Protective Antigen-Specific Memory B Cells Persist Years after Anthrax Vaccination and Correlate with Humoral Immunity

Abstract
:1. Introduction
2. Results and Discussion
2.1. Functional PA-Specific Memory B Cells Are Retained for Years Following Vaccination

{kind=link}
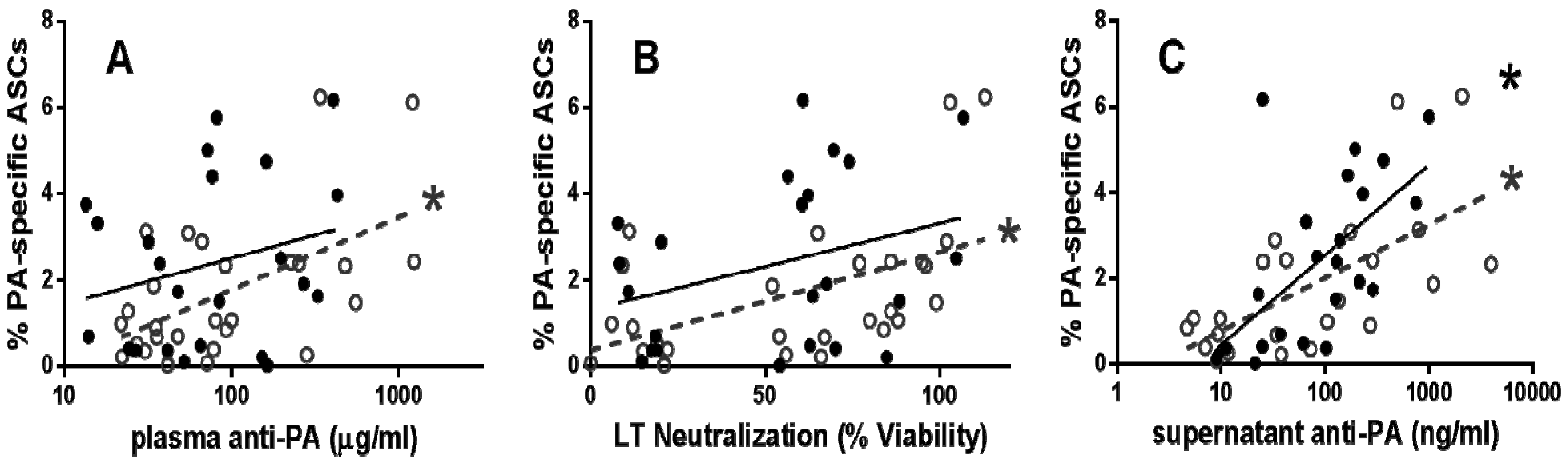{kind=link}
| Early post-vaccination individuals (n = 27) | Late post-vaccination individuals (n = 23) | |
|---|---|---|
| Age at collection | ||
| Average (SD) | 26.2 (5.2) | 34.4 (5.6) |
| Median (range) | 25 (19–41) | 35 (23–45) |
| Years since last vaccination | ||
| Average (SD) | 0.39 (0.09) | 3.09 (0.68) |
| Median (range) | 0.42 (0.25–0.5) | 3.05 (2.10–4.33) |
| Number of vaccinations | ||
| Average (SD) | 4.4 (1.2) | 6 (1.4) |
| Median (range) | 4 (3–7) | 6 (4–9) |

