
Screening and Identification of Novel Ochratoxin A-Producing Fungi from Grapes

Abstract
:
1. Introduction
2. Results
2.1. Preliminary Screening of OTA-Producing Fungi by 365 nm Ultraviolet Light
2.2. Confirmation of Positive Strains by HPLC-FLD
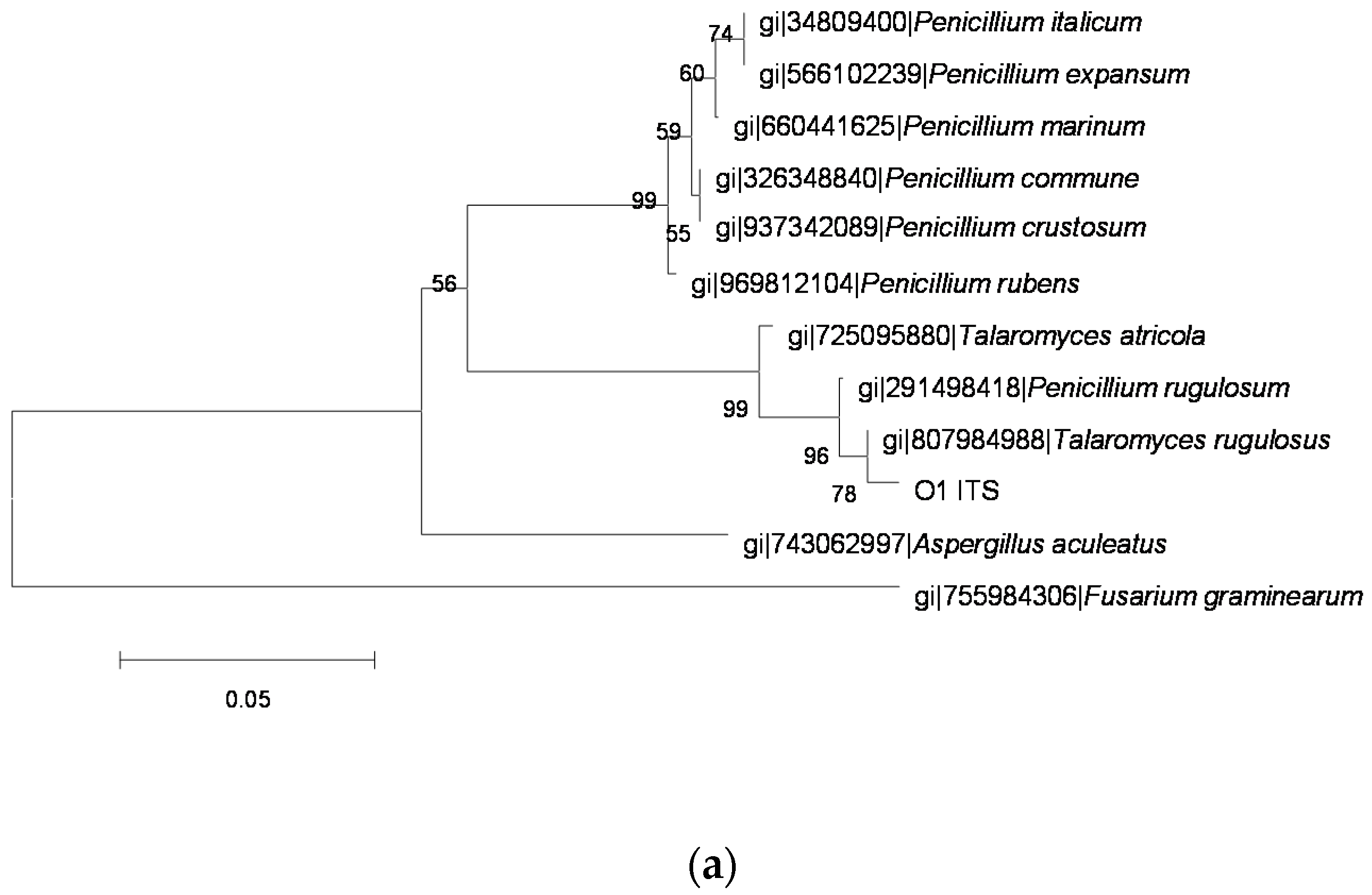
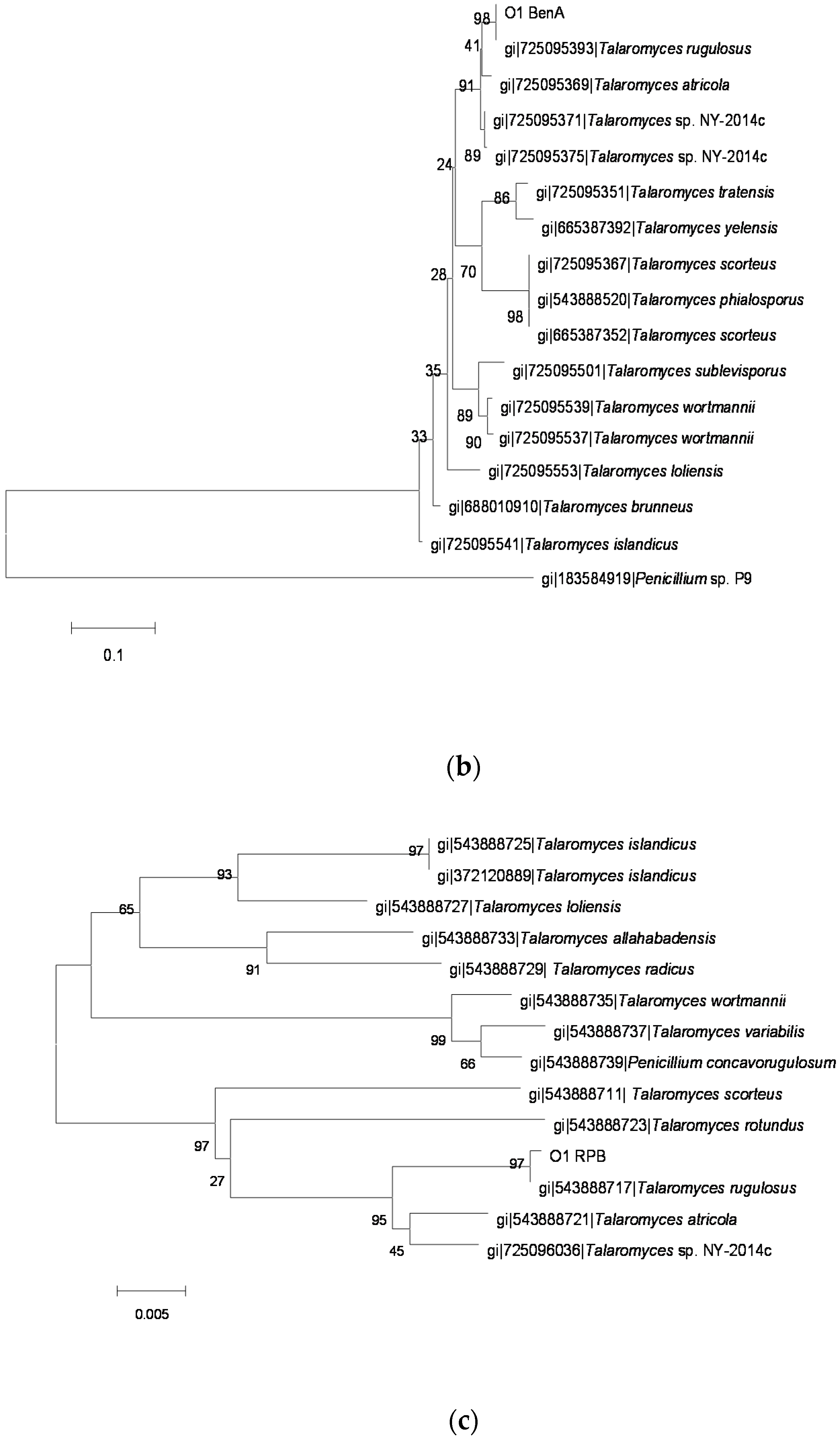
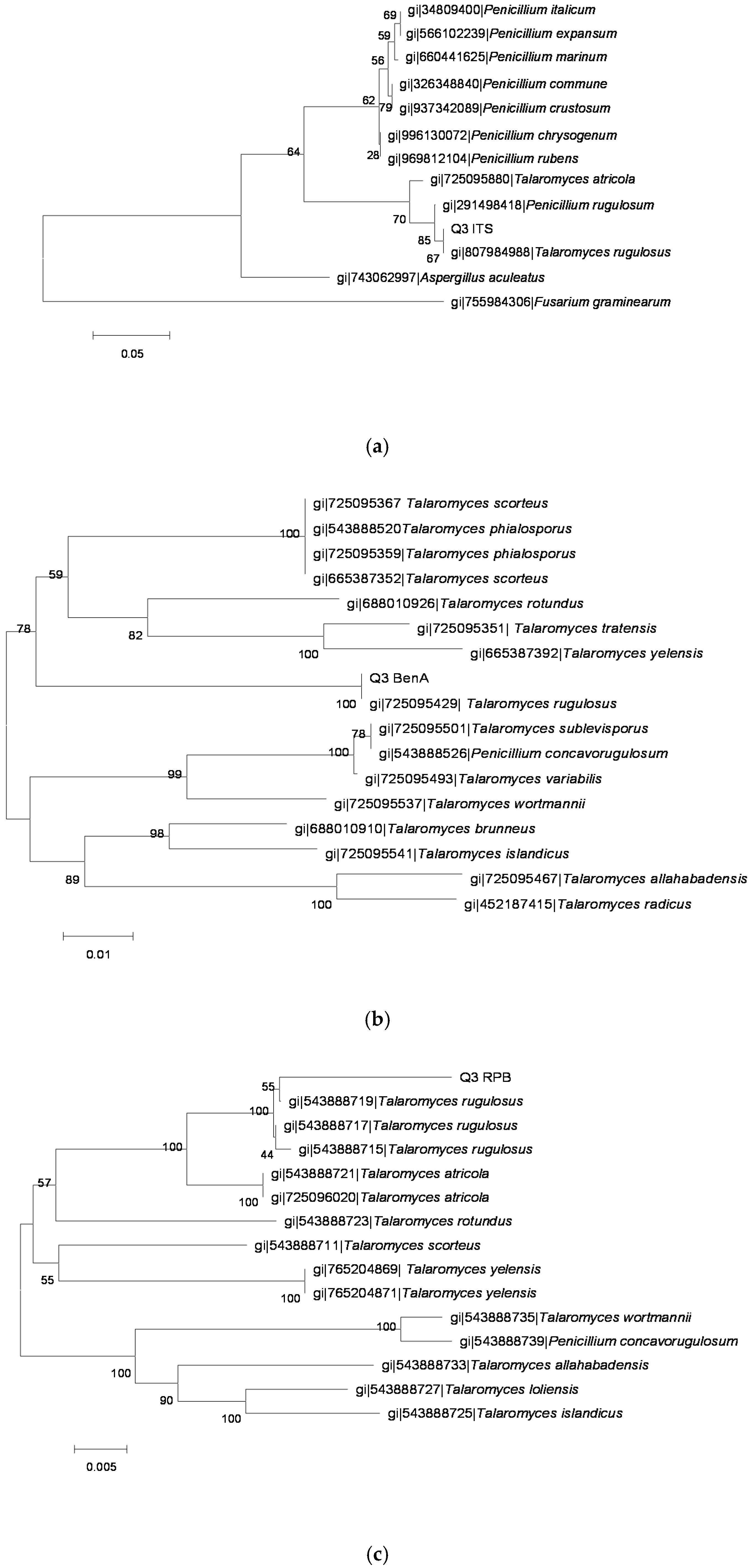
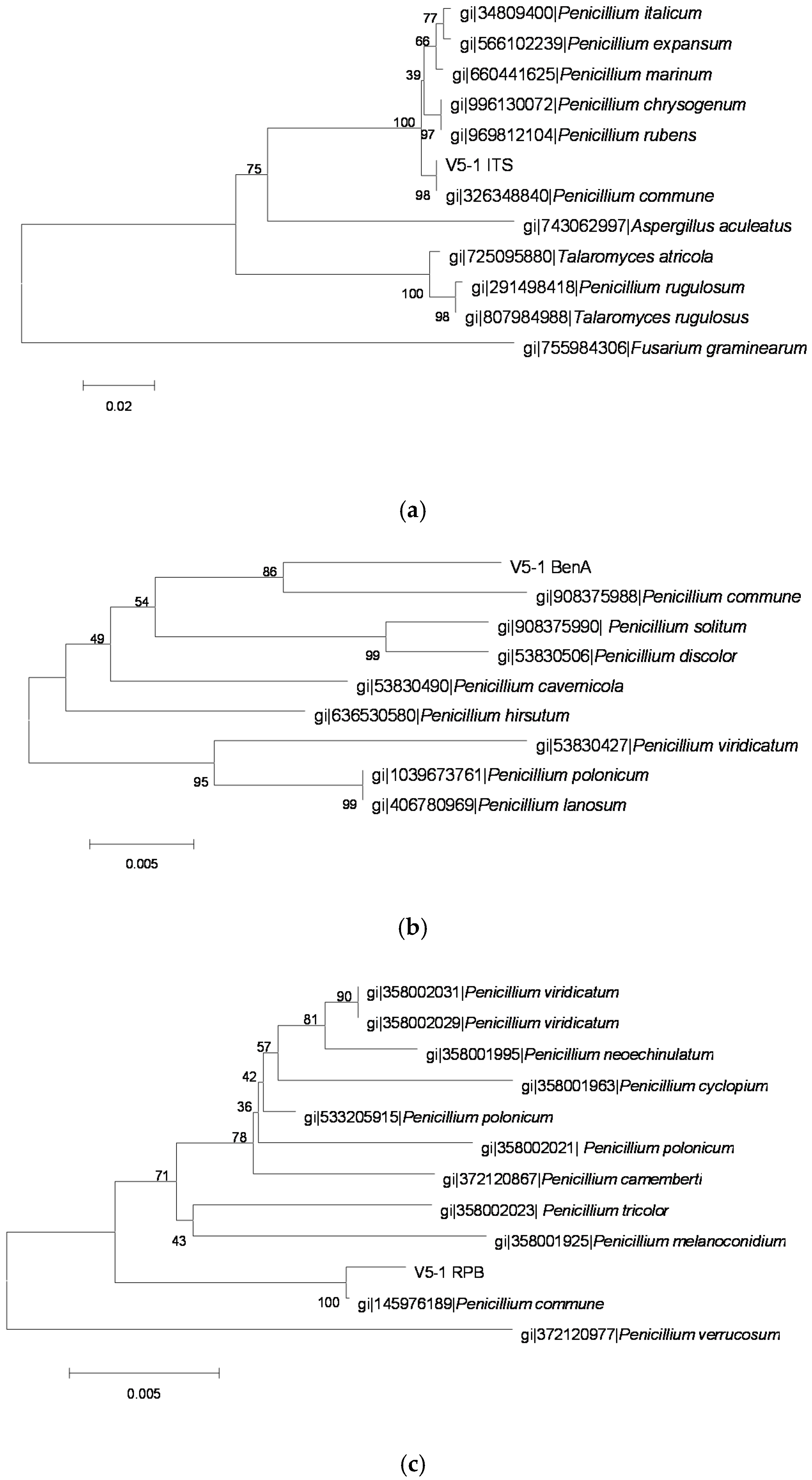
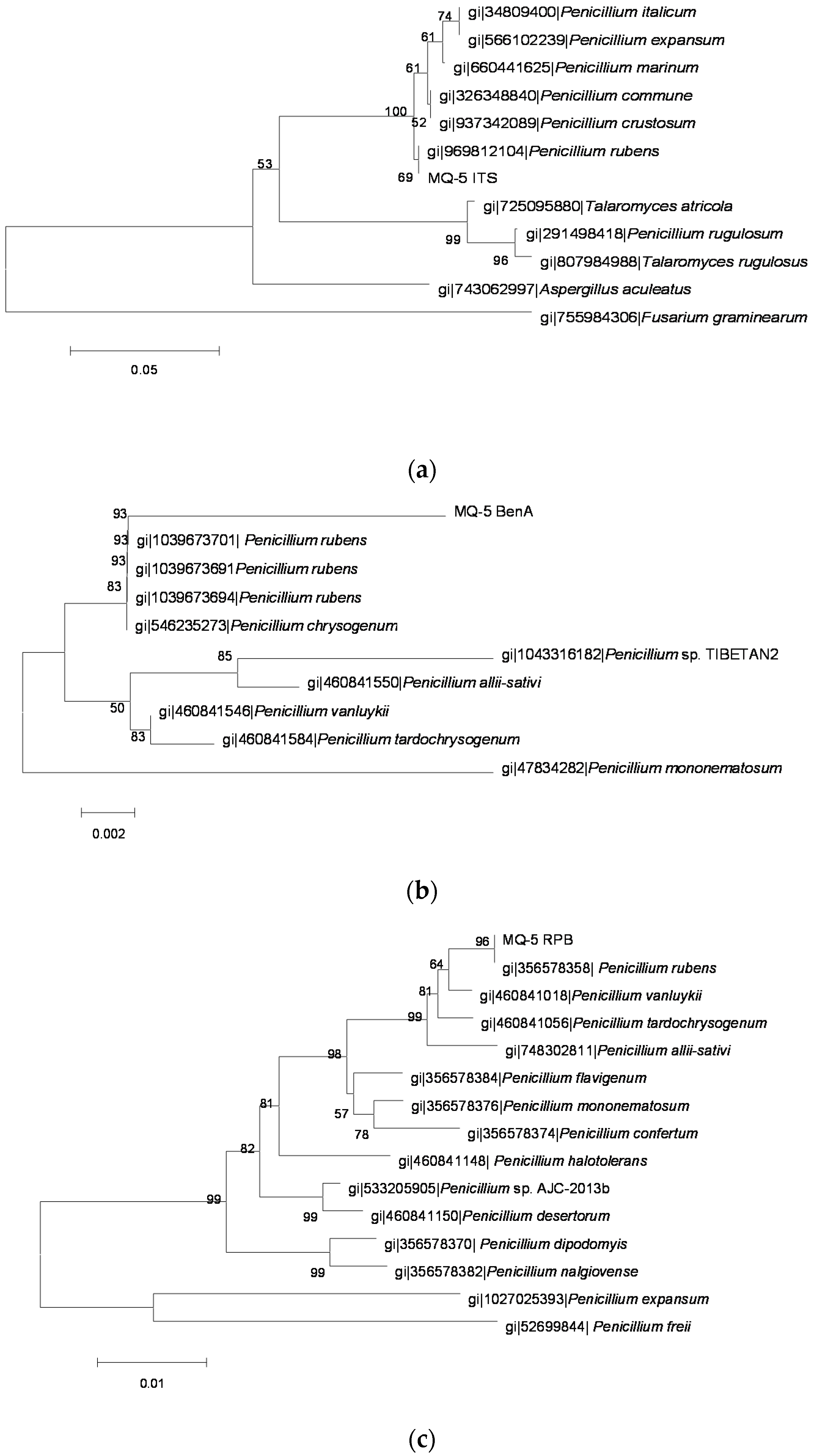
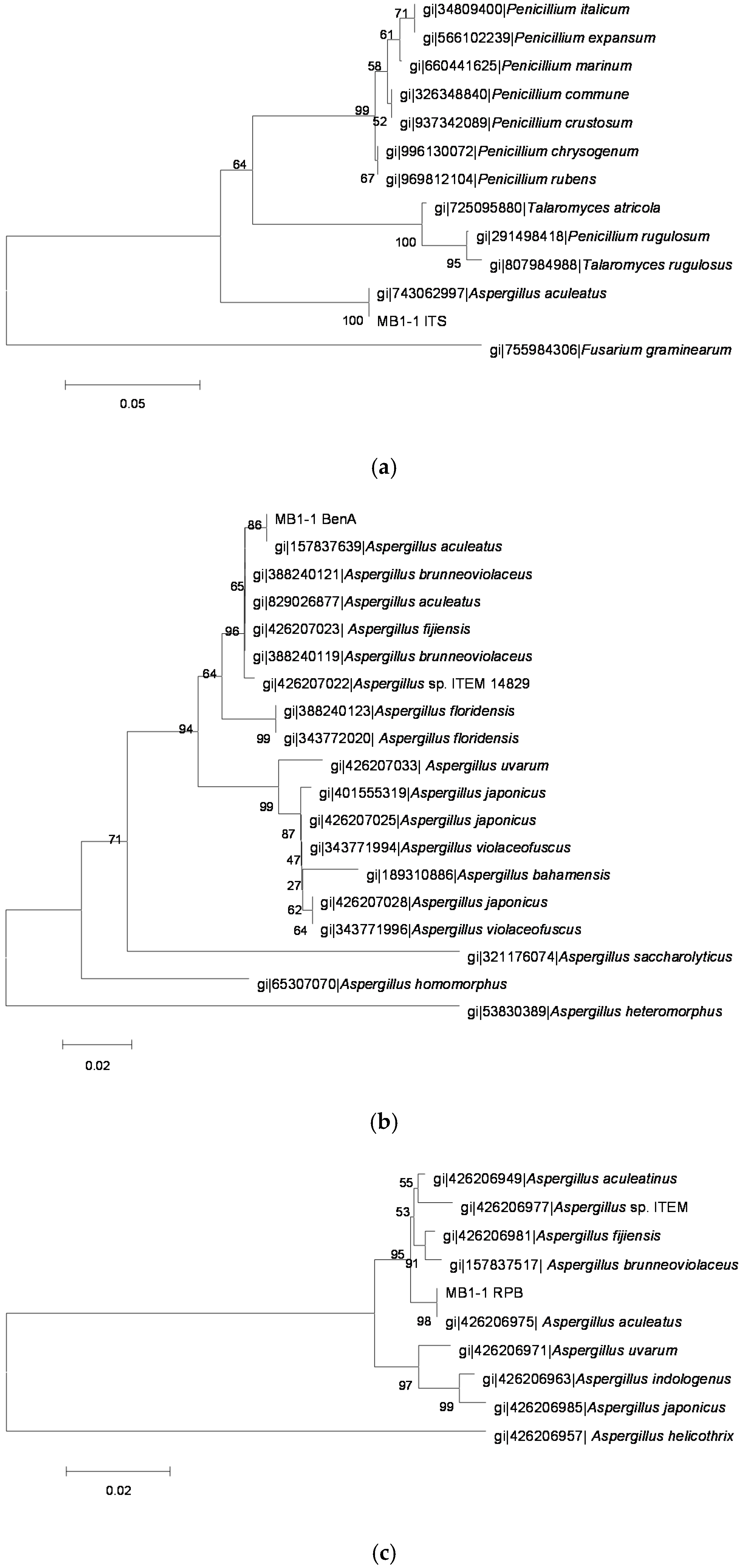
