Impact of Nitrogen Sources on Gene Expression and Toxin Production in the Diazotroph Cylindrospermopsis raciborskii CS-505 and Non-Diazotroph Raphidiopsis brookii D9


Abstract
:1. Introduction
2. Results
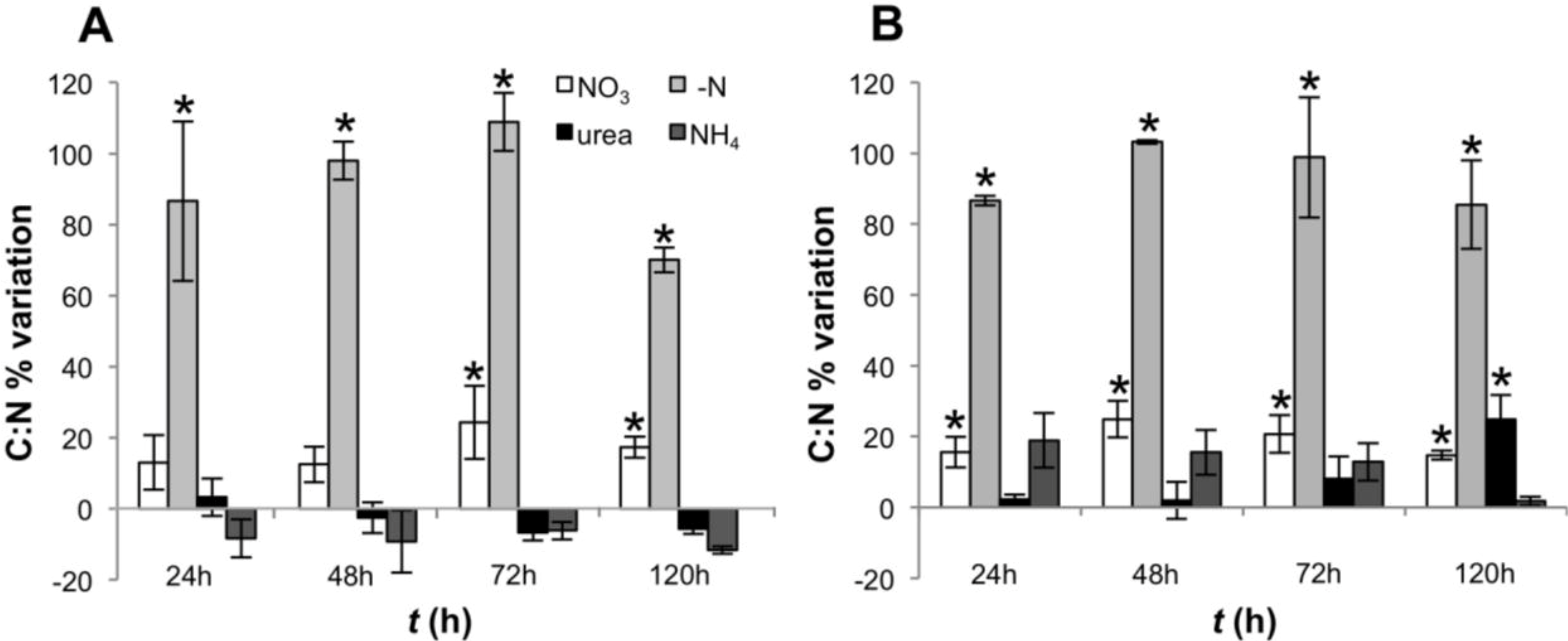
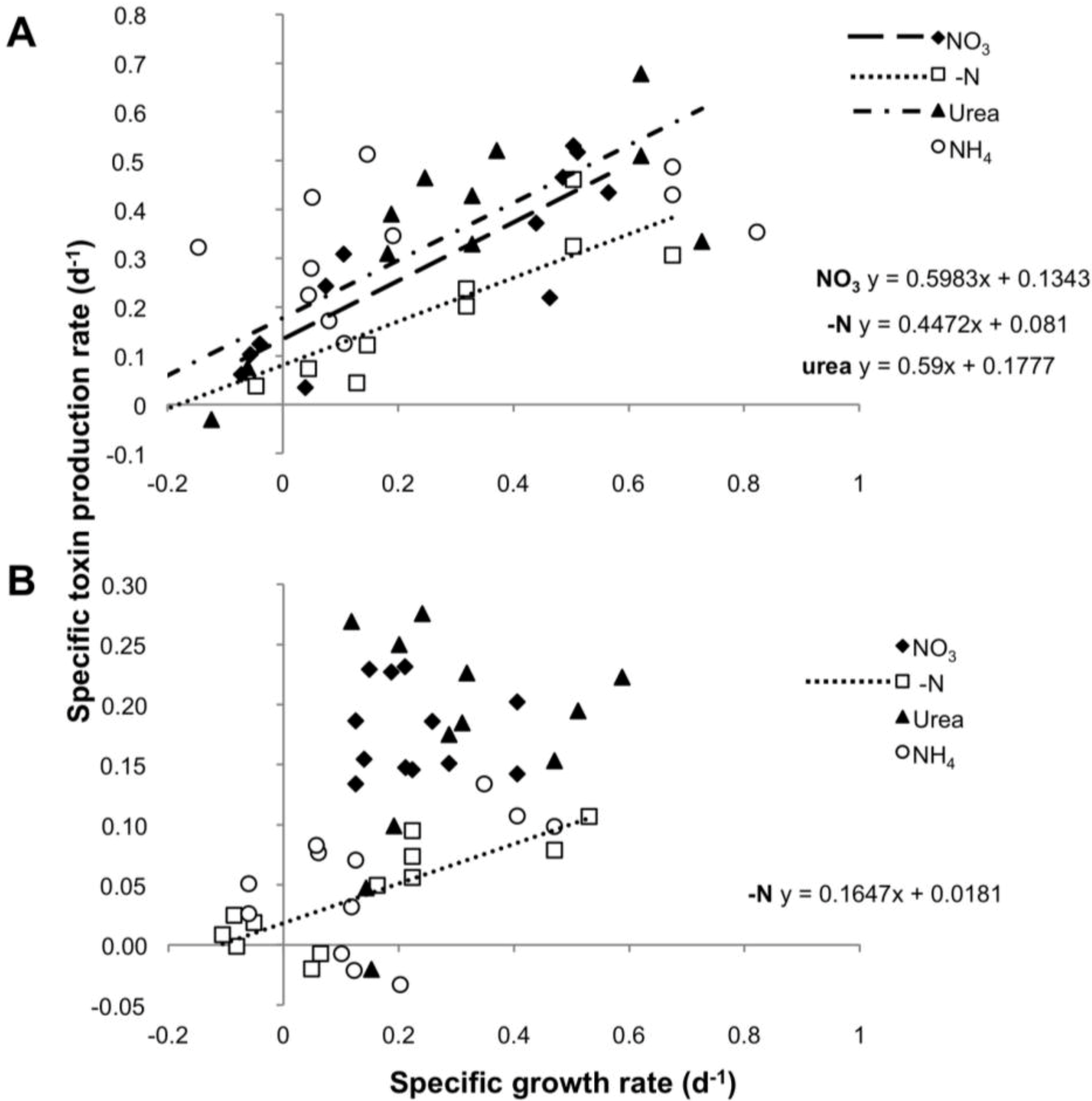
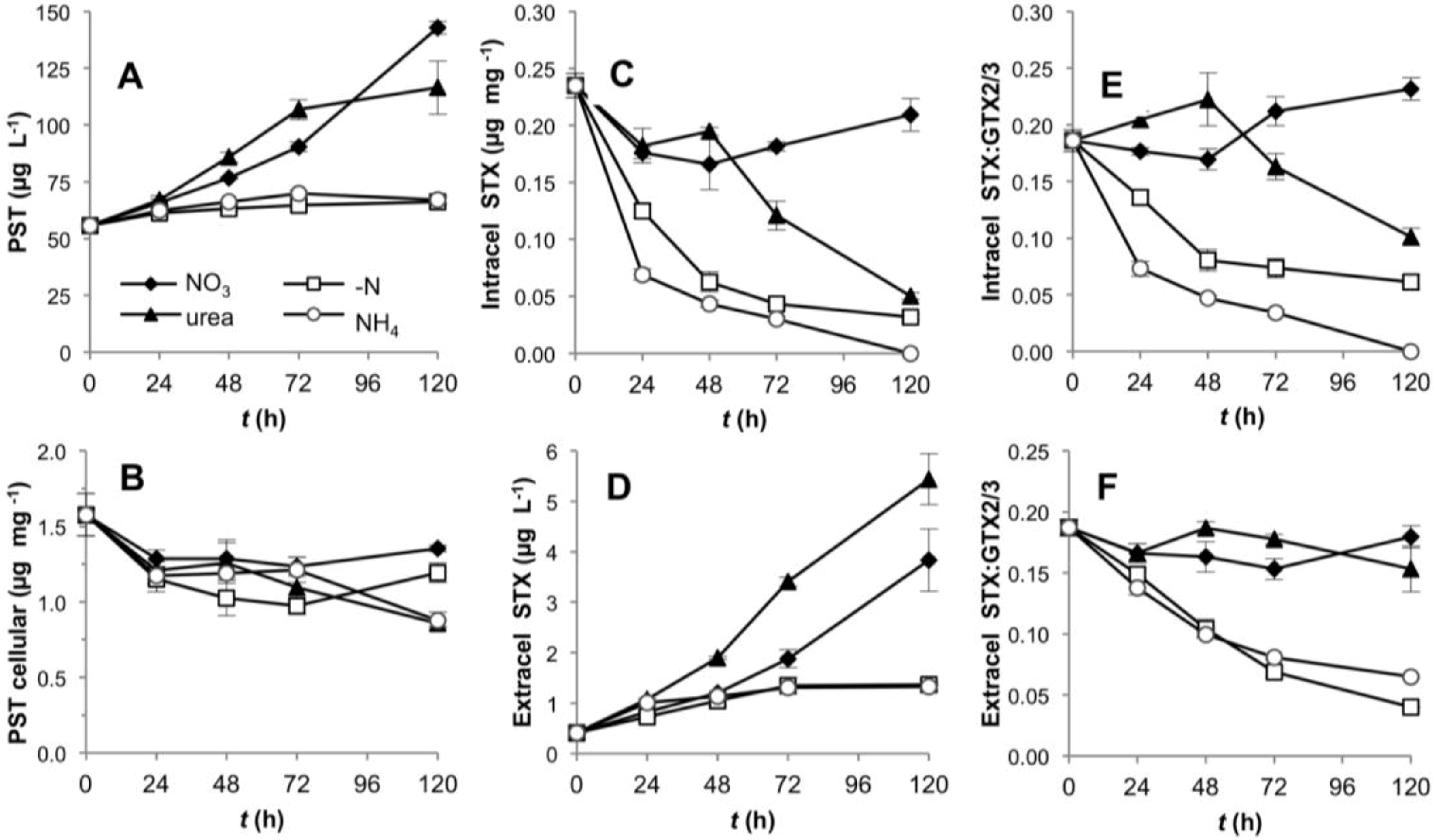
2.1. Growth and Toxin Production under Different N-Regimes





2.2. NtcA Binding Boxes and Transcriptional Regulation of cyr and sxt Genes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Predicted Function/Reference | NtcA Motif | Time (h) | % Transcript Relative to Time 0 | |||
|---|---|---|---|---|---|---|
| NO3 | NH4 | urea | -N | |||
| ntcA (CS-505) Transcriptional regulator in nitrogen metabolism [37] | Y | 24 | 60.3 ± 3.9 | 89 ± 7.5 | 132.3 ± 15.4 | 110.5 ± 12.7 |
| 48 | 135.9 ± 22 | 139.1 ± 4.8 | 220.7 ± 20 | 283.1 ± 17.6 | ||
| 72 | 107.2 ± 16.3 | 80.6 ± 12.8 | NM | 155.9 ± 14.9 | ||
| 120 | 136.2 ± 20.1 | 47.8 ± 9.1 | NM | 181.8 ± 7.6 | ||
| ntcA (D9) Transcriptional regulator in nitrogen metabolism [37] | Y | 24 | 63.5 ± 6.8 | 98.7 ± 8.7 | 87.8 ± 13.3 | NM |
| 48 | 54.2 ± 9.5 | 56.3 ± 2.8 | 111 ± 15.7 | NM | ||
| 72 | 16.6 ± 5.8 | 27.4 ± 1.3 | NM | NM | ||
| 120 | 49 ± 1 | 30.5 ± 5.1 | NM | NM | ||
| cyrB NRPS/PKS, second step in CYN biosynthesis [8] | Y | 24 | 62.8 ± 1.7 | 93 ± 21 | 121.9 ± 25.2 | 35.1 ± 9 |
| 48 | 107.4 ± 4.7 | 141.8 ± 15.9 | 124.2 ± 58.7 | 75.3 ± 7.4 | ||
| 72 | 126.3 ± 3.2 | 73 ± 4.6 | NM | 64.1 ± 8.9 | ||
| 120 | 20.1 ± 5 | 29.8 ± 8.9 | NM | 65.4 ± 2.1 | ||
| cyrI Hydroxylation of C-7 in doCYN to form CYN [8,38] | N | 24 | 73.8 ± 8.8 | 80 ±7.8 | 121.9 ± 25.2 | 35.1 ± 9 |
| 48 | 96.4 ± 18.6 | 141.8 ± 15.9 | 124.2 ± 58.7 | 75.3 ± 7.4 | ||
| 72 | 110.3 ± 11.7 | 73 ± 4.6 | NM | 64.1 ± 8.9 | ||
| 120 | 66.7 ± 16 | 29.8 ± 8.9 | NM | 65.4 ± 2.1 | ||
| cyrJ Sulfotransferase [8] | Y | 24 | 74.4 ± 6.1 | 74.2 ± 3.3 | 106.2 ± 18.5 | 66.4 ± 10.2 |
| 48 | 107.2 ± 1.6 | 114.9 ± 1.7 | 179 ± 21 | 89.9 ± 12.3 | ||
| 72 | 43.5 ± 3 | 76.7 ± 6.6 | NM | 60.7 ± 17.3 | ||
| 120 | 54 ± 9.5 | 31.9 ± 9.8 | NM | 64.1 ± 9.4 | ||
| cyrK Transport of CYN/doCYN [8] | Y | 24 | 93.2 ± 4.6 | 68.3 ± 5.6 | 52.4 ± 1.8 | 106.3 ± 15.9 |
| 48 | 101 ± 0.4 | 96.9 ± 13.5 | 63.6 ±12.2 | 114.9 ± 10 | ||
| 72 | 44.5 ± 5.9 | 60.9 ± 1.7 | NM | 86.6 ± 11.6 | ||
| 120 | 38.1 ± 7.3 | 29.2 ± 9.9 | NM | 96.7 ± 13.4 | ||
| sxtSUL Sulfotransferase rendering GTX2/3 [39] | N | 24 | 38.5 ± 8.4 | 160.2 ± 5.7 | 61.3 ± 13 | NM |
