In Vitro Toxic Effects of Puff Adder (Bitis arietans) Venom, and Their Neutralization by Antivenom

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
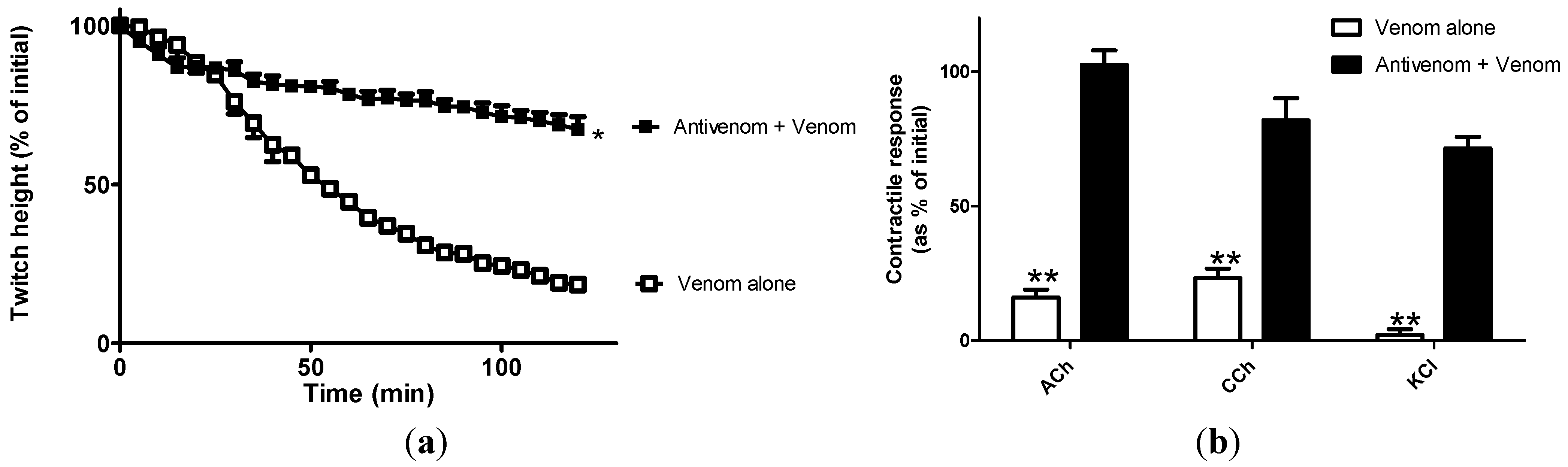
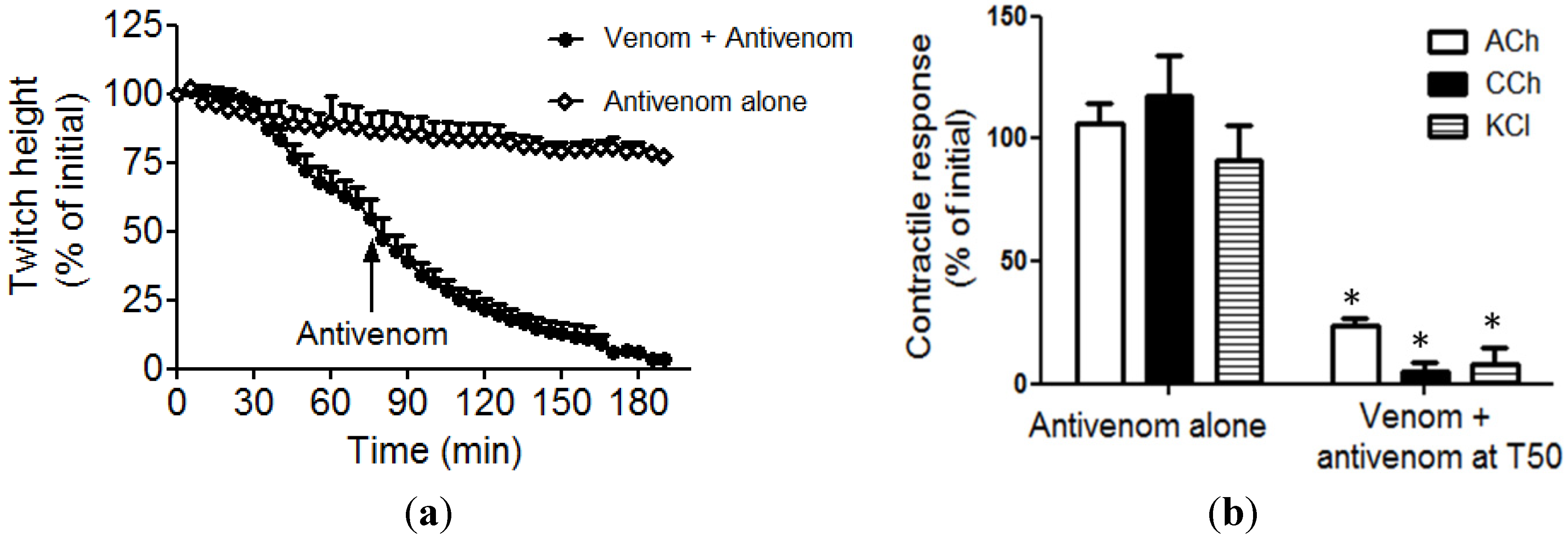
2.1. Neurotoxicity


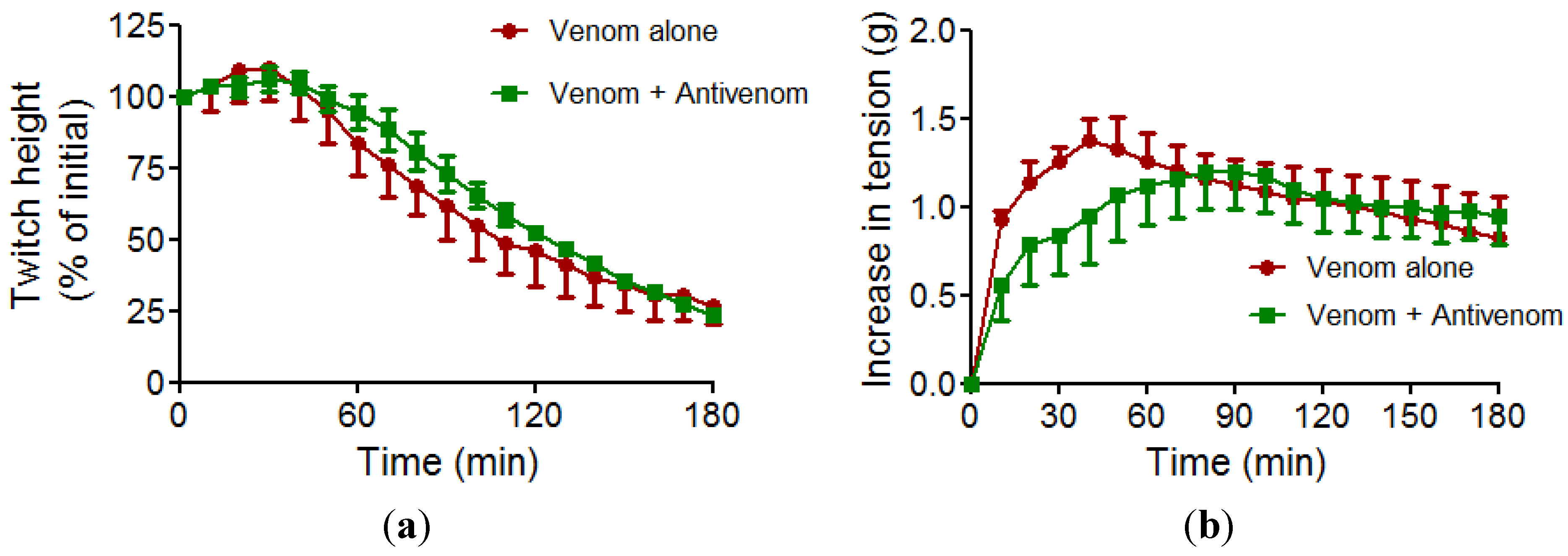
2.2. Myotoxicity

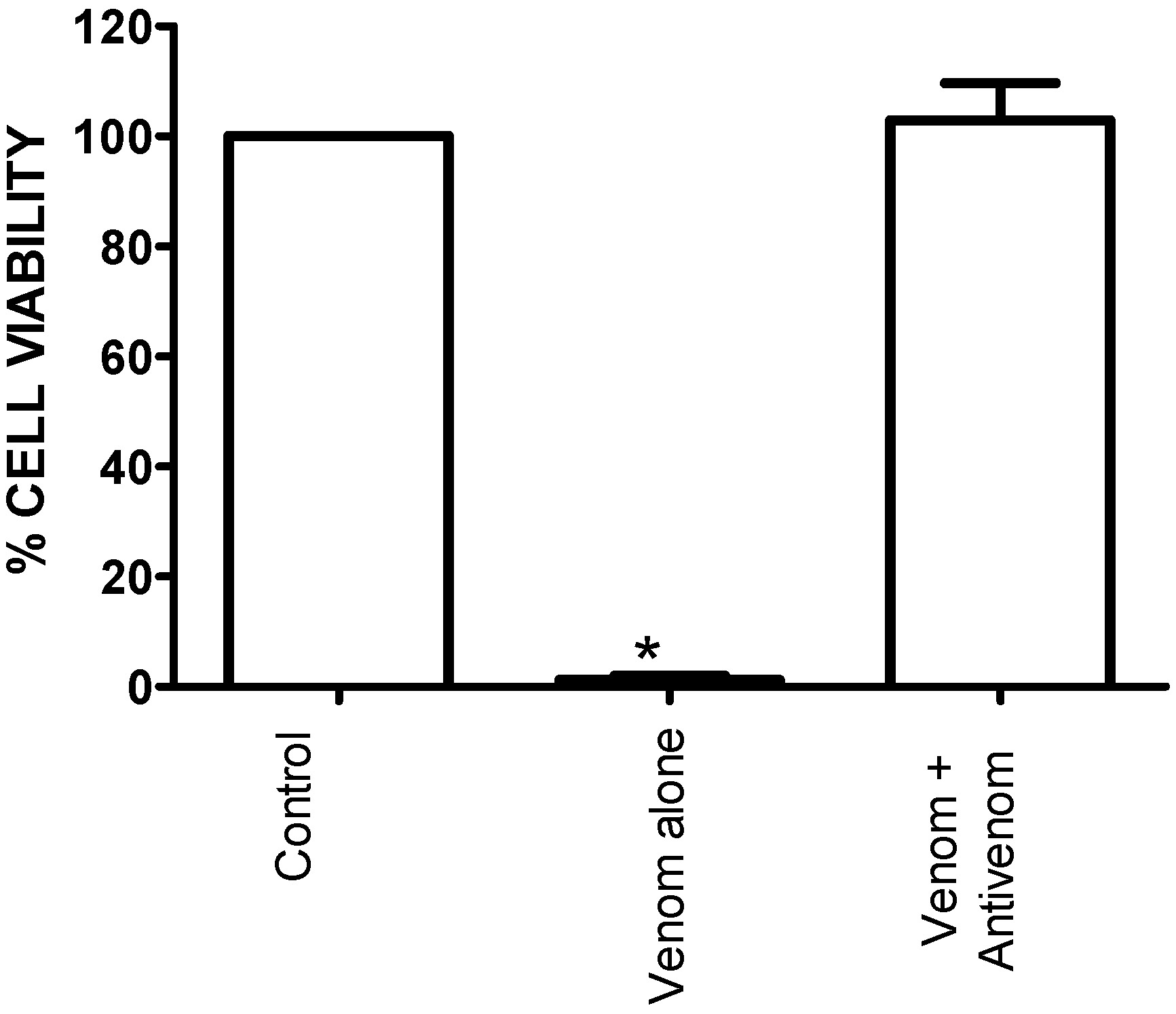
2.3. Effect of Venom on Cell Viability

2.4. 2D Gel Electrophoresis and Western Immunoblotting

2.5. Coagulation Studies
3. Experimental Section
3.1. Drugs and Chemicals
3.2. Venoms and Antivenoms
3.3. Neurotoxicity Studies
3.4. Myotoxicity Studies
3.5. Cell Culture
3.6. Cell Proliferation Assay (MTS Assay)
3.6.1. Effect of Venom
3.6.2. Antivenom Effects
3.7. 2D Gel Electrophoresis and Western Immunoblotting
3.7.1. Iso-Electric Focusing
3.7.2. SDS-PAGE
3.7.3. Western Immunoblotting
3.8. Coagulation Studies
3.8.1. Effect of Venom
3.8.2. Antivenom Effects
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author contributions
Conflicts of Interest
References
- Barlow, A.; Baker, K.; Hendry, C.R.; Peppin, L.; Phelps, T.; Tolley, K.A.; Wuster, C.E.; Wuster, W. Phylogeography of the widespread African puff adder (Bitis arietans) reveals multiple Pleistocene refugia in southern Africa. Mol. Ecol. 2013, 22, 1134–1157. [Google Scholar] [CrossRef]
