Identification and Characterization of the Insecticidal Toxin “Makes Caterpillars Floppy” in Photorhabdus temperata M1021 Using a Cosmid Library


Abstract
:1. Introduction
2. Results and Discussion
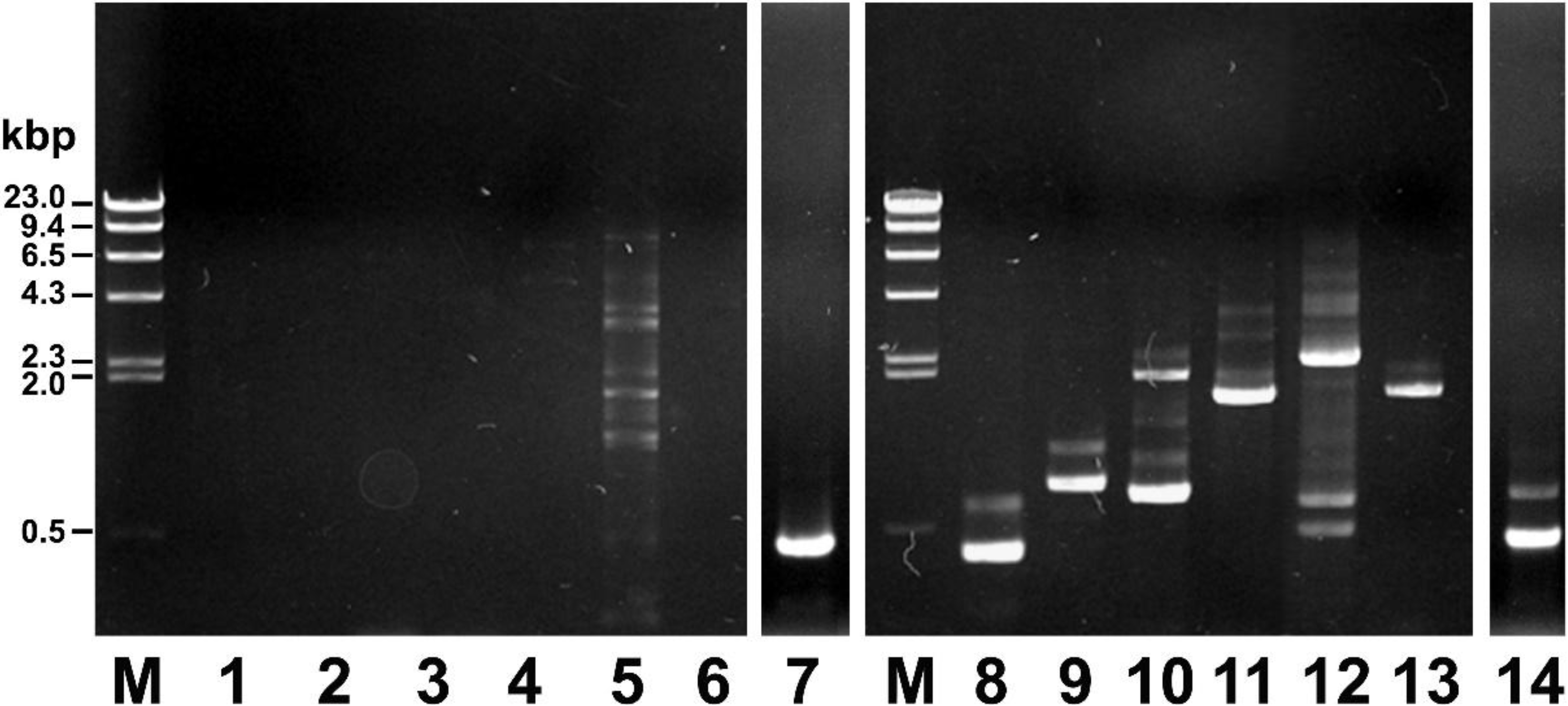
2.1. Screening of the Cosmid Library

2.2. Sequence Alignment and Bioinformatics Analysis of the mcf Gene

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Names of the Target Genes | Primers | Primer Sequence (5'→3') |
|---|---|---|
| Partial tccB gene (0.46 kb) | TccB1 TccB2 | GTG ACT CAG CTT TCA ATC GCC AT GAT AGA TCC GCC ACC ATA TCC AG |
| Partial tcc gene locus (0.84 kb) | Tcc1 Tcc2 | AAR YTG GCT GAA GAG CAY GTG CAA TAC CCT TAC CTG TGC |
| Partial tcd gene locus (0.77 kb) | Tcd1 Tcd2 | AAG ACC GTT TTT CCC GTT ATG AAT A ATC ACC GGA TTG CAC CAC ATG |
| Partial tca gene locus (1.6 kb) | Tca1 Tca2 | GGA ATT CAT GCT GAG CTA TTG GCA A GGT CGA CGC GAA TGG TAT AGC GAA T |
| Partial tcdA gene locus (2.2 kb) | TcdA1 TcdA2 | ACC GAT GGA TTT CAG TGG CGC T GCG TCG ACT TGG CGA ATG GTG TAG CGA A |
| Partial chitinase gene (1.6 kb) | ChiA1 ChiA2 | GCG AAT TCC ATA TGT TTA AAA CAA TTG TAT CG CTG AAA GTA CAG GTT CTC ATT TAA TTT GCA CGC TTC ACA |
| Partial mcf gene locus (0.87 kb) | Mcf1 Mcf2 | CCT GAC GCC GTT GCC CGA TGA CAC AAC AGG GCA CCG GAT TCA GAG ATG |



2.3. Insecticidal Bioassay of the Recombinant Clones against Insect Larvae

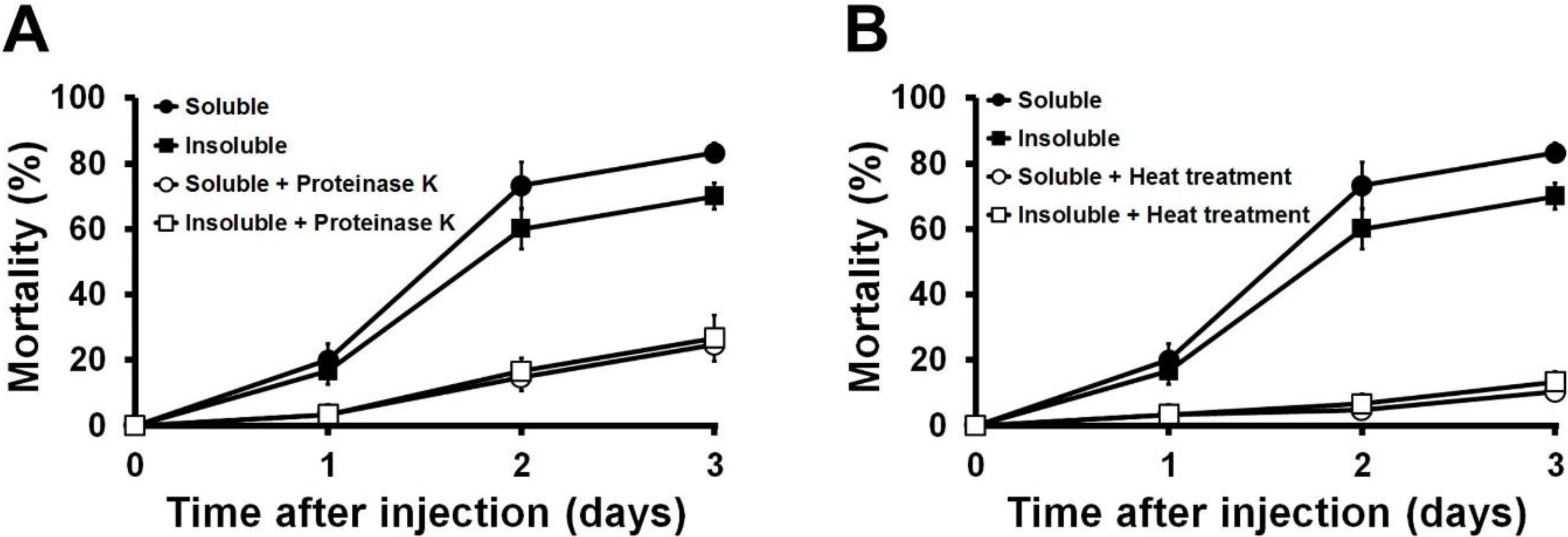
2.4. Effects of Proteinase K and Heat Treatments on the Insecticidal Activity of the Toxin Protein

3. Experimental Section
3.1. Bacterial Culture and Isolation of Genomic DNA
3.2. Construction of the Cosmid Library
3.3. Breeding of Larvae
3.4. Toxicity Assays of the Recombinant Clones against the Model Larvae
3.5. Sequence-Based Analysis of the Selected Cosmid Clone
3.6. Effects of Heat Treatment and Proteinase K Digestion on the Toxin Protein
4. Conclusions
Supplementary Files
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Akhurst, R.J.; Boemare, N.E. A numerical taxonomic study of the genus Xenorhabdus (enterobacteriaceae) and proposed elevation of the subspecies of X. nematophilus to species. J. Gen. Microbiol. 1988, 134, 1835–1845. [Google Scholar]
- Wilkinson, P.; Waterfield, N.R.; Crossman, L.; Corton, C.; Sanchez-Contreras, M.; Vlisidou, I.; Barron, A.; Bignell, A.; Clark, L.; Ormond, D.; et al. Comparative genomics of the emerging human pathogen Photorhabdus asymbiotica with the insect pathogen Photorhabdus luminescens. BMC Genomics 2009, 10, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Park, G.S.; Khan, A.R.; Hong, S.J.; Jang, E.K.; Ullah, I.; Jung, B.K.; Choi, J.; Yoo, N.K.; Park, K.J.; Shin, J.H. Draft genome sequence of entomopathogenic bacterium Photorhabdus temperata strain M1021, isolated from nematodes. Genome Announc. 2013, 1, 1–2. [Google Scholar]