| 48 | 36.2 ± 9.4 | 131.4 ± 21.7 | 98.4 ± 31.4 | NM | ||
| 72 | 20.5 ± 1.2 | 47.2 ± 4.8 | NM | NM | ||
| 120 | 19.5 ± 3 | 46.2 ± 6 | NM | NM | ||
| sxtDIOX Hydroxylation of STX prior to GTX2/3 formation [39] | N | 24 | 85.8 ± 7.6 | 116.1 ± 25 | 57 ± 3.2 | NM |
| 48 | 146 ± 11.4 | 104.9 ± 20.3 | 78 ± 7 | NM | ||
| 72 | 74.5 ± 6.4 | 51.8 ± 4.7 | NM | NM | ||
| 120 | 71.6 ± 1.3 | 75.5 ± 1.2 | NM | NM | ||
| sxtM PST export [11] | Y | 24 | 83.8 ± 11.7 | 161.3 ± 18.2 | 69.2 ± 6.4 | NM |
| 48 | 60.8 ± 2.8 | 141 ± 25.5 | 122.5 ± 25 | NM | ||
| 72 | 45.11 ± 4.4 | 71.9 ± 5.1 | NM | NM | ||
| 120 | 48.1 ± 3.9 | 80 ± 6.6 | NM | NM | ||
| sxtF PST export [11] | N | 24 | 84.5 ± 13 | 152.3 ± 14.3 | 86.8 ± 2.9 | NM |
| 48 | 64.9 ± 3.6 | 83 ± 14.1 | 86.9 ± 5.5 | NM | ||
| 72 | 52.4 ± 3 | 72.6 ± 11 | NM | NM | ||
| 120 | 65.9 ± 6.2 | 70.5 ± 6 | NM | NM | ||
| sxtA ACP/aminotransferase, first step in STX biosynthesis [11] | Y | 24 | 116.2 ± 10.5 | 165.8 ± 10.6 | 124 ± 3.7 | NM |
| 48 | 77.5 ± 6.5 | 97 ± 10.4 | 160.7 ± 26 | NM | ||
| 72 | 67.6 ± 2 | 36.6 ± 2.2 | NM | NM | ||
| 120 | 58.2 ± 4.3 | 30.6 ± 1.2 | NM | NM | ||
| sxtO Donor of sulfate group [11] | N | 24 | 90.4 ± 3.8 | 60.3 ± 0.8 | 36 ± 6.9 | NM |
| 48 | 60.6 ± 3.9 | 51.4 ± 4.2 | 57.5 ± 8.5 | NM | ||
| 72 | 30.1 ± 2.9 | 22 ± 3.6 | NM | NM | ||
| 120 | 35.6 ± 7.8 | 15.3 ± 1.5 | NM | NM | ||
| sxtI Carbamoyltransferase [11] | N | 24 | 47.3 ± 4.2 | 179.1 ± 8.8 | 86.5 ± 8.1 | NM |
| 48 | 51.9 ± 3.1 | 106.9 ± 10.7 | 104.9 ± 9.5 | NM | ||
| 72 | 35.5 ± 4.1 | 59.1 ± 5.7 | NM | NM | ||
| 120 | 39.5 ± 5.7 | 49.1 ± 8.4 | NM | NM | ||
| sxtU Reduction of C-1, eighth step in STX biosynthesis [11] | N | 24 | 61.8 ± 8.2 | 125.8 ± 17.3 | 91.4 ± 13.5 | NM |
| 48 | 62.2 ± 2.8 | 123.4 ± 21.2 | 132.3 ± 13.4 | NM | ||
| 72 | 40.2 ± 0.3 | 82.1 ± 5.6 | NM | NM | ||
| 120 | 37.6 ± 4.8 | 53.2 ± 1.7 | NM | NM | ||
3. Discussion
4. Experimental Section
4.1. Cultures and Experimental Settings
4.2. Analysis of Toxins
4.3. Statistical Analysis
4.4. RNA Isolation
4.5. qRT-PCR Conditions and Motif Search
5. Conclusions
Supplementary Files
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Herrero, A.; Muro-Pastor, A.M.; Flores, E. Nitrogen control in cyanobacteria. J. Bacteriol. 2001, 183, 411–425. [Google Scholar] [CrossRef]
- Carmichael, W.W. The cyanotoxins. Adv. Bot. Res. 1997, 27, 211–256. [Google Scholar] [CrossRef]
