
Zearalenone and Its Derivatives α-Zearalenol and β-Zearalenol Decontamination by Saccharomyces cerevisiae Strains Isolated from Bovine Forage

 ,
,
Abstract
:1. Introduction
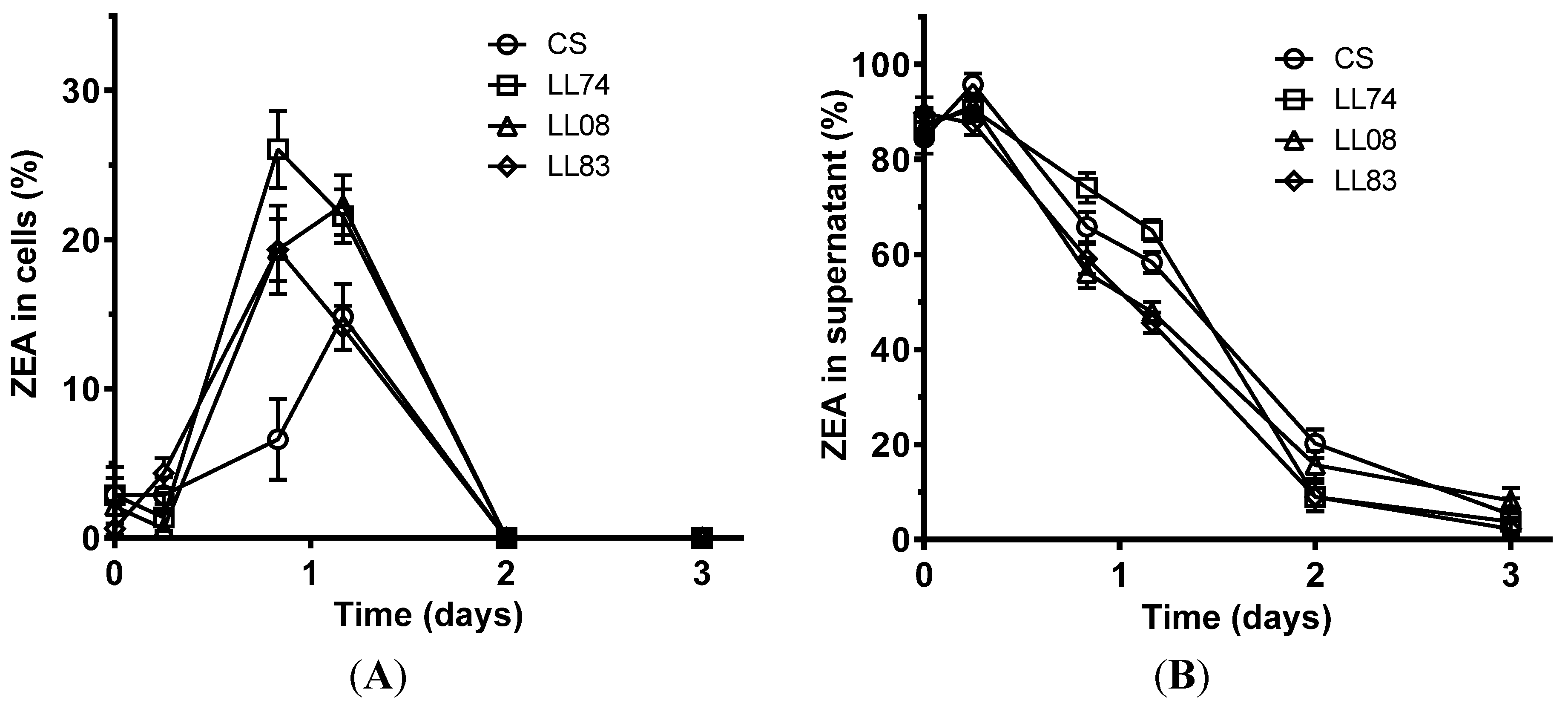
2. Results and Discussion
2.1. Adsorption of ZEA, α-ZOL, and β-ZOL by S. cerevisiae Strains



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | ZEA (µg/mL) | Yeast cells (mg/mL) | pH 3 | pH 6 |
|---|---|---|---|---|
| Adsorption * (%) | Adsorption * (%) | |||
| Controls | 2.1 | 0.0 | --- | --- |
| CS | 2.1 | 2.0 | 31.2 ± 2.3 a | 17.1 ± 1.8 a |
| LL74 | 2.1 | 2.0 | 33.1 ± 1.5 a | 14.4 ± 2.5 a |
| LL08 | 2.1 | 2.0 | 43.1 ± 3.1 b | 23.7 ± 1.6 b |
| LL83 | 2.1 | 2.0 | 34.2 ± 2.6 a | 13.2 ± 1.0 a |
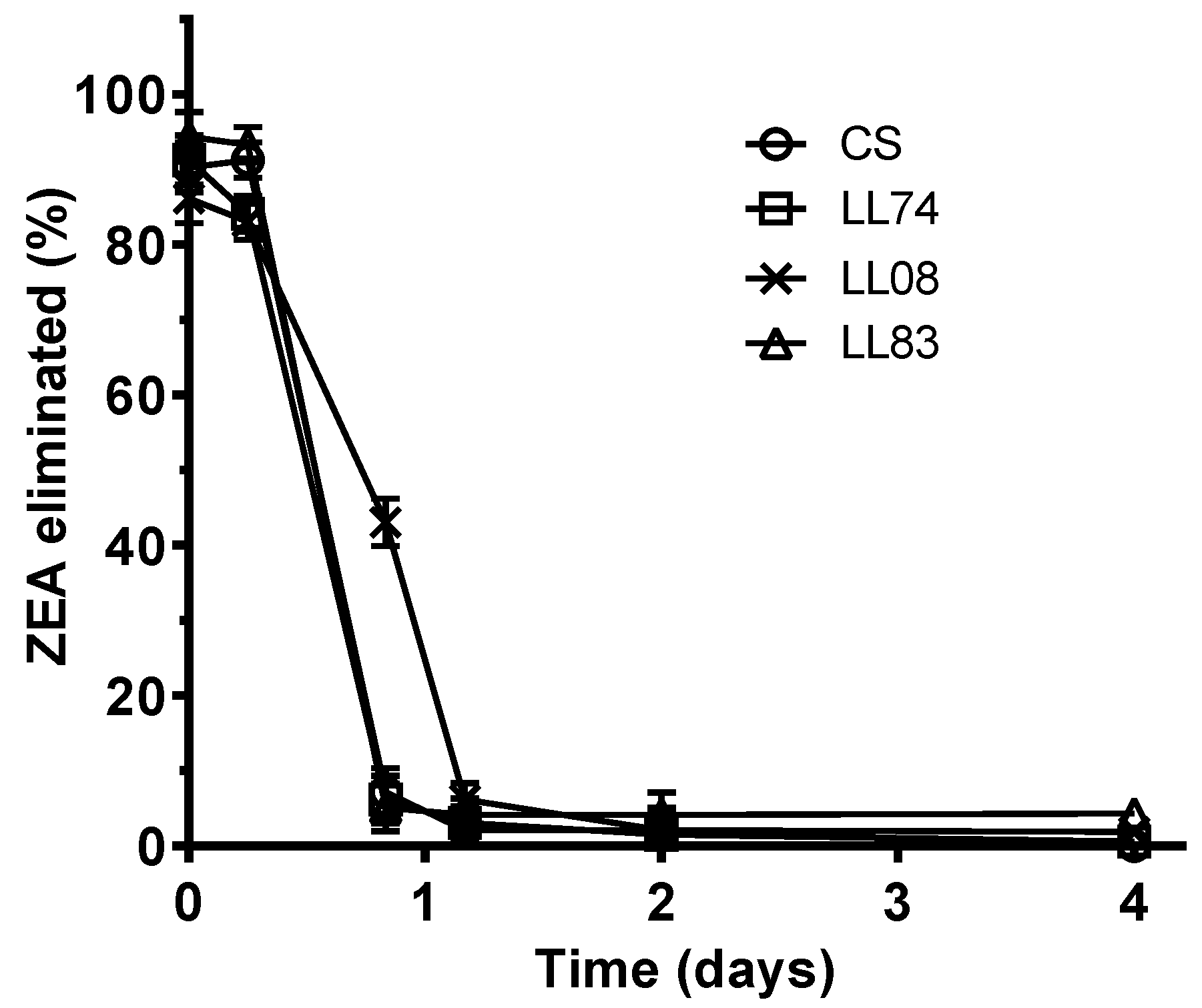
2.2. Biotransformation of ZEA, α-ZOL, and β-ZOL by S. cerevisiae Strains
| Strains | ZEA * (μg/mL) | Logistic model | Curve Fitted Errors | ||
|---|---|---|---|---|---|
| DT50 (h) | DT90 (h) | R2 | Sy.x (%) | ||
| Controls | 1.95 ± 0.08 | --- | --- | --- | --- |
| CS | 0.04 ± 0.01 a | 9 | 18 | 0.9996 | 1.2 |
| LL74 | 0.06 ± 0.02 a | 9 | 18 | 0.9995 | 1.3 |
| LL08 | 0.12 ± 0.05 b | 17 | 28 | 0.9925 | 4.9 |
| LL83 | 0.08 ± 0.02 a,b | 9 | 18 | 0.9961 | 3.8 |


3. Experimental Section
3.1. Biological Material
3.2. Adsorption of ZEA, α-ZOL and β-ZOL by S. cerevisiae Strains
3.3. Biotransformation of ZEA, α-ZOL and β-ZOL by S. cerevisiae Strains
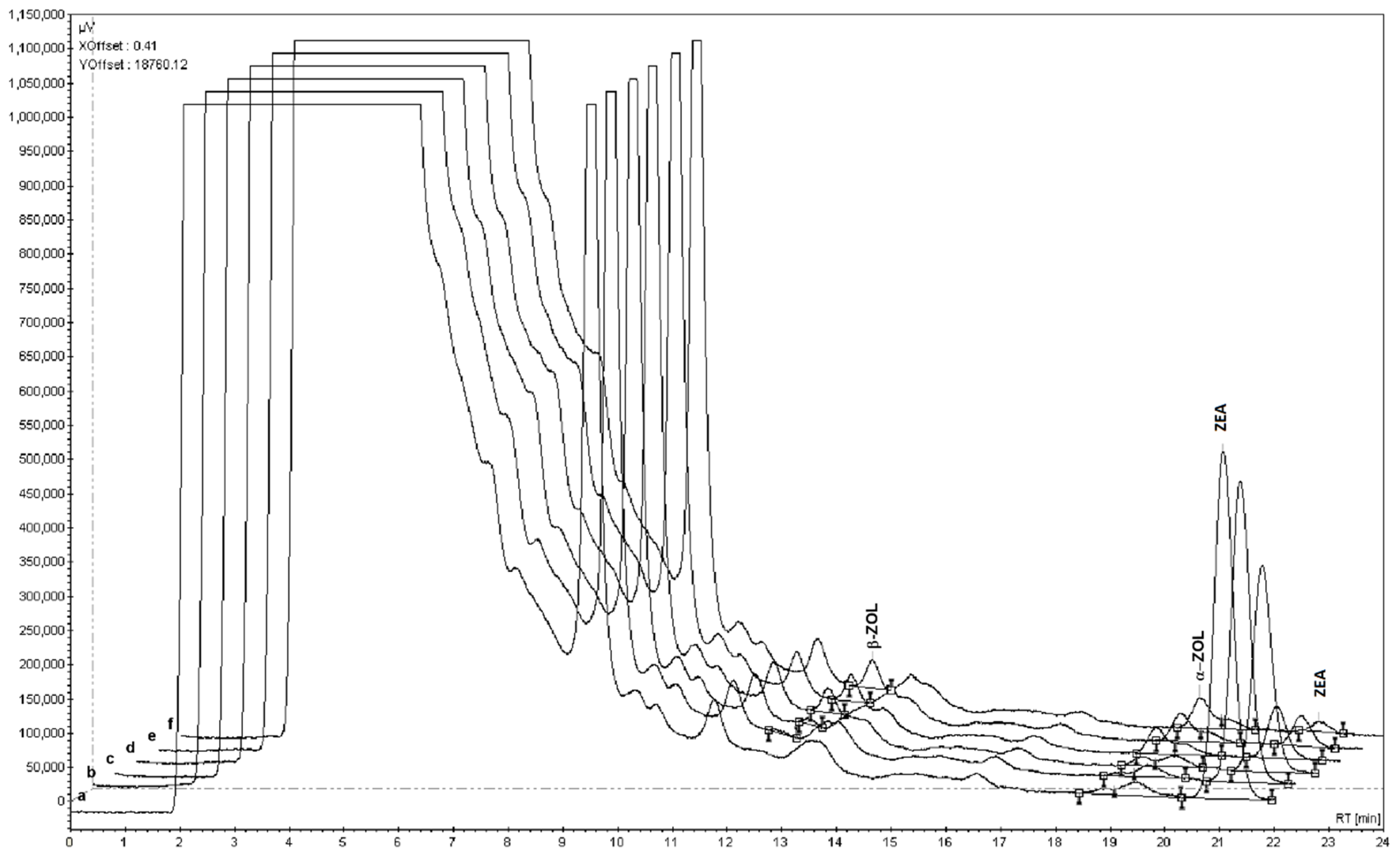
3.4. Extraction and Detection of Mycotoxins