- Duchaud, E.; Rusniok, C.; Frangeul, L.; Buchrieser, C.; Givaudan, A.; Taourit, S.; Bocs, S.; Boursaux-Eude, C.; Chandler, M.; Charles, J.F.; et al. The genome sequence of the entomopathogenic bacterium Photorhabdus luminescens. Nat. Biotechnol. 2003, 21, 1307–1313. [Google Scholar]
- Rodou, A.; Ankrah, D.O.; Stathopoulos, C. Toxins and secretion systems of Photorhabdus luminescens. Toxins 2010, 2, 1250–1264. [Google Scholar] [CrossRef]
- Forst, S.; Dowds, B.; Boemare, N.; Stackebrandt, E. Xenorhabdus and Photorhabdus spp.: Bugs that kill bugs. Annu. Rev. Microbiol. 1997, 51, 47–72. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Dowling, A.; Waterfield, N.R. Insecticidal toxins from Photorhabdus bacteria and their potential use in agriculture. Toxicon 2006, 49, 436–451. [Google Scholar] [CrossRef]
- Forst, S.; Nealson, K. Molecular biology of the symbiotic-pathogenic bacteria Xenorhabdus spp. and Photorhabdus spp. Microbiol. Rev. 1996, 60, 21–43. [Google Scholar]
- Jallouli, W.; Zouari, N.; Jaoua, S. Involvement of oxidative stress and growth at high cell density in the viable but nonculturable state of Photorhabdus temperata ssp. temperata strain K122. Process Biochem. 2010, 45, 706–713. [Google Scholar] [CrossRef]
- Jallouli, W.; Hammami, W.; Zouari, N.; Jaoua, S. Medium optimization for biomass production and morphology variance overcome of Photorhabdus temperata ssp. temperata strain K122. Process Biochem. 2008, 43, 1338–1344. [Google Scholar] [CrossRef]
- Daborn, P.J.; Waterfield, N.; Silva, C.P.; Au, C.P.; Sharma, S.; ffrench-Constant, R.H. A single Photorhabdus gene, makes caterpillars floppy (mcf), allows Escherichia coli to persist within and kill insects. Proc. Natl. Acad. Sci. USA 2002, 99, 10742–10747. [Google Scholar]
- Hofmann, F.; Busch, C.; Prepens, U.; Just, I.; Aktories, K. Localization of the glucosyltransferase activity of Clostridium difficile toxin B to the N-terminal part of the holotoxin. J. Biol. Chem. 1997, 272, 11074–11078. [Google Scholar] [CrossRef]
- Khandelwal, P.; Bhatnagar, N.B. Insecticidal activity associated with the outer membrane vesicles of Xenorhabdus nematophilus. Appl. Environ. Microbiol. 2003, 69, 2032–2037. [Google Scholar] [CrossRef]
