Application of Hydrogen Peroxide to the Control of Eutrophic Lake Systems in Laboratory Assays

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Physicochemical and Biological Conditions of the Shallow Lake

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Time Zero | Light | 48 h after H2O2 addition | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 0.17 (mg L−1) | 0.33 (mg L−1) | 0.83 (mg L−1) | 1.67 (mg L−1) | 3.33 (mg L−1) | |||
| Conductivity (μS cm−1) | 599 ± 15 | Yes | 618 ± 12 | 666 ± 12 | 662 ± 20 | 665 ± 27 | 630 ± 19 | 640 ± 18 |
| No | 680 ± 18 | 667 ± 19 | 670 ± 21 | 660 ± 25 | 630 ± 20 | 645 ± 20 | ||
| pH | 9.47 ± 0.20 | Yes | 8.88 ± 0.10 | 9.05 ± 0.13 | 9.00 ± 0.12 | 9.06 ± 0.09 | 9.19 ± 0.12 | 9.22 ± 0.12 |
| No | 7.87 ± 0.12 | 7.45 ± 0.18 | 7.44 ± 0.10 | 7.68 ± 0.15 | 8.06 ± 0.15 | 8.20 ± 0.10 | ||
| Optical Density | 0.37 ± 0.02 | Yes | 0.08 ± 0.01 | 0.07 ± 0.01 | 0.09 ± 0.02 | 0.09 ± 0.01 | 0.08 ± 0.02 | 0.08 ± 0.02 |
| No | 0.08 ± 0.02 | 0.07 ± 0.01 | 0.09 ± 0.02 | 0.09 ± 0.01 | 0.08 ± 0.01 | 0.07 ± 0.01 | ||
| Total Phosphorus (mg L−1) | 0.89 ± 0.10 | Yes | 0.87 ± 0.10 | 0.83 ± 0.12 | 0.82 ± 0.09 | 0.81 ± 0.12 | 0.87 ± 0.03 | 0.90 ± 0.08 |
| No | 0.89 ± 0.09 | 0.79 ± 0.10 | 0.81 ± 0.12 | 0.96 ± 0.15 | 0.91 ± 0.06 | 0.94 ± 0.09 | ||