2.2. Functional PA-Specific Memory B Cells Are Retained for Years Following Vaccination

3. Experimental Section
3.1. Collection of Human Blood Samples
3.2. Anti-PA IgG Concentration and LTNA
3.3. Memory B Cell ELISPOT
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pauli, N.T.; Henry Dunand, C.J.; Wilson, P.C. Exploiting human memory B cell heterogeneity for improved vaccine efficacy. Front. Immunol. 2011, 2, 77. [Google Scholar]
- Crotty, S.; Ahmed, R. Immunological memory in humans. Semin. Immunol. 2004, 16, 197–203. [Google Scholar]
- Crotty, S.; Aubert, R.D.; Glidewell, J.; Ahmed, R. Tracking human antigen-specific memory B cells: a sensitive and generalized ELISPOT system. J. Immunol. Methods 2004, 286, 111–122. [Google Scholar] [CrossRef]
- Hammarlund, E.; Lewis, M.W.; Hansen, S.G.; Strelow, L.I.; Nelson, J.A.; Sexton, G.J.; Hanifin, J.M.; Slifka, M.K. Duration of antiviral immunity after smallpox vaccination. Nat. Med. 2003, 9, 1131–1137. [Google Scholar] [CrossRef]
- Crotty, S.; Felgner, P.; Davies, H.; Glidewell, J.; Villarreal, L.; Ahmed, R. Cutting edge: Long-term B cell memory in humans after smallpox vaccination. J. Immunol. 2003, 171, 4969–4973. [Google Scholar] [CrossRef]
- Leyendeckers, H.; Odendahl, M.; Lohndorf, A.; Irsch, J.; Spangfort, M.; Miltenyi, S.; Hunzelmann, N.; Assenmacher, M.; Radbruch, A.; Schmitz, J.; et al. Correlation analysis between frequencies of circulating antigen-specific IgG-bearing memory B cells and serum titers of antigen-specific IgG. Eur. J. Immunol. 1999, 29, 1406–1417. [Google Scholar]
- Ndungu, F.M.; Olotu, A.; Mwacharo, J.; Nyonda, M.; Apfeld, J.; Mramba, L.K.; Fegan, G.W.; Bejon, P.; Marsh, K. Memory B cells are a more reliable archive for historical antimalarial responses than plasma antibodies in no-longer exposed children. Proc. Natl. Acad. Sci. USA 2012, 109, 8247–8252. [Google Scholar] [CrossRef]
- Plotkin, S.A. Vaccines: Correlates of vaccine-induced immunity. Clin. Infect. Dis. 2008, 47, 401–409. [Google Scholar] [CrossRef]
- Plotkin, S.A. Correlates of protection induced by vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef]
- Reuveny, S.; White, M.D.; Adar, Y.Y.; Kafri, Y.; Altboum, Z.; Gozes, Y.; Kobiler, D.; Shafferman, A.; Velan, B. Search for correlates of protective immunity conferred by anthrax vaccine. Infect. Immun. 2001, 69, 2888–2893. [Google Scholar] [CrossRef]
- Quinn, C.P.; Sabourin, C.L.; Niemuth, N.A.; Li, H.; Semenova, V.A.; Rudge, T.L.; Mayfield, H.J.; Schiffer, J.; Mittler, R.S.; Ibegbu, C.C.; et al. A three-dose intramuscular injection schedule of anthrax vaccine adsorbed generates sustained humoral and cellular immune responses to protective antigen and provides long-term protection against inhalation anthrax in rhesus macaques. Clin. Vaccine Immunol. 2012, 19, 1730–1745. [Google Scholar] [CrossRef]
- Crowe, S.R.; Ash, L.L.; Engler, R.J.; Ballard, J.D.; Harley, J.B.; Farris, A.D.; James, J.A. Select human anthrax protective antigen epitope-specific antibodies provide protection from lethal toxin challenge. J. Infect. Dis. 2010, 202, 251–260. [Google Scholar] [CrossRef]
- Crowe, S.R.; Garman, L.; Engler, R.J.; Farris, A.D.; Ballard, J.D.; Harley, J.B.; James, J.A. Anthrax vaccination induced anti-lethal factor IgG: Fine specificity and neutralizing capacity. Vaccine 2011, 29, 3670–3678. [Google Scholar]
- Tross, D.; Klinman, D.M. Effect of CpG oligonucleotides on vaccine-induced B cell memory. J. Immunol. 2008, 181, 5785–5790. [Google Scholar]
- Quinn, C.P.; Dull, P.M.; Semenova, V.; Li, H.; Crotty, S.; Taylor, T.H.; Steward-Clark, E.; Stamey, K.L.; Schmidt, D.S.; Stinson, K.W.; et al. Immune responses to Bacillus anthracis protective antigen in patients with bioterrorism-related cutaneous or inhalation anthrax. J. Infect. Dis. 2004, 190, 1228–1236. [Google Scholar] [CrossRef]
- Goossens, P.L. Animal models of human anthrax: The Quest for the Holy Grail. Mol. Aspects Med. 2009, 30, 467–480. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Anthrax in Humans and Animals, 4th ed.; WHO: Geneva, Switzerland, 2004. [Google Scholar]
- Allen, J.S.; Skowera, A.; Rubin, G.J.; Wessely, S.; Peakman, M. Long-lasting T cell responses to biological warfare vaccines in human vaccinees. Clin. Infect. Dis. 2006, 43, 1–7. [Google Scholar]
- Nguyen, M.L.; Terzyan, S.; Ballard, J.D.; James, J.A.; Farris, A.D. The major neutralizing antibody responses to recombinant anthrax lethal factor and edema factors are directed to non-cross-reactive epitopes. Infect. Immun. 2009, 77, 4713–4723. [Google Scholar]
- Mohamed, N.; Li, J.; Ferreira, C.S.; Little, S.F.; Friedlander, A.M.; Spitalny, G.L.; Casey, L.S. Enhancement of anthrax lethal toxin cytotoxicity: A subset of monoclonal antibodies against protective antigen increases lethal toxin-mediated killing of murine macrophages. Infect. Immun. 2004, 72, 3276–3283. [Google Scholar]
- Quinn, C.P.; Semenova, V.A.; Elie, C.M.; Romero-Steiner, S.; Greene, C.; Li, H.; Stamey, K.; Steward-Clark, E.; Schmidt, D.S.; Mothershed, E.; et al. Specific, sensitive, and quantitative enzyme-linked immunosorbent assay for human immunoglobulin G antibodies to anthrax toxin protective antigen. Emerg. Infect. Dis. 2002, 8, 1103–1110. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Garman, L.; Smith, K.; Farris, A.D.; Nelson, M.R.; Engler, R.J.M.; James, J.A. Protective Antigen-Specific Memory B Cells Persist Years after Anthrax Vaccination and Correlate with Humoral Immunity. Toxins 2014, 6, 2424-2431. https://doi.org/10.3390/toxins6082424
Garman L, Smith K, Farris AD, Nelson MR, Engler RJM, James JA. Protective Antigen-Specific Memory B Cells Persist Years after Anthrax Vaccination and Correlate with Humoral Immunity. Toxins. 2014; 6(8):2424-2431. https://doi.org/10.3390/toxins6082424
Chicago/Turabian StyleGarman, Lori, Kenneth Smith, A. Darise Farris, Michael R. Nelson, Renata J. M. Engler, and Judith A. James. 2014. "Protective Antigen-Specific Memory B Cells Persist Years after Anthrax Vaccination and Correlate with Humoral Immunity" Toxins 6, no. 8: 2424-2431. https://doi.org/10.3390/toxins6082424