- Lanois, A.; Pages, S.; Bourot, S.; Canoy, A.S.; Givaudan, A.; Gaudriault, S. Transcriptional analysis of a Photorhabdus sp. Variant reveals transcriptional control of phenotypic variation and multifactorial pathogenicity in insects. Appl. Environ. Microbiol. 2011, 77, 1009–1020. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.; Waterfield, N.; Daborn, P.; Joyce, S.; Bennett, H.; Au, C.; Dowling, A.; Boundy, S.; Reynolds, S.; Clarke, D. Photorhabdus: Towards a functional genomic analysis of a symbiont and pathogen. FEMS Microbiol. Rev. 2003, 26, 433–456. [Google Scholar] [CrossRef]
- Schnepf, E.; Crickmore, N.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar]
- Ehlers, R.-U.; Niemann, I. Molecular identification of Photorhabdus luminescens strains by amplification of specific fragments of the 16s ribosomal DNA. Syst. Appl. Microbiol. 1998, 21, 509–519. [Google Scholar] [CrossRef]
- Fischer-Le Saux, M.; Viallard, V.; Brunel, B.; Normand, P.; Boemare, N.E. Polyphasic classification of the genus Photorhabdus and proposal of new taxa: P. luminescens subsp. luminescens subsp. nov., P. Luminescens subsp. akhurstii subsp. nov., P. luminescens subsp. laumondii subsp. nov., P. temperata sp. nov., P. temperata subsp. temperata subsp. nov. and P. asymbiotica sp. nov. Int. J. Syst. Bacteriol. 1999, 49, 1645–1656. [Google Scholar]
- Blackburn, M.; Golubeva, E.; Bowen, D.; Ffrench-Constant, R.H. A novel insecticidal toxin from Photorhabdus luminescens, toxin complex a (Tca), and its histopathological effects on the midgut of Manduca sexta. Appl. Environ. Microbiol. 1998, 64, 3036–3041. [Google Scholar]
- Yao, Q.; Cui, J.; Zhu, Y.; Wang, G.; Hu, L.; Long, C.; Cao, R.; Liu, X.; Huang, N.; Chen, S.; et al. A bacterial type iii effector family uses the papain-like hydrolytic activity to arrest the host cell cycle. Proc. Natl. Acad. Sci. USA 2009, 106, 3716–3721. [Google Scholar]
- Morgan, J.A.; Sergeant, M.; Ellis, D.; Ousley, M.; Jarrett, P. Sequence analysis of insecticidal genes from Xenorhabdus nematophilus PMFI296. Appl. Environ. Microbiol. 2001, 67, 2062–2069. [Google Scholar]
- Brown, S.E.; Cao, A.T.; Hines, E.R.; Akhurst, R.J.; East, P.D. A novel secreted protein toxin from the insect pathogenic bacterium Xenorhabdus nematophila. J. Biol. Chem. 2004, 279, 14595–14601. [Google Scholar]
- Pullinger, G.D.; Adams, T.E.; Mullan, P.B.; Garrod, T.I.; Lax, A.J. Cloning, expression, and molecular characterization of the dermonecrotic toxin gene of Bordetella spp. Infect. Immun. 1996, 64, 4163–4171. [Google Scholar]