| Dissolved total Phosphorus (mg L−1) | 0.20 ± 0.08 | Yes | 0.18 ± 0.08 | 0.40 ± 0.10 | 0.40 ± 0.09 | 0.55 ± 0.13 | 0.53 ± 0.09 | 0.55 ± 0.09 |
| No | 0.16 ± 0.08 | 0.30 ± 0.12 | 0.31 ± 0.13 | 0.47 ± 0.12 | 0.49 ± 0.10 | 0.53 ± 010 | ||
| Chl-a (μg L−1) | 530 ± 10 | Yes | 445 ± 12 | 117 ± 5 | 74 ± 3 | 77 ± 8 | 71 ± 9 | 58 ± 8 |
| No | 420 ± 22 | 320 ± 9 | 75 ± 6 | 81 ± 8 | 117 ± 7 | 111 ± 5 | ||
| COD (mgO2 L−1) | 243 ± 12 | Yes | 212 ± 8 | 142 ± 6 | 129 ± 9 | 130 ± 9 | 120 ± 10 | 126 ± 5 |
| No | 213 ± 10 | 226 ± 15 | 205 ± 11 | 200 ± 12 | 180 ± 9 | 119 ± 7 | ||
| Total coliforms (MPN × 100 mL−1) | 4600 | Yes | 4600 | 2800 | 2400 | 150 | 90 | <3 |
| No | 4600 | 2800 | 2400 | 280 | 93 | <3 | ||
| Fecal coliforms (MPN × 100 mL−1) | 4600 | Yes | 4600 | 2100 | 2100 | 130 | 11 | <3 |
| No | 4600 | 2100 | 2100 | 210 | 14 | <3 | ||
2.2. Effect of H2O2 on Total and Faecal Coliform Bacteria Counts
2.3. Effect of H2O2 on Chemical Parameters

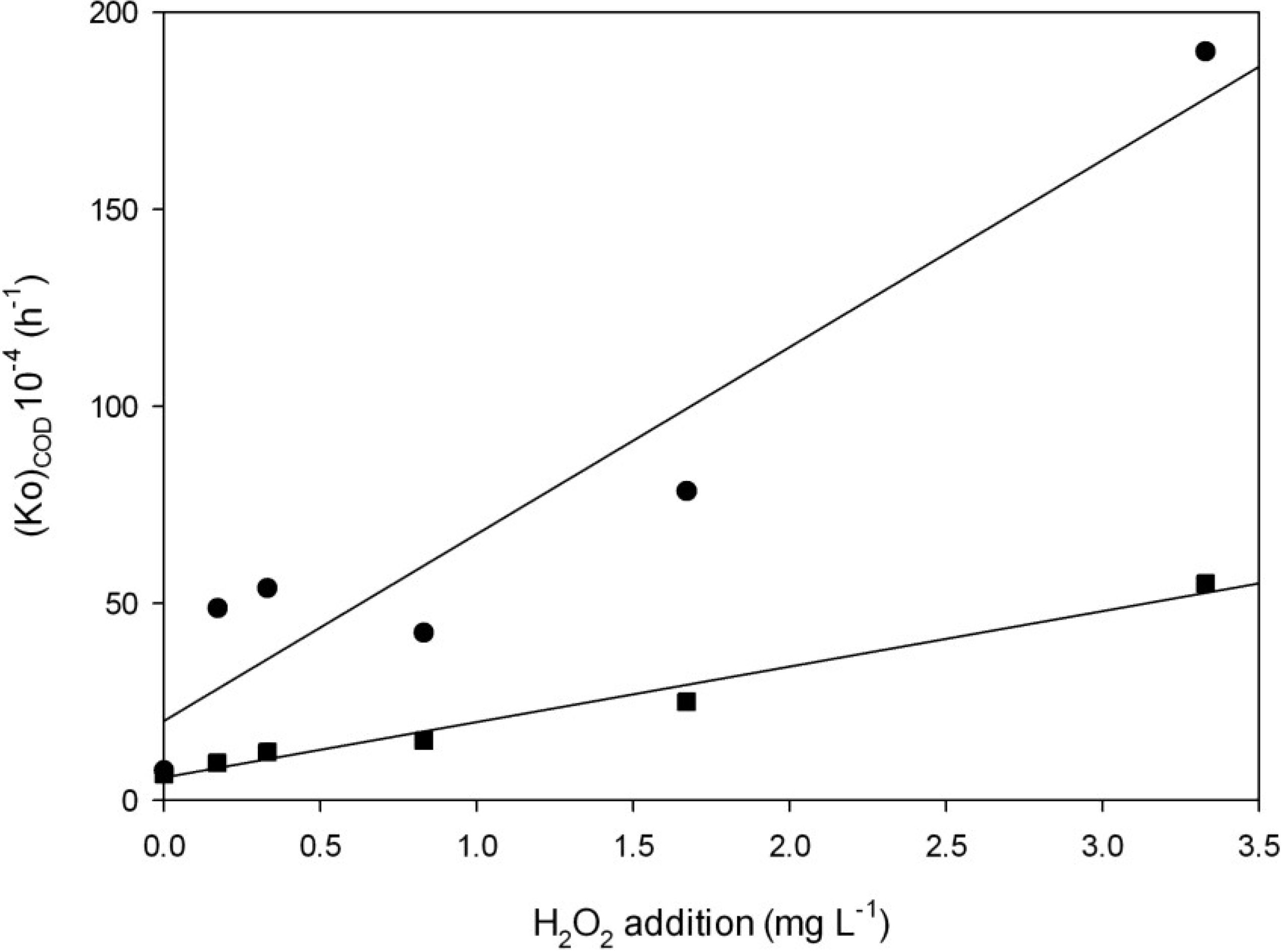
2.3.1. Effect of H2O2 and Light on Chemical Oxygen Demand (COD)
| Chemical oxygen demand (COD) Decay | ||||||
| H2O2 addition | With light | Without light | ||||
| (Ko)COD (h)−1 | R2 | (tm)COD (h) | (Ko)COD (h)−1 | R2 | (tm)COD (h) | |
| 0 | 7.5 × 10−4 ± 5.1 × 10−4 | 0.55 | 925 ± 629 a | 6.5 × 10−4 ± 1.2 × 10−4 | 0.86 | 1050 ± 194 a |