- Sivonen, K.; Jones, G. Cyanobacterial toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences,Monitoring and Management; Chrorus, I., Bartam, J., Eds.; E & FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- Tillett, D.; Dittmann, E.; Erhard, M.; Döhren, von, H.; Börner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef]
- Edwards, D.J.; Gerwick, W.H. Lyngbyatoxin biosynthesis: Sequence of biosynthetic gene cluster and identification of a novel aromatic prenyltransferase. J. Am. Chem. Soc. 2004, 126, 11432–11433. [Google Scholar] [CrossRef]
- Moffitt, M.C.; Neilan, B.A. Characterization of the nodularin synthetase gene cluster and proposed theory of the evolution of cyanobacterial hepatotoxins. Appl. Environ. Microbiol. 2004, 70, 6353–6362. [Google Scholar] [CrossRef]
- Méjean, A.; Mann, S.; Maldiney, T.; Vassiliadis, G.; Lequin, O.; Ploux, O. Evidence that biosynthesis of the neurotoxic alkaloids anatoxin-a and homoanatoxin-a in the cyanobacterium Oscillatoria PCC 6506 occurs on a modular polyketide synthase initiated by L-proline. J. Am. Chem. Soc. 2009, 131, 7512–7513. [Google Scholar]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl. Environ. Microbiol. 2008, 74, 716–722. [Google Scholar] [CrossRef]
- Grindberg, R.V.; Ishoey, T.; Brinza, D.; Esquenazi, E.; Coates, R.C.; Liu, W.-T.; Gerwick, L.; Dorrestein, P.C.; Pevzner, P.; Lasken, R.; et al. Single cell genome amplification accelerates identification of the apratoxin biosynthetic pathway from a complex microbial assemblage. PLoS One 2011, 6, e18565. [Google Scholar]
- Hackett, J.D.; Wisecaver, J.H.; Brosnahan, M.L.; Kulis, D.M.; Anderson, D.M.; Bhattacharya, D.; Plumley, F.G.; Erdner, D.L. Evolution of saxitoxin synthesis in cyanobacteria and dinoflagellates. Mol. Biol. Evol. 2013, 30, 70–78. [Google Scholar] [CrossRef]
- Kellmann, R.; Mihali, T.K.; Jeon, Y.J.; Pickford, R.; Pomati, F.; Neilan, B.A. Biosynthetic intermediate analysis and functional homology reveal a saxitoxin gene cluster in cyanobacteria. Appl. Environ. Microbiol. 2008, 74, 4044–4053. [Google Scholar] [CrossRef]
- Reynolds, C.S. The Ecology of Phytoplankton; Cambridge University Press: New York, NY, USA, 2006. [Google Scholar]
- Orr, P.T.; Jones, G.J. Relationship between microcystin production and cell division rates in nitrogen-limited Microcystis aeruginosa cultures. Limnol. Oceanogr. 1998, 43, 1604–1614. [Google Scholar] [CrossRef]
- Long, B.M.; Jones, G.J.; Orr, P.T. Cellular microcystin content in N-limited Microcystis aeruginosa can be predicted from growth rate. Appl. Environ. Microbiol. 2001, 67, 278–283. [Google Scholar]