- Ffrench-Constant, R.H.; Waterfield, N.; Burland, V.; Perna, N.T.; Daborn, P.J.; Bowen, D.; Blattner, F.R. A genomic sample sequence of the entomopathogenic bacterium Photorhabdus luminescens W14: Potential implications for virulence. Appl. Environ. Microbiol. 2000, 66, 3310–3329. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov (accessed on 14 March 2014).
- Predict Protein Website. Available online: http://www.predictprotein.org (accessed on 14 March 2014).
- SIB Bioinformatics Resource Portal. Available online: http://www.expasy.org (accessed on 14 March 2014).
- Cheng, E.H.Y.A.; Wei, M.C.; Weiler, S.; Flavell, R.A.; Mak, T.W.; Lindsten, T.; Korsmeyer, S.J. BCL-2, BCL-XL sequester BH3 domain-only molecules preventing bax- and bak-mediated mitochondrial apoptosis. Mol. Cell 2001, 8, 705–711. [Google Scholar]
- Kelekar, A.; Thompson, C.B. Bcl-2-family proteins: The role of the BH3 domain in apoptosis. Trends Cell Biol. 1998, 8, 324–330. [Google Scholar] [CrossRef]
- Sattler, M.; Liang, H.; Nettesheim, D.; Meadows, R.P.; Harlan, J.E.; Eberstadt, M.; Yoon, H.S.; Shuker, S.B.; Chang, B.S.; Minn, A.J.; et al. Structure of Bcl-XL-Bak peptide complex: Recognition between regulators of apoptosis. Science 1997, 275, 983–986. [Google Scholar]
- Just, I.; Selzer, J.; Wilm, M.; von Eichel-Streiber, C.; Mann, M.; Aktories, K. Glucosylation of Rho proteins by Clostridium difficile toxin B. Nature 1995, 375, 500–503. [Google Scholar] [CrossRef]
- Just, I.; Wilm, M.; Selzer, J.; Rex, G.; von Eichel-Streiber, C.; Mann, M.; Aktories, K. The enterotoxin from Clostridium difficile (ToxA) monoglucosylates the rho proteins. J. Biol. Chem. 1995, 270, 13932–13936. [Google Scholar] [CrossRef]
- Czuprynski, C.J.; Welch, R.A. Biological effects of RTX toxins: The possible role of lipopolysaccharide. Trends Microbiol. 1995, 3, 480–483. [Google Scholar]
- Shin, M.K.; Cha, S.B.; Lee, W.J.; Yoo, H.S. Predicting genetic traits and epitope analysis of ApxIVA in Actinobacillus pleuropneumoniae. J. Microbiol. 2011, 49, 462–468. [Google Scholar] [CrossRef]
- Chattopadhyay, A.; Bhatnagar, N.B.; Bhatnagar, R. Bacterial insecticidal toxins. Crit. Rev. Microbiol. 2004, 30, 33–54. [Google Scholar] [CrossRef]
- Prochazkova, K.; Shuvalova, L.A.; Minasov, G.; Voburka, Z.; Anderson, W.F.; Satchell, K.J. Structural and molecular mechanism for autoprocessing of MARTX toxin of Vibrio cholerae at multiple sites. J. Biol. Chem. 2009, 284, 26557–26568. [Google Scholar]
- Linhartova, I.; Bumba, L.; Masin, J.; Basler, M.; Osicka, R.; Kamanova, J.; Prochazkova, K.; Adkins, I.; Hejnova-Holubova, J.; Sadilkova, L.; et al. Rtx proteins: A highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar]