| 0.17 mg·L−1 | 4.9 × 10−3 ± 1.5 × 10−3 | 0.70 | 142 ± 43 b | 9.5 × 10−4 ± 3.8 × 10−4 | 0.55 | 728 ± 291 a |
| 0.33 mg·L−1 | 5.4 × 10−3 ± 1.0 × 10−3 | 0.56 | 128 ± 24 b | 1.3 × 10−3 ± 6.3 × 10−4 | 0.45 | 533 ± 258 a |
| 0.83 mg·L−1 | 4.3 × 10−3 ± 9.5 × 10−4 | 0.87 | 163 ± 36 b | 1.5 × 10−3 ± 1.5 × 10−4 | 0.44 | 459 ± 46 b |
| 1.67 mg·L−1 | 7.8 × 10−3 ± 4.9 × 10−3 | 0.70 | 88 ± 55 b | 2.6 × 10−3 ± 7.5 × 10−4 | 0.34 | 277 ± 80 b |
| 3.33 mg·L−1 | 1.9 × 10−2 ± 9.9 × 10−3 | 0.85 | 36 ± 18 b | 5.6 × 10−3 ± 2.5 × 10−3 | 0.50 | 126 ± 56c c |
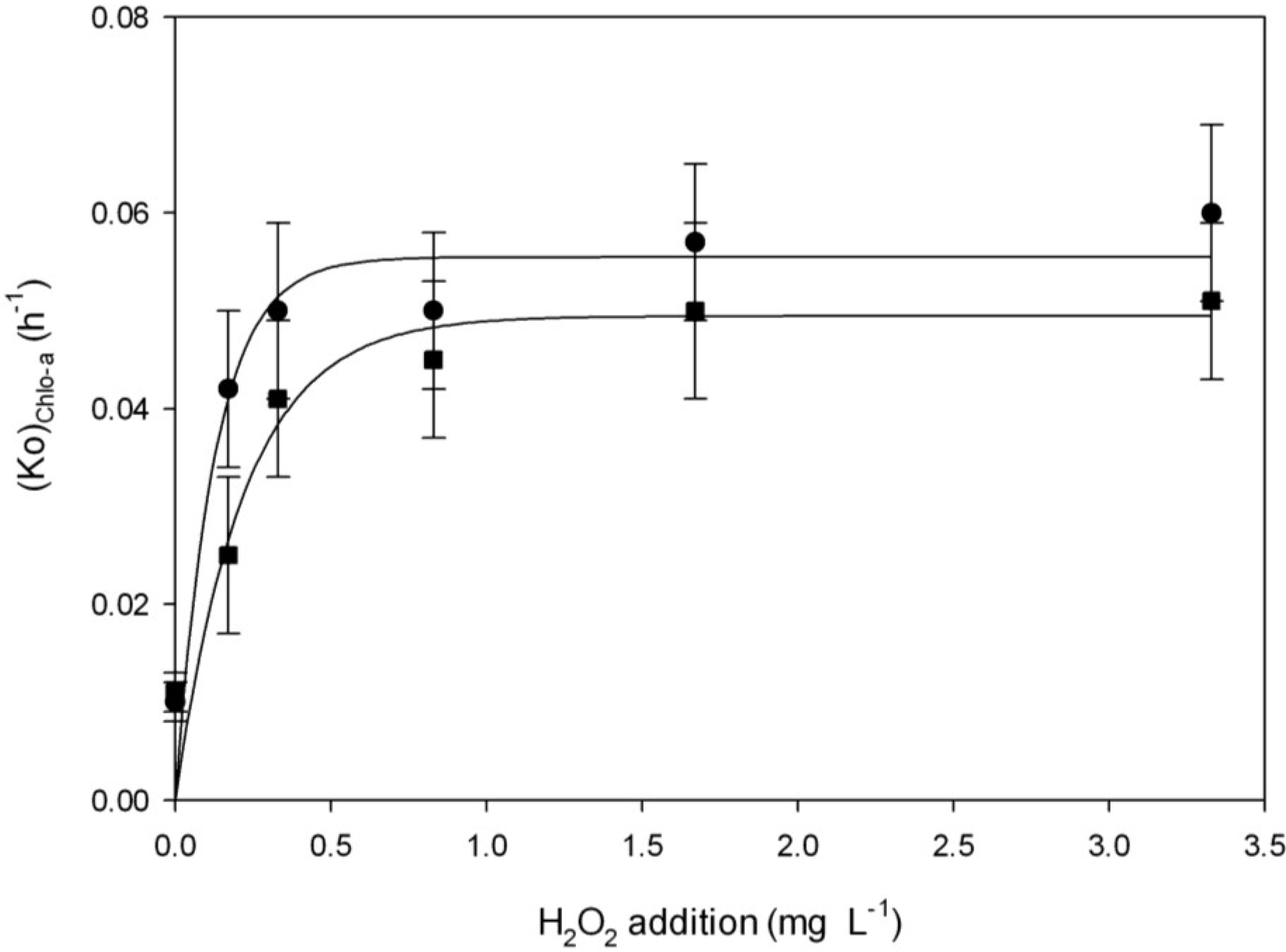
| Chlorophyll-a (Chl-a) Decay | ||||||
| H2O2 addition | With light | Without light | ||||
| (Ko)Chl-a (h)−1 | R2 | (tm)Chl-a (h) | (Ko)Chl-a (h)−1 | R2 | (tm)Chl-a (h) | |
| 0 | 0.010 ± 0.002 | 0.84 | 69 ± 13.8 a | 0.011 ± 0.002 | 0.86 | 63 ± 11.4 a |
| 1.67 mg·L−1 | 0.057 ± 0.008 | 0.89 | 12 ± 1.7 b | 0.050 ± 0.009 | 0.79 | 14 ± 2.5 b |
| 0.17 mg·L−1 | 0.042 ± 0.008 | 0.86 | 16 ± 3.1 b | 0.025 ± 0.008 | 0.86 | 28 ± 8.8 b |
| 0.33 mg·L−1 | 0.050 ± 0.009 | 0.83 | 14 ± 2.5 b | 0.041 ± 0.008 | 0.87 | 17 ± 3.3 b |
| 0.83 mg·L−1 | 0.050 ± 0.008 | 0.87 | 14 ± 2.2 b | 0.045 ± 0.008 | 0.85 | 15 ± 2.6 b |
| 3.33 mg·L−1 | 0.060 ± 0.009 | 0.85 | 12 ± 1.8 b | 0.051 ± 0.008 | 0.79 | 14 ± 2.1 b |

2.3.2. Effect of H2O2 and Light on Chl-a Degradation


2.4. The Effect of H2O2 and Light on Phytoplankton Counts