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5. [Google Scholar] [CrossRef]
- Warrell, D.A.; Ormerod, L.D.; Davidson, N.M. Bites by puff-adder (Bitis arietans) in Nigeria, and value of antivenom. Br. Med. J. 1975, 4, 697–700. [Google Scholar] [CrossRef]
- Lavonas, E.J.; Tomaszewski, C.A.; Ford, M.D.; Rouse, A.M.; Kerns, W.P., 2nd. Severe puff adder (Bitis arietans) envenomation with coagulopathy. J. Toxicol. Clin. Toxicol. 2002, 40, 911–918. [Google Scholar] [CrossRef]
- Vulfius, C.A.; Gorbacheva, E.V.; Starkov, V.G.; Osipov, A.V.; Kasheverov, I.E.; Andreeva, T.V.; Astashev, M.E.; Tsetlin, V.I.; Utkin, Y.N. An unusual phospholipase A(2) from puff adder Bitis arietans venom—A novel blocker of nicotinic acetylcholine receptors. Toxicon 2011, 57, 787–793. [Google Scholar] [CrossRef]
- Matsui, T.; Hamako, J.; Matsushita, T.; Nakayama, T.; Fujimura, Y.; Titani, K. Binding site on human von Willebrand factor of bitiscetin, a snake venom-derived platelet aggregation inducer. Biochemistry 2002, 41, 7939–7946. [Google Scholar] [CrossRef]
- Jennings, B.R.; Spearman, C.W.; Kirsch, R.E.; Shephard, E.G. A novel high molecular weight fibrinogenase from the venom of Bitis arietans. Biochim. Biophys. Acta 1999, 1427, 82–91. [Google Scholar]
