef]
- RReyes-Robles, T.; Alonzo, F., 3rd; Kozhaya, L.; Lacy, D.B.; Unutmaz, D.; Torres, V.J. Staphylococcus Aureus leukotoxin Ed targets the chemokine receptors Cxcr1 and Cxcr2 to kill leukocytes and promote infection. Cell Host Microbe 2013, 14, 453–459. [Google Scholar] [CrossRef]
- Aslam, R.; Laventie, B.J.; Marban, C.; Prevost, G.; Keller, D.; Strub, J.M.; Dorsselaer, A.; Haikel, Y.; Taddei, C.; Metz-Boutigue, M.H. Activation of neutrophils by the two-component leukotoxin luke/d from staphylococcus aureus: proteomic analysis of the secretions. J. Proteome. Res. 2013, 12, 3667–3678. [Google Scholar] [CrossRef]
- Staali, L.; Monteil, H.; Colin, D.A. The staphylococcal pore-forming leukotoxins open Ca2+ channels in the membrane of human polymorphonuclear neutrophils. J. Membr. Biol. 1998, 162, 209–216. [Google Scholar] [CrossRef]
- DuMont, A.L.; Yoong, P.; Day, C.J.; Alonzo, F., 3rd; McDonald, W.H.; Jennings, M.P.; Torres, V.J. Staphylococcus aureus lukab cytotoxin kills human neutrophils by targeting the Cd11b subunit of the integrin Mac-1. Proc. Natl. Acad. Sci. USA 2013, 110, 10794–10799. [Google Scholar]
- Malachowa, N.; Kobayashi, S.D.; Braughton, K.R.; Whitney, A.R.; Parnell, M.J.; Gardner, D.J.; Deleo, F.R. Staphylococcus aureus leukotoxin gh promotes inflammation. J. Infect. Dis. 2012, 206, 1185–1193. [Google Scholar] [CrossRef]
- Ventura, C.L.; Malachowa, N.; Hammer, C.H.; Nardone, G.A.; Robinson, M.A.; Kobayashi, S.D.; DeLeo, F.R. Identification of a novel staphylococcus aureus two-component leukotoxin using cell surface proteomics. PLoS ONE 2010, 5, e11634. [Google Scholar] [CrossRef]
- Malachowa, N.; Kobayashi, S.D.; Freedman, B.; Dorward, D.W.; Deleo, F.R. Staphylococcus aureus leukotoxin Gh promotes formation of neutrophil extracellular traps. J. Immunol. 2013, 191, 6022–6029. [Google Scholar] [CrossRef]
- Zivkovic, A.; Sharif, O.; Stich, K.; Doninger, B.; Biaggio, M.; Colinge, J.; Bilban, M.; Mesteri, I.; Hazemi, P.; Lemmens-Gruber, R.; Knapp, S. Tlr 2 and Cd14 mediate innate immunity and lung inflammation to staphylococcal panton-valentine leukocidin in Vivo. J. Immunol. 2011, 186, 1608–1617. [Google Scholar] [CrossRef]
- Perret, M.; Badiou, C.; Lina, G.; Burbaud, S.; Benito, Y.; Bes, M.; Cottin, V.; Couzon, F.; Juruj, C.; Dauwalder, O.; et al. Cross-Talk between staphylococcus aureus leukocidins-intoxicated macrophages and lung epithelial cells triggers chemokine secretion in an inflammasome-dependent manner. Cell. Microbiol. 2012, 14, 1019–1036. [Google Scholar] [CrossRef]
- Fackrell, H.B.; Wiseman, G.M. Properties of the gamma haemolysin of staphylococcus aureus “Smith 5r”. J. Gen. Microbiol. 1976, 92, 11–24. [Google Scholar] [CrossRef]
- Yokota, K.; Sugawara, N.; Nariya, H.; Kaneko, J.; Tomita, T.; Kamio, Y. Further study on the two pivotal parts of Hlg2 for the full hemolytic activity of staphylococcal gamma-hemolysin. Biosci. Biotechnol. Biochem. 1998, 62, 1745–1750. [Google Scholar] [CrossRef]