3. Experimental Methodology
3.1. Sample Collection
3.2. Laboratory Incubations and Experimental Design
3.3. Analyzed Parameters and Analytical Methods
Trophic State Index
3.4. Electron Microscopy Studies
3.5. Modelling COD and Chl-a Decay
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Echenique, R.O.; Aguilera, A. Cyanobacteria toxígenas: Aspectos generales para su identificación taxonómica. In Cianobacterias y Cianotoxinas: Identificación, Toxicología, Monitoreo y Evaluación de Riesgo; Giannuzzi, L., Ed.; Moglia Impresiones: Buenos Aires, Argentina, 2009; pp. 37–51. [Google Scholar]
- Environmental Protection Agency. Cyanobacteria and Cyanotoxins: Information for Drinking Water System; Office of Water 4304T, EPA-810F11001; Environmental Protection Agency: Washington, DC, USA, 2012. [Google Scholar]
- Carmichael, W.W. A world overview. One-hundred-twenty-seven years of research on toxic Cyanobacteria. Where do we go from here? In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Kenneth, H., Hudnell, H., Eds.; Springer: New York, NY, USA, 2008; pp. 105–126. [Google Scholar]
- Berry, J. Cyanobacterial Toxins in Food-Webs: Implications for Human and Environmental Health Current Topics in Public Health. Available online: http://dx.doi.org/10.5772/55111 (accessed on 18 January 2014).