- Sevilla, E.; Martin-Luna, B.; Vela, L.; Teresa Bes, M.; Luisa Peleato, M.; Fillat, M. Microcystin-LR synthesis as response to nitrogen: Transcriptional analysis of the mcyD gene in Microcystis aeruginosa PCC7806. Ecotoxicology 2010, 19, 1167–1173. [Google Scholar] [CrossRef]
- Tonk, L.; van De Waal, D.B.; Slot, P.; Huisman, J.; Matthijs, H.C.P.; Visser, P.M. Amino acid availability determines the ratio of microcystin variants in the cyanobacterium Planktothrix agardhii. FEMS Microbiol. Ecol. 2008, 65, 383–390. [Google Scholar] [CrossRef]
- Paerl, H. Coastal eutrophication and harmful algal blooms: Importance of atmospheric deposition andgroundwater as “new” nitrogen and other nutrient sources. Limnol. Oceanogr. 1997, 42, 1154–1165. [Google Scholar] [CrossRef]
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Progress in understanding harmful algal blooms: Paradigm shifts and new technologies for research, monitoring, and management. Annu. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef]
- Leong, S.C.Y.; Murata, A.; Nagashima, Y.; Taguchi, S. Variability in toxicity of the dinoflagellate Alexandrium tamarense in response to different nitrogen sources and concentrations. Toxicon 2004, 43, 407–415. [Google Scholar] [CrossRef]
- Cembella, A.D. Ecophysiology and metabolism of paralytic shellfish toxins in marine microalgae. In Physiological Ecology of Harmful Algal Blooms; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer Verlag: Heidelberg, Germany, 1998; pp. 381–403. [Google Scholar]
- Dias, E.; Pereira, P.; Franca, S. Production of paralytic shellfish toxins by Aphanizomenon sp. LMECYA 31 (cyanobacteria). J. Phycol. 2002, 38, 705–712. [Google Scholar] [CrossRef]
- Saker, M.L.; Neilan, B.A. Varied diazotrophies, morphologies, and toxicities of genetically similar isolates of Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) from northern Australia. Appl. Environ. Microbiol. 2001, 67, 1839–1845. [Google Scholar] [CrossRef]
- Shalev-Malul, G.; Lieman-Hurwitz, J.; Viner-Mozzini, Y.; Sukenik, A.; Gaathon, A.; Lebendiker, M.; Kaplan, A. An AbrB-like protein might be involved in the regulation of cylindrospermopsin production by Aphanizomenon ovalisporum. Environ. Microbiol. 2008, 10, 988–999. [Google Scholar] [CrossRef]