- Gravet, A.; Colin, D.A.; Keller, D.; Girardot, R.; Monteil, H.; Prevost, G. Characterization of a novel structural member, luke-lukd, of the Bi-component staphylococcal leucotoxins family. FEBS Lett. 1998, 436, 202–208. [Google Scholar] [CrossRef]
- Morinaga, N.; Kaihou, Y.; Noda, M. Purification, cloning and characterization of variant luke-lukd with strong leukocidal activity of staphylococcal Bi-component leukotoxin family. Microbiol. Immunol. 2003, 47, 81–90. [Google Scholar] [CrossRef]
- Prevost, G.; Cribier, B.; Couppie, P.; Petiau, P.; Supersac, G.; Finck-Barbancon, V.; Monteil, H.; Piemont, Y. Panton-Valentine leucocidin and gamma-hemolysin from staphylococcus aureus Atcc 49775 are encoded by distinct genetic loci and have different biological activities. Infect. Immun. 1995, 63, 4121–4129. [Google Scholar]
- Dalla Serra, M.; Coraiola, M.; Viero, G.; Comai, M.; Potrich, C.; Ferreras, M.; Baba-Moussa, L.; Colin, D.A.; Menestrina, G.; Bhakdi, S.; et al. Staphylococcus aureus bicomponent gamma-hemolysins, Hlga, Hlgb, and Hlgc, can form mixed pores containing all components. J. Chem. Inf. Model. 2005, 45, 1539–1545. [Google Scholar] [CrossRef]
- Sugawara, N.; Tomita, T.; Kamio, Y. Assembly of staphylococcus aureus gamma-hemolysin into a pore-forming ring-shaped complex on the surface of human erythrocytes. FEBS Lett. 1997, 410, 333–337. [Google Scholar] [CrossRef]
- Siqueira, J.A.; Speeg-Schatz, C.; Freitas, F.I.; Sahel, J.; Monteil, H.; Prevost, G. Channel-Forming leucotoxins from staphylococcus aureus cause severe inflammatory reactions in a rabbit eye model. J. Med. Microbiol. 1997, 46, 486–494. [Google Scholar] [CrossRef]
- Gao, H.; Neff, T.A.; Guo, R.F.; Speyer, C.L.; Sarma, J.V.; Tomlins, S.; Man, Y.; Riedemann, N.C.; Hoesel, L.M.; Younkin, E.; et al. Evidence for a functional role of the second C5a receptor C5l2. FASEB J. 2005, 19, 1003–1005. [Google Scholar]
- Woodruff, T.M.; Nandakumar, K.S.; Tedesco, F. Inhibiting the C5-C5a receptor axis. Mol. Immunol. 2011, 48, 1631–1642. [Google Scholar] [CrossRef]
- Archer, N.K.; Harro, J.M.; Shirtliff, M.E. Clearance of staphylococcus aureus nasal carriage is T cell dependent and mediated through interleukin-17a expression and neutrophil influx. Infect. Immun. 2013, 81, 2070–2075. [Google Scholar] [CrossRef]
- Frank, K.M.; Zhou, T.; Moreno-Vinasco, L.; Hollett, B.; Garcia, J.G.; Bubeck Wardenburg, J. Host response signature to staphylococcus aureus alpha-hemolysin implicates pulmonary Th17 response. Infect. Immun. 2012, 80, 3161–3169. [Google Scholar] [CrossRef]
- Ma, C.S.; Chew, G.Y.; Simpson, N.; Priyadarshi, A.; Wong, M.; Grimbacher, B.; Fulcher, D.A.; Tangye, S.G.; Cook, M.C. Deficiency of Th17 cells in hyper ige syndrome due to mutations in stat3. J. Exp. Med. 2008, 205, 1551–1557. [Google Scholar] [CrossRef]
- Spellberg, B.; Daum, R. Development of a vaccine against staphylococcus aureus. Semin. Immunopathol. 2012, 34, 335–348. [Google Scholar] [CrossRef]
- Dumont, A.L.; Yoong, P.; Liu, X.; Day, C.J.; Chumbler, N.M.; James, D.B.; Alonzo III, F.; Bode, N.J.; Lacy, D.B.; Jennings, M.P.; et al. Identification of a Crucial Residue Required for Staphylococcus Aureus Lukab Cytotoxicity and Receptor Recognition. Infect. Immun. 2013. [Google Scholar] [CrossRef]