- Intergovernmental Panel on Climate Change (IPCC). Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report. In Climate Change 2007; Parry, M.L., Canziani, O.F., Palutikof, J.P., van derLinden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Drábková, M. Methods for Control of the Cyanobacterial Blooms Development in Lakes. Ph.D. Thesis, Masaryk University, Faculty of Science, Research Centre for Environmental Chemistry and Ecotoxicology, Brno, Czech Republic, 2007. [Google Scholar]
- McElhiney, J.; Lawton, L.A. Detection of the cyanobacterial hepatotoxins microcystins. Toxicol. Appl. Pharmacol. 2005, 203, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; World Health Organization, St Edmundsbury Press: Bury St Edmunds, Suffolk, London, UK, 1999. [Google Scholar]
- Barrington, D.J.; Ghadouani, A. Application of hydrogen peroxide for the removal of toxic cyanobacteria and other phytoplankton from wastewater. Environ. Sci. Technol. 2008, 42, 8916–8921. [Google Scholar] [CrossRef] [PubMed]
- Drábková, M.; Admiraal, W.; Marsálek, B. Combined exposure to hydrogen peroxide and light: Selective effects on cyanobacteria, green algae, and diatoms. Environ. Sci. Technol. 2007, 41, 309–314. [Google Scholar]
- Kay, S.H.; Quimby, P.C.; Ouzts, J.D. Photo-enhancement of hydrogen peroxide toxicity to submersed vascular plants and algae. J. Aquat. Plant Manag. 1984, 22, 25–34. [Google Scholar]
- Svrcek, C.; Smith, D.W. Cyanobacteria toxins and the current state of knowledge on water treatment options: A review. J. Environ. Eng. 2004, 3, 155–185. [Google Scholar] [CrossRef]
- Matthijs, H.; Visser, P.; Reeze, B.; Meeuse, J.; Slot, P.; Wijn, G.; Talens, R.; Huisman, J. Selective suppression of harmful cyanobacteria in an entire lake with hydrogen peroxide. Water Res. 2012, 46, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, M.G.; de la Cruz, A.A.; Dionysiou, D.D. Cyanotoxins: New generation of water contaminants. J. Environ. Eng. 2005, 131, 1239–1243. [Google Scholar] [CrossRef]
- Wang, H.; Wang, H. Mitigation of Lake Eutrophication: Loosen nitrogen control and focus on phosphorus abatement. Prog. Nat. Sci. 2009, 19, 1445–1451. [Google Scholar] [CrossRef]
- Ruiz, M.; Granero, M.; Rodríguez, M.I.; Bustamante, M.A.; Ruibal, A.L. Importancia de los sedimentos como fuente interna de fósforo en el Embalse San Roque (Córdoba): Determinación de una metodología para su estudio. In Proceedings of the XX Congreso Nacional del agua y III Simposio de Recursos Hídricos del Cono Sur, Mendoza, Argentina, 9–14 May 2005.