- Bormans, M.; Lengronne, M.; Brient, L.; Duval, C. Cylindrospermopsin accumulation and release by the benthic cyanobacterium Oscillatoria sp. PCC 6506 under different light conditions and growth phases. Bull. Environ. Contam. Toxicol. 2014, 92, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Cirés, S.; Wörmer, L.; Timón, J.; Wiedner, C.; Quesada, A. Cylindrospermopsin production and release by the potentially invasive cyanobacterium Aphanizomenon ovalisporum under temperature and light gradients. Harmful Algae 2011, 10, 668–675. [Google Scholar] [CrossRef]
- Preußel, K.; Wessel, G.; Fastner, J.; Chorus, I. Response of cylindrospermopsin production and release in Aphanizomenon flos-aquae (cyanobacteria) to varying light and temperature conditions. Harmful Algae 2009, 8, 645–650. [Google Scholar] [CrossRef]
- Bar-Yosef, Y.; Sukenik, A.; Hadas, O.; Viner-Mozzini, Y.; Kaplan, A. Enslavement in the water body by toxic Aphanizomenon ovalisporum, inducing alkaline phosphatase in phytoplanktons. Curr. Biol. 2010, 20, 1557–1561. [Google Scholar] [CrossRef]
- Saker, M.L.; Griffiths, D.J. The effect of temperature on growth and cylindrospermopsin content of seven isolates of Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) from water bodies in northern Australia. Phycologia 2000, 39, 349–354. [Google Scholar] [CrossRef]
- Bácsi, I.; Vasas, G.; Surányi, G.; M-Hamvas, M.; Máthé, C.; Tóth, E.; Grigorszky, I.; Gáspár, A.; Tóth, S.; Borbely, G. Alteration of cylindrospermopsin production in sulfate- or phosphate-starved cyanobacterium Aphanizomenon ovalisporum. FEMS Microbiol. Lett. 2006, 259, 303–310. [Google Scholar] [CrossRef]
- Saker, M.L.; Neilan, B.A.; Griffiths, D.J. Two morphological forms of Cylindrospermopsis raciborskii (cyanobacteria) isolated from Solomon dam, Palm Island, Queensland. J. Phycol. 1999, 35, 599–606. [Google Scholar]
- Soto, K.; Stucken, K.; Méndez, M.A.; Lagos, N.; Cembella, A.D.; Krock, B.; Vásquez, M. The Effects of Chloramphenicol,Arginine and Temperature on PST-production by Cylindrospermopsis raciborskii Strain D9. In Proceedings of the 12th International Conference on Harmful Algae, Copenhagen, Denmark, 4–8 September 2006; Moestrup, O., Ed.; International Society for the Study of Harmful Algae and Intergovernmental Oceanographic Commission of UNESCO: Copenhagen, Denmark, 2008; pp. 330–333. [Google Scholar]
- Stucken, K.; John, U.; Cembella, A.; Murillo, A.A.; Soto-Liebe, K.; Fuentes-Valdés, J.J.; Friedel, M.; Plominsky, Á.M.; Vásquez, M.; Glöckner, G. The smallest known genomes of multicellular and toxic cyanobacteria: Comparison, minimal gene sets for linked traits and the evolutionary implications. PLoS One 2010, 5, e9235. [Google Scholar]