- Cook, D.A.; Owen, T.; Wagstaff, S.C.; Kinne, J.; Wernery, U.; Harrison, R.A. Analysis of camelid antibodies for antivenom development: Neutralisation of venom-induced pathology. Toxicon 2010, 56, 373–380. [Google Scholar] [CrossRef]
- Kalam, Y.; Isbister, G.K.; Mirtschin, P.; Hodgson, W.C.; Konstantakopoulos, N. Validation of a cell-based assay to differentiate between the cytotoxic effects of elapid snake venoms. J. Pharmacol. Toxicol. methods 2011, 63, 137–142. [Google Scholar] [CrossRef]
- Kuruppu, S.; Smith, A.I.; Isbister, G.K.; Hodgson, W.C. Neurotoxins from Australo-Papuan elapids: A biochemical and pharmacological perspective. Crit. Rev. Toxicol. 2008, 38, 73–86. [Google Scholar] [CrossRef]
- Barber, C.M.; Isbister, G.K.; Hodgson, W.C. Solving the “Brown snake paradox”: In vitro characterisation of Australasian snake presynaptic neurotoxin activity. Toxicol. Letters 2012, 210, 318–323. [Google Scholar] [CrossRef]
- Currie, B.J. Snakebite in tropical Australia, Papua New Guinea and Irian Jaya. Emerg. Med. 2000, 12, 285–294. [Google Scholar] [CrossRef]
- Wickramaratna, J.C.; Hodgson, W.C. A pharmacological examination of venoms from three species of death adder (Acanthophis antarcticus, Acanthophis praelongus and Acanthophis pyrrhus). Toxicon 2001, 39, 209–216. [Google Scholar] [CrossRef]