- Eleftherianos, I.; Waterfield, N.R.; Bone, P.; Boundy, S.; Ffrench-Constant, R.H.; Reynolds, S.E. A single locus from the entomopathogenic bacterium Photorhabdus luminescens inhibits activated Manduca sexta phenoloxidase. FEMS Microbiol. Lett. 2009, 293, 170–176. [Google Scholar]
- Schaller, A.; Kuhn, R.; Kuhnert, P.; Nicolet, J.; Anderson, T.J.; MacInnes, J.I.; Segers, R.P.; Frey, J. Characterization of Apxiva, a new RTX determinant of Actinobacillus pleuropneumoniae. Microbiology 1999, 145, 2105–2116. [Google Scholar]
- Aloy, P.; Russell, R.B. Interprets: Protein interaction prediction through tertiary structure. Bioinformatics 2003, 19, 161–162. [Google Scholar] [CrossRef]
- Ebeling, W.; Hennrich, N.; Klockow, M.; Metz, H.; Orth, H.D.; Lang, H. Proteinase K from Tritirachium album Limber. Eur. J. Biochem. 1974, 47, 91–97. [Google Scholar]
- Kraus, E.; Kiltz, H.H.; Femfert, U.F. The specificity of proteinase K against oxidized insulin B chain. Hoppe Seylers Z. Physiol. Chem. 1976, 357, 233–237. [Google Scholar]
- Lindler, L.E.; Plano, G.V.; Burland, V.; Mayhew, G.F.; Blattner, F.R. Complete DNA sequence and detailed analysis of the Yersinia pestis KIM5 plasmid encoding murine toxin and capsular antigen. Infect. Immun. 1998, 66, 5731–5742. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- European Bioinformatics Institute. Available online: http://www.ebi.ac.uk/Tools/clustalw2 (accessed on 14 March 2014).
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ullah, I.; Jang, E.-K.; Kim, M.-S.; Shin, J.-H.; Park, G.-S.; Khan, A.R.; Hong, S.-J.; Jung, B.-K.; Choi, J.; Park, Y.; et al. Identification and Characterization of the Insecticidal Toxin “Makes Caterpillars Floppy” in Photorhabdus temperata M1021 Using a Cosmid Library. Toxins 2014, 6, 2024-2040. https://doi.org/10.3390/toxins6072024
Ullah I, Jang E-K, Kim M-S, Shin J-H, Park G-S, Khan AR, Hong S-J, Jung B-K, Choi J, Park Y, et al. Identification and Characterization of the Insecticidal Toxin “Makes Caterpillars Floppy” in Photorhabdus temperata M1021 Using a Cosmid Library. Toxins. 2014; 6(7):2024-2040. https://doi.org/10.3390/toxins6072024
Chicago/Turabian StyleUllah, Ihsan, Eun-Kyung Jang, Min-Sung Kim, Jin-Ho Shin, Gun-Seok Park, Abdur Rahim Khan, Sung-Jun Hong, Byung-Kwon Jung, JungBae Choi, YeongJun Park, and et al. 2014. "Identification and Characterization of the Insecticidal Toxin “Makes Caterpillars Floppy” in Photorhabdus temperata M1021 Using a Cosmid Library" Toxins 6, no. 7: 2024-2040. https://doi.org/10.3390/toxins6072024