- Stucken, K. Physiogenomics of Cylindrospermopsis raciborskii/Raphidiopsis brookii: With Emphasis on cyanobacterial Evolution,Nitrogen Control and Toxin Biosynthesis; LAP Lambert Academic Publishing: Saarbrücken, Germany, 2010. [Google Scholar]
- Herrero, A.; Muro-Pastor, A.M.; Valladares, A.; Flores, E. Cellular differentiation and the NtcA transcription factor in filamentous cyanobacteria. FEMS Microbiol. Rev. 2004, 28, 469–487. [Google Scholar] [CrossRef]
- Su, Z.; Olman, V.; Mao, F.; Xu, Y. Comparative genomics analysis of NtcA regulons in cyanobacteria: regulation of nitrogen assimilation and its coupling to photosynthesis. Nucleic Acids Res. 2005, 33, 5156–5171. [Google Scholar] [CrossRef]
- Luque, I.; Flores, E.; Herrero, A. Molecular mechanism for the operation of nitrogen control in cyanobacteria. EMBO J. 1994, 13, 2682–2689. [Google Scholar]
- Vega-Palas, M.A.; Madueño, F.; Herrero, A.; Flores, E. Identification and cloning of a regulatory gene for nitrogen assimilation in the cyanobacterium Synechococcus sp. strain PCC 7942. J. Bacteriol. 1990, 172, 643–647. [Google Scholar]
- Mazmouz, R.; Chapuis-Hugon, F.; Pichon, V.; Méjean, A.; Ploux, O. The last step of the biosynthesis of the cyanotoxins cylindrospermopsin and 7-epi-cylindrospermopsin is catalysed by CyrI, a 2-oxoglutarate-dependent iron oxygenase. ChemBioChem 2011, 12, 858–862. [Google Scholar] [CrossRef]
- Soto-Liebe, K.; Méndez, M.A.; Fuenzalida, L.; Krock, B.; Cembella, A.; Vásquez, M. PSP toxin release from the cyanobacterium Raphidiopsis brookii D9 (Nostocales) can be induced by sodium and potassium ions. Toxicon 2012, 60, 1324–1334. [Google Scholar] [CrossRef]
- Plominsky, Á.M.; Larsson, J.; Bergman, B.; Delherbe, N.; Osses, I.; Vásquez, M. Dinitrogen fixation is restricted to the terminal heterocysts in the invasive cyanobacterium Cylindrospermopsis raciborskii CS-505. PLoS One 2013, 8, e51682. [Google Scholar]
- Dyble, J.; Tester, P.A.; Litaker, A.R.W. Effects of light intensity on cylindrospermopsin production in the cyanobacterial HAB species Cylindrospermopsis raciborskii. Afr.J. Mar. Sci. 2006, 28, 309–312. [Google Scholar] [CrossRef]
- Davis, T.W.; Orr, P.T.; Boyer, G.L.; Burford, M.A. Investigating the production and release of cylindrospermopsin and deoxy-cylindrospermopsin by Cylindrospermopsis raciborskii over a natural growth cycle. Harmful Algae 2014, 31, 18–25. [Google Scholar] [CrossRef]