- Crachi, M.T.; Hammer, L.W.; Hodgson, W.C. The effects of antivenom on the in vitro neurotoxicity of venoms from the taipans Oxyuranus scutellatus, Oxyuranus microlepidotus and Oxyuranus scutellatus canni. Toxicon 1999, 37, 1771–1778. [Google Scholar] [CrossRef]
- Ponte, C.G.; Nobrega, E.L.; Fernandes, V.C.; da Silva, W.D.; Suarez-Kurtz, G. Inhibition of the myotoxic activities of three African Bitis venoms (B. rhinoceros, B. arietans and B. nasicornis) by a polyvalent antivenom. Toxicon 2010, 55, 536–540. [Google Scholar] [CrossRef]
- Mendez, I.; Gutierrez, J.M.; Angulo, Y.; Calvete, J.J.; Lomonte, B. Comparative study of the cytolytic activity of snake venoms from African spitting cobras (Naja spp., Elapidae) and its neutralization by a polyspecific antivenom. Toxicon 2011, 58, 558–564. [Google Scholar] [CrossRef]
- Warrell, D. Clinical Features of Envenoming from Snake Bites. In Envenomings and Their Treatments; Bon, C., Goyffon, M., Eds.; Foundation Marcel Merieux: Lyon, France, 1996; pp. 63–76. [Google Scholar]
- O’Leary, M.A.; Isbister, G.K. A turbidimetric assay for the measurement of clotting times of procoagulant venoms in plasma. J. Pharmacol. Toxicol. Methods 2010, 61, 27–31. [Google Scholar] [CrossRef]