- Kaneko, J.; Ozawa, T.; Tomita, T.; Kamio, Y. Sequential binding of staphylococcal gamma-hemolysin to human erythrocytes and complex formation of the hemolysin on the cell surface. Biosci. Biotechnol. Biochem. 1997, 61, 846–851. [Google Scholar] [CrossRef]
- Bogomolski-Yahalom, V.; Matzner, Y. Disorders of neutrophil function. Blood Rev. 1995, 9, 183–190. [Google Scholar] [CrossRef]
- Kretschmer, D.; Gleske, A.K.; Rautenberg, M.; Wang, R.; Koberle, M.; Bohn, E.; Schoneberg, T.; Rabiet, M.J.; Boulay, F.; Klebanoff, S.J.; et al. Human formyl peptide receptor 2 senses highly pathogenic staphylococcus aureus. Cell Host Microbe 2010, 7, 463–473. [Google Scholar] [CrossRef]
- Berube, B.J.; Bubeck Wardenburg, J. Staphylococcus aureus alpha-toxin: Nearly a century of intrigue. Toxins 2013, 5, 1140–1166. [Google Scholar] [CrossRef]
- de Haas, C.J.; Veldkamp, K.E.; Peschel, A.; Weerkamp, F.; Van Wamel, W.J.; Heezius, E.C.; Poppelier, M.J.; Van Kessel, K.P.; van Strijp, J.A. Chemotaxis inhibitory protein of staphylococcus aureus, a bacterial antiinflammatory agent. J. Exp. Med. 2004, 199, 687–695. [Google Scholar] [CrossRef]
- Graves, S.F.; Kobayashi, S.D.; Braughton, K.R.; Whitney, A.R.; Sturdevant, D.E.; Rasmussen, D.L.; Kirpotina, L.N.; Quinn, M.T.; DeLeo, F.R. Sublytic concentrations of staphylococcus aureus panton-valentine leukocidin alter human pmn gene expression and enhance bactericidal capacity. J. Leukoc. Biol. 2012, 92, 361–374. [Google Scholar] [CrossRef]
- Vojtov, N.; Ross, H.F.; Novick, R.P. Global repression of exotoxin synthesis by staphylococcal superantigens. Proc. Natl. Acad. Sci. USA 2002, 99, 10102–10107. [Google Scholar] [CrossRef]
- Pilsczek, F.H.; Salina, D.; Poon, K.K.; Fahey, C.; Yipp, B.G.; Sibley, C.D.; Robbins, S.M.; Green, F.H.; Surette, M.G.; Sugai, M.; et al. A novel mechanism of rapid nuclear neutrophil extracellular trap formation in response to staphylococcus aureus. J. Immunol. 2010, 185, 7413–7425. [Google Scholar] [CrossRef]
- Stuart, L.M.; Ezekowitz, R.A. Phagocytosis: Elegant complexity. Immunity 2005, 22, 539–550. [Google Scholar] [CrossRef]
- Tseng, C.W.; Kyme, P.; Low, J.; Rocha, M.A.; Alsabeh, R.; Miller, L.G.; Otto, M.; Arditi, M.; Diep, B.A.; Nizet, V.; et al. Staphylococcus aureus panton-valentine leukocidin contributes to inflammation and muscle tissue injury. PLoS ONE 2009, 4, e6387. [Google Scholar] [CrossRef]
- Kebaier, C.; Chamberland, R.R.; Allen, I.C.; Gao, X.; Broglie, P.M.; Hall, J.D.; Jania, C.; Doerschuk, C.M.; Tilley, S.L.; Duncan, J.A. Staphylococcus aureus alpha-hemolysin mediates virulence in a murine model of severe pneumonia through activation of the Nlrp3 inflammasome. J. Infect. Dis. 2012, 205, 807–817. [Google Scholar] [CrossRef]
- Powers, M.E.; Kim, H.K.; Wang, Y.; Bubeck Wardenburg, J. Adam10 mediates vascular injury induced by staphylococcus aureus alpha-hemolysin. J. Infect. Dis. 2012, 206, 352–356. [Google Scholar] [CrossRef]