- Yunes, J.S.; La Rocha De, S.; Giroldo, D.; Silveira, S.B.D.; Comin, R.; Bicho, M.D.S.; Melcher, S.S.; Sant’anna, C.L.; Vieira, A.A.H. Release of carbohydrates and proteins by a subtropical strain of Raphidiopsis brookii (cyanobacteria) able to produce saxitoxin at three nitrate concentrations. J. Phycol. 2009, 45, 585–591. [Google Scholar] [CrossRef]
- Hawkins, P.R.; Putt, E.; Falconer, I.; Humpage, A. Phenotypical variation in a toxic strain of the phytoplankter, Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) during batch culture. Environ. Toxicol. 2001, 16, 460–467. [Google Scholar] [CrossRef]
- Orr, P.T.; Rasmussen, J.P.; Burford, M.A.; Eaglesham, G.K.; Lennox, S.M. Evaluation of quantitative real-time PCR to characterise spatial and temporal variations in cyanobacteria, Cylindrospermopsis raciborskii (Woloszynska) Seenaya et Subba Raju and cylindrospermopsin concentrations in three subtropical Australian reservoirs. Harmful Algae 2010, 9, 243–254. [Google Scholar] [CrossRef]
- Vázquez-Bermúdez, M.F.; Herrero, A.; Flores, E. 2-oxoglutarate increases the binding affinity of the NtcA (nitrogen control) transcription factor for the Synechococcus glnA promoter. FEBS Lett. 2002, 512, 71–74. [Google Scholar] [CrossRef]
- Muro-Pastor, M.I.; Reyes, J.C.; Florencio, F.J. Cyanobacteria perceive nitrogen status by sensing intracellular 2-oxoglutarate levels. J. Biol. Chem. 2001, 276, 38320–38328. [Google Scholar]
- Jiang, Y.; Xiao, P.; Yu, G.; Sano, T.; Pan, Q.; Li, R. Molecular basis and phylogenetic implications of deoxycylindrospermopsin biosynthesis in the cyanobacterium Raphidiopsis curvata. Appl. Environ. Microbiol. 2012, 78, 2256–2263. [Google Scholar] [CrossRef]
- Pomati, F.; Burns, B.P.; Neilan, B.A. Use of ion-channel modulating agents to study cyanobacterial Na+-K+ fluxes. Biol. Res. 2004, 6, 137–143. [Google Scholar]
- Van De Waal, D.B.; Verspagen, J.M.H.; Lürling, M.; van Donk, E.; Visser, P.M.; Huisman, J. The ecological stoichiometry of toxins produced by harmful cyanobacteria: An experimental test of the carbon-nutrient balance hypothesis. Ecol. Lett. 2009, 12, 1326–1335. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B.; Kokociński, M.; Jurczak, T.; Lipski, D.; Wiktorowicz, K. Interspecific allelopathy in cyanobacteria: cylindrospermopsin and Cylindrospermopsis raciborskii effect on the growth and metabolism of Microcystis aeruginosa. Harmful Algae 2014, 35, 1–8. [Google Scholar] [CrossRef]