- World Health Organization (WHO) 1999. Available online: http://www.who.int (accessed on 25 January 2014).
- Hou, G.; Song, L.; Liu, J.; Xiao, B.; Lui, Y. Modeling of cyanobacterial blooms in hypereutrophic Lake Dianchi, China. J. Freshw. Ecol. 2004, 19, 623–629. [Google Scholar] [CrossRef]
- Chapman, D. Water Quality Assessment; United Nations Educational, Scientific and Cultural Organization (UNESCO), World Health Organization (WHO) and UNEP Chapman and Hall: London, UK, 1992. [Google Scholar]
- United States Environmental Protection Agency (USEPA). Bacterial Water Quality Standards for Recreational Water (Freshwater and Marine Waters); EPA/823/R-98/003; USEPA: Washington, DC, USA, 2003. [Google Scholar]
- Yoshpe-Purer, M.S.; Eylan, E. Disinfection of water by hydrogen peroxide. Health Lab. Sci. 1968, 5, 233–238. [Google Scholar]
- Liao, C.; Sapers, G.M. Attachment and growth of Salmonella chester on apple fruits and in vivo response of attached bacteria to sanitizer treatments. J. Food Prot. 2000, 63, 876–883. [Google Scholar] [PubMed]
- Forney, C.F.; Rij, R.E.; Dennis-Arrue, R.; Smilanick, J.L. Vapor phase hydrogen peroxide inhibits post-harvest decay of table grapes. Hort. Sci. 1991, 26, 1512–1514. [Google Scholar]
- Davidson, M.P.; Branen, A.L. Antimicrobials in Foods, 2nd ed.; Marcel Dekker, Inc.: New York, NY, USA, 2003. [Google Scholar]
- Lillard, H.S.; Thomson, J.E. Efficacy of hydrogen peroxide as a bactericide in poultry chiller water. J. Food Sci. 1983, 48, 125–126. [Google Scholar] [CrossRef]
- Juven, B.J.; Pierson, M.D. Antibacterial effects of hydrogen peroxide and methods for its detection and quantitation. J. Food Prot. 1996, 59, 1233–1241. [Google Scholar]
- Golterman, H.L. Mineralization of algae under sterile conditions or by bacterial breakdown. Verh. Int. Ver. Limnol. 1964, 15, 544–548. [Google Scholar]
- Fallon, R.D.; Brock, T.D. Decomposition of blue-green algal (Cyanobacterial) blooms in lake Mendota, Wisconsin. Appl. Environ. Microbiol. 1979, 37, 5820–5830. [Google Scholar]
- Agustina, T.E.; Ang, H.M.; Vareek, V.K. A review of synergistic effect of photocatalysis and ozonation on wastewater treatment. J. Photochem. Photobiol. C 2005, 6, 264–273. [Google Scholar] [CrossRef]
- Cooper, W.J.; Zika, R.G. Photochemical formation of hydrogen peroxide in surface and ground waters exposed to sunlight. Science 1983, 220, 711–712. [Google Scholar] [CrossRef] [PubMed]
- Pisarevsky, A.M.; Polozova, I.P.; Hockridge, P.M. Chemical oxygen demand. Russ. J. Appl. Chem. 2005, 78, 101–107. [Google Scholar] [CrossRef]
- Zhang, Y.L.; van Dijk, M.A.; Liu, M.L.; Zhu, G.W.; Qin, B.Q. The contribution of phytoplankton degradation to chromophoric dissolved organic matter (CDOM) in eutrophic shallow lakes: Field and experimental evidence. Water Res. 2009, 43, 4685–4697. [Google Scholar] [CrossRef] [PubMed]
- Barroin, G.; Feuillade, G. Hydrogen peroxide as a potential algaecide for Oscillatoria rubescens D.C. Water Res. 1986, 20, 619–623. [Google Scholar] [CrossRef]