- Isbister, G.K.; Woods, D.; Alley, S.; O’Leary, M.A.; Seldon, M.; Lincz, L.F. Endogenous thrombin potential as a novel method for the characterization of procoagulant snake venoms and the efficacy of antivenom. Toxicon 2010, 56, 75–85. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fernandez, S.; Hodgson, W.; Chaisakul, J.; Kornhauser, R.; Konstantakopoulos, N.; Smith, A.I.; Kuruppu, S. In Vitro Toxic Effects of Puff Adder (Bitis arietans) Venom, and Their Neutralization by Antivenom. Toxins 2014, 6, 1586-1597. https://doi.org/10.3390/toxins6051586
Fernandez S, Hodgson W, Chaisakul J, Kornhauser R, Konstantakopoulos N, Smith AI, Kuruppu S. In Vitro Toxic Effects of Puff Adder (Bitis arietans) Venom, and Their Neutralization by Antivenom. Toxins. 2014; 6(5):1586-1597. https://doi.org/10.3390/toxins6051586
Chicago/Turabian StyleFernandez, Steven, Wayne Hodgson, Janeyuth Chaisakul, Rachelle Kornhauser, Nicki Konstantakopoulos, Alexander Ian Smith, and Sanjaya Kuruppu. 2014. "In Vitro Toxic Effects of Puff Adder (Bitis arietans) Venom, and Their Neutralization by Antivenom" Toxins 6, no. 5: 1586-1597. https://doi.org/10.3390/toxins6051586