- Isvánovics, V.; Shafik, H.M.; Présing, M.; Juhos, S. Growth and phosphate uptake kinetics of the cyanobacterium, Cylindrospermopsis raciborskii (Cyanophyceae) in throughflow cultures. Freshwater Biol. 2000, 43, 257–275. [Google Scholar] [CrossRef]
- Castro, D.; Vera, D.; Lagos, N.; García, C.; Vásquez, M. The effect of temperature on growth and production of paralytic shellfish poisoning toxins by the cyanobacterium Cylindrospermopsis raciborskii C10. Toxicon 2004, 44, 483–489. [Google Scholar] [CrossRef]
- Lawton, L.; Marsalek, B.; Padisák, J.; Chorus, I. Determination of cyanobacteria in the laboratory. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences,Monitoring and Management; Chorus, I., Bartram, J., Eds.; E&FN Spon: London, UK, 1999; pp. 1–28. [Google Scholar]
- Tillmann, U.; Alpermann, T.L.; da Purificação, R.C.; Krock, B.; Cembella, A. Intra-population clonal variability in allelochemical potency of the toxigenic dinoflagellate Alexandrium tamarense. Harmful Algae 2009, 8, 759–769. [Google Scholar] [CrossRef]
- Anderson, D.M.; Kulis, D.M.; Sullivan, J.J.; Hall, S.; Lee, C. Dynamics and physiology of saxitoxin production by the dinoflagellates Alexandrium spp. Mar. Biol. 1990, 104, 511–524. [Google Scholar] [CrossRef]
- Whelan, J.A.; Russell, N.B.; Whelan, M.A. A method for the absolute quantification of cDNA using real-time PCR. J. Immunol. Methods 2003, 278, 261–269. [Google Scholar] [CrossRef]
- Services in computational biology, Pattern search. Available online: http://nbc11.biologie.uni-kl.de (accessed on 13 June 2014).
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stucken, K.; John, U.; Cembella, A.; Soto-Liebe, K.; Vásquez, M. Impact of Nitrogen Sources on Gene Expression and Toxin Production in the Diazotroph Cylindrospermopsis raciborskii CS-505 and Non-Diazotroph Raphidiopsis brookii D9. Toxins 2014, 6, 1896-1915. https://doi.org/10.3390/toxins6061896
Stucken K, John U, Cembella A, Soto-Liebe K, Vásquez M. Impact of Nitrogen Sources on Gene Expression and Toxin Production in the Diazotroph Cylindrospermopsis raciborskii CS-505 and Non-Diazotroph Raphidiopsis brookii D9. Toxins. 2014; 6(6):1896-1915. https://doi.org/10.3390/toxins6061896
Chicago/Turabian StyleStucken, Karina, Uwe John, Allan Cembella, Katia Soto-Liebe, and Mónica Vásquez. 2014. "Impact of Nitrogen Sources on Gene Expression and Toxin Production in the Diazotroph Cylindrospermopsis raciborskii CS-505 and Non-Diazotroph Raphidiopsis brookii D9" Toxins 6, no. 6: 1896-1915. https://doi.org/10.3390/toxins6061896