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hussein, H.S.; Brasel, J.M. Toxicity, metabolism, and impact of mycotoxins on humans and animals. Toxicol. 2001, 167, 101–134. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Malekinejad, H.; Maas-Bakker, R.F.; Fink-Gremmels, J. Bioactivation of zearalenone by porcine hepatic biotransformation. Vet. Res. 2005, 36, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Shier, W.T.; Shier, A.C.; Xie, W.; Mirocha, C.J. Structure-activity relationships for human estrogenic activity in zearalenone mycotoxins. Toxicon 2001, 39, 1435–1438. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on the risks for public health related to the presence of zearalenone in food. EFSA J. 2011, 9, 124. [Google Scholar]
- Çelýk, K.; Denlý, M.; Savas, T. Reduction of toxic effects of aflatoxin B1 by using baker yeast (Saccharomyces cerevisiae) in growing broiler chicks diets. Rev. Bras. Zootec. 2003, 32, 615–619. [Google Scholar] [CrossRef]
- Santin, E.; Paulillo, A.C.; Krabbe, E.L.; Alessi, A.C.; Polveiro, W.J.C.; Maiorka, A. Low level of aflatoxin in broiler at experimental conditions. Use of cell wall yeast as adsorbent of aflatoxin. Arch. Vet. Sci. 2003, 8, 51–55. [Google Scholar] [CrossRef]
- Joannis-Cassan, C.; Tozlovanu, M.; Hadjeba-Medjdoub, K.; Ballet, N.; Pfohl-Leszkowicz, A. Binding of zearalenone, aflatoxin B1, and ochratoxin A by yeast-based products: a method for quantification of adsorption performance. J. Food Prot. 2011, 74, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Chaucheyras-Durand, F.; Durand, H. Probiotics in animal nutrition and health. Benef. Microbes. 2010, 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Shetty, P.H.; Jespersen, L. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci. Tech. 2006, 17, 48–55. [Google Scholar] [CrossRef]