- Randhawa, V.; Thakkar, M.; Wei, L. Applicability of hydrogen peroxide in brown tide control—Culture and microcosm studies. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Qian, H.; Hu, B.; Yu, S.; Pan, X.; Wu, T.; Fu, Z. The effects of hydrogen peroxide on the circadian rhythms of Microcystis aeruginosa. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Skurlatov, Y.I.; Ernestova, L.S. The impact of human activities on freshwater aquatic systems. Acta Hydrochim. Hydrobiol. 1998, 26, 5–12. [Google Scholar] [CrossRef]
- Samuilov, V.; Timofee, K.N.; Sinitsyn, S.V.; Bezryadno, D.V. H2O2 induced inhibition of photosynthetic O2 evolution by Anabaena variabilis cells. Biochem. Mosc. 2004, 69, 926–933. [Google Scholar] [CrossRef]
- Tytler, E.G.; Wong, T.; Codd, G.A. Photoinactivation in vivo of superoxide dismutase and catalase in the cyanobacterium Microcystis aeruginosa. FEMS Microbiol. Lett. 1984, 23, 239–242. [Google Scholar] [CrossRef]
- Tel-Or, E.; Huflejt, M.E.; Packer, L. Hydroperoxide metabolism in cyanobacteria. Arch. Biochem. Biophys. 1986, 246, 396–402. [Google Scholar]
- Mikula, P.; Zezulka, S.; Jancula, D.; Marsalek, B. Metabolic activity and membrane integrity changes in Microcystis aeruginosa—New findings on hydrogen peroxide toxicity in cyanobacteria. Eur. J. Phycol. 2005, 27, 195–206. [Google Scholar]
- World Health Organization (WHO). GEMS/WATER Operational Guide; WHO: Geneva, Switzerland, 1987. [Google Scholar]
- American Public Health Association. Standard Methods for Examination of Water and Wastewater, 21th ed.; American Public Health Association, American Water Works Association, Water Environment Federation: Washington, DC, USA, 2005. [Google Scholar]
- Komarek, J.; Anagnostidis, K. Modern approach to the classification system of cyanophytes. 2. Chroococcales. Arch. Hydrobiol. Suppl. 1986, 73, 157–226. [Google Scholar]
- Utermöhl, H. Vervolkommung der quantitative phytoplankton methodik. Mitt. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar]
- Carlson, R.E. A trophic state index for lakes. Limnol. Oceanogr. 1977, 22, 361–369. [Google Scholar] [CrossRef]
- Aizaki, M.O.; Otsuki, M.; Fukushima, M.; Muraoka, H. Application of Carlson’s trophic state index to Japanese lakes and relationships between the index and other parameters. Verh. Int. Ver. Limnol. 1981, 21, 675–681. [Google Scholar]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bauzá, L.; Aguilera, A.; Echenique, R.; Andrinolo, D.; Giannuzzi, L. Application of Hydrogen Peroxide to the Control of Eutrophic Lake Systems in Laboratory Assays. Toxins 2014, 6, 2657-2675. https://doi.org/10.3390/toxins6092657
Bauzá L, Aguilera A, Echenique R, Andrinolo D, Giannuzzi L. Application of Hydrogen Peroxide to the Control of Eutrophic Lake Systems in Laboratory Assays. Toxins. 2014; 6(9):2657-2675. https://doi.org/10.3390/toxins6092657
Chicago/Turabian StyleBauzá, Letizia, Anabella Aguilera, Ricardo Echenique, Darío Andrinolo, and Leda Giannuzzi. 2014. "Application of Hydrogen Peroxide to the Control of Eutrophic Lake Systems in Laboratory Assays" Toxins 6, no. 9: 2657-2675. https://doi.org/10.3390/toxins6092657