- Adhikari, R.P.; Ajao, A.O.; Aman, M.J.; Karauzum, H.; Sarwar, J.; Lydecker, A.D.; Johnson, J.K.; Nguyen, C.; Chen, W.H.; Roghmann, M.C. Lower antibody levels to staphylococcus aureus exotoxins are associated with sepsis in hospitalized adults with invasive S. aureus infections. J. Infect. Dis. 2012, 206, 915–923. [Google Scholar] [CrossRef]
- Fritz, S.A.; Tiemann, K.M.; Hogan, P.G.; Epplin, E.K.; Rodriguez, M.; Al-Zubeidi, D.N.; Bubeck Wardenburg, J.; Hunstad, D.A. A serologic correlate of protective immunity against community-onset staphylococcus aureus infection. Clin. Infect. Dis. 2013, 56, 1154–1161. [Google Scholar]
- Hermos, C.R.; Yoong, P.; Pier, G.B. High levels of antibody to panton-valentine leukocidin are not associated with resistance to staphylococcus aureus-associated skin and soft-tissue infection. Clin. Infect. Dis. 2010, 51, 1138–1146. [Google Scholar] [CrossRef]
- Laventie, B.J.; Rademaker, H.J.; Saleh, M.; de Boer, E.; Janssens, R.; Bourcier, T.; Subilia, A.; Marcellin, L.; van Haperen, R.; Lebbink, J.H.; et al. Heavy chain-only antibodies and tetravalent bispecific antibody neutralizing staphylococcus aureus leukotoxins. Proc. Natl. Acad. Sci. USA 2011, 108, 16404–16409. [Google Scholar] [CrossRef]
- Foletti, D.; Strop, P.; Shaughnessy, L.; Hasa-Moreno, A.; Casas, M.G.; Russell, M.; Bee, C.; Wu, S.; Pham, A.; Zeng, Z.; et al. Mechanism of action and in Vivo Efficacy of a human-derived antibody against staphylococcus aureus alpha-hemolysin. J. Mol. Biol. 2013, 425, 1641–1654. [Google Scholar] [CrossRef]
- Ragle, B.E.; Bubeck Wardenburg, J. Anti-alpha-hemolysin monoclonal antibodies mediate protection against staphylococcus aureus pneumonia. Infect. Immun. 2009, 77, 2712–2718. [Google Scholar] [CrossRef]
- Tkaczyk, C.; Hua, L.; Varkey, R.; Shi, Y.; Dettinger, L.; Woods, R.; Barnes, A.; MacGill, R.S.; Wilson, S.; Chowdhury, P.; et al. Identification of anti-alpha toxin monoclonal antibodies that reduce the severity of staphylococcus aureus dermonecrosis and exhibit a correlation between affinity and potency. Clin. Vaccine Immunol. 2012, 19, 377–385. [Google Scholar] [CrossRef]
- Varshney, A.K.; Wang, X.; Scharff, M.D.; Macintyre, J.; Zollner, R.S.; Kovalenko, O.V.; Martinez, L.R.; Byrne, F.R.; Fries, B.C. Staphylococcal enterotoxin b-specific monoclonal antibody 20b1 successfully treats diverse staphylococcus aureus infections. J. Infect. Dis. 2013. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Aman, M.J.; Adhikari, R.P. Staphylococcal Bicomponent Pore-Forming Toxins: Targets for Prophylaxis and Immunotherapy. Toxins 2014, 6, 950-972. https://doi.org/10.3390/toxins6030950
Aman MJ, Adhikari RP. Staphylococcal Bicomponent Pore-Forming Toxins: Targets for Prophylaxis and Immunotherapy. Toxins. 2014; 6(3):950-972. https://doi.org/10.3390/toxins6030950
Chicago/Turabian StyleAman, M. Javad, and Rajan P. Adhikari. 2014. "Staphylococcal Bicomponent Pore-Forming Toxins: Targets for Prophylaxis and Immunotherapy" Toxins 6, no. 3: 950-972. https://doi.org/10.3390/toxins6030950