- Yiannikouris, A.; Poughon, L.; Cameleyre, X.; Dussap, C.G.; Francois, J.; Bertin, G.; Jouany, J.P. A novel technique to evaluate interactions between Saccharomyces cerevisiae cell wall and mycotoxins: Application to zearalenone. Biotechnol. Lett. 2003, 25, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Yiannikouris, A.; Andre, G.; Buleon, A.; Jeminet, G.; Canet, I.; Francois, J.; Bertin, G.; Jouany, J.P. Comprehensive conformational study of key interactions involved in zearalenone complexation with beta-D-glucans. Biomacromolecules 2004, 5, 2176–2185. [Google Scholar] [CrossRef] [PubMed]
- Aravind, K.L.; Patil, V.S.; Devegowda, G.; Umakantha, B.; Ganpule, S.P. Efficacy of esterified glucomannan to counteract mycotoxicosis in naturally contaminated feed on performance and serum biochemical and hematological parameters in broilers. Poult. Sci. 2003, 82, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, A.O.; Parlat, S.S.; Yildirim, I. Effect of dietary addition of live yeast (Saccharomyces cerevisiae) on some performance parameters of adult Japanese quail induced by aflatoxicosis. Rev. Med. Vet. 2004, 155, 38–41. [Google Scholar]
- Sahebghalam, H.; Mohamadi Sani, A.; Mehraban, M. Assessing the ability of Saccharomyces cerevisiae to bind Aflatoxin B1 from contaminated medium. Nutr. Food Sci. 2013, 43, 392–397. [Google Scholar] [CrossRef]
- Piotrowska, M.; Masek, A. Saccharomyces Cerevisiae cell wall components as tools for ochratoxin A decontamination. Toxins 2015, 7, 1151–1162. [Google Scholar] [CrossRef] [PubMed]
- Baptista, A.S.; Horii, J.; Calori-Domingues, M.A.; da Glória, E.M.; Salgado, J.M.; Vizioli, M.R. Thermolysed and active yeast to reduce the toxicity of aflatoxin. Sci. Agric. 2002, 59, 257–260. [Google Scholar] [CrossRef]
- Keller, K.M.; De Oliveira, Á.A.; De Almeida, T.X.; Keller, L.A.M.; Queiroz, B.D.; Nunes, L.M.T.; Cavaglieri, L.R.; Rosa, C.A.D.R. Effect of yeast cell wall on the performance of broiler chickens intoxicated with aflatoxin B1. Rev. Bras. Med. Vet. 2012, 34, 101–105. [Google Scholar]
- Armando, M.R.; Pizzolitto, R.P.; Dogi, C.A.; Cristofolini, A.; Merkis, C.; Poloni, V.; Dalcero, A.M.; Cavaglieri, L.R. Adsorption of ochratoxin A and zearalenone by potential probiotic Saccharomyces cerevisiae strains and its relation with cell wall thickness. J. Appl. Microbiol. 2012, 113, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Böswald, C.; Engelhardt, G.; Vogel, H.; Wallnöfer, P.R. Metabolism of the Fusarium mycotoxins zearalenone and deoxynivalenol by yeast strains of technological relevance. Nat. Toxins 1995, 3, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.M.; Kanhere, S.R.; Daley, E.F.; Farber, J.M. Fermentation of wort containing deoxynivalenol and zearalenone. Mycotoxin Res. 1992, 8, 58–66. [Google Scholar] [CrossRef] [PubMed]
- González Pereyra, M.L.; Sulyok, M.; Baralla, V.; Dalcero, A.M.; Krska, R.; Chulze, S.; Cavaglieri, L.R. Evaluation of zearalenone, α-zearalenol, β-zearalenol, zearalenone 4-sulfate and β-zearalenol 4-glucoside levels during the ensiling process. World Mycotoxin J. 2014, 7, 291–295. [Google Scholar] [CrossRef]
- FOCUS. Guidance Document on Estimating Persistence and Degradation Kinetics from Environmental Fate Studies on Pesticides in EU Registration, Report of the FOCUS work group on degradation kinetics. EC Document Reference Sanco/10058/2005 version 2.0; 2006; pp. 1–434. [Google Scholar]
- Niderkorn, V.; Morgavi, D.P.; Pujos, E.; Tissandier, A.; Boudra, H. Screening of fermentative bacteria for their ability to bind and biotransform deoxynivalenol, zearalenone and fumonisins in an in vitro simulated corn silage model. Food Addit. Contam. 2007, 24, 406–415. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keller, L.; Abrunhosa, L.; Keller, K.; Rosa, C.A.; Cavaglieri, L.; Venâncio, A. Zearalenone and Its Derivatives α-Zearalenol and β-Zearalenol Decontamination by Saccharomyces cerevisiae Strains Isolated from Bovine Forage. Toxins 2015, 7, 3297-3308. https://doi.org/10.3390/toxins7083297
Keller L, Abrunhosa L, Keller K, Rosa CA, Cavaglieri L, Venâncio A. Zearalenone and Its Derivatives α-Zearalenol and β-Zearalenol Decontamination by Saccharomyces cerevisiae Strains Isolated from Bovine Forage. Toxins. 2015; 7(8):3297-3308. https://doi.org/10.3390/toxins7083297
Chicago/Turabian StyleKeller, Luiz, Luís Abrunhosa, Kelly Keller, Carlos Alberto Rosa, Lilia Cavaglieri, and Armando Venâncio. 2015. "Zearalenone and Its Derivatives α-Zearalenol and β-Zearalenol Decontamination by Saccharomyces cerevisiae Strains Isolated from Bovine Forage" Toxins 7, no. 8: 3297-3308. https://doi.org/10.3390/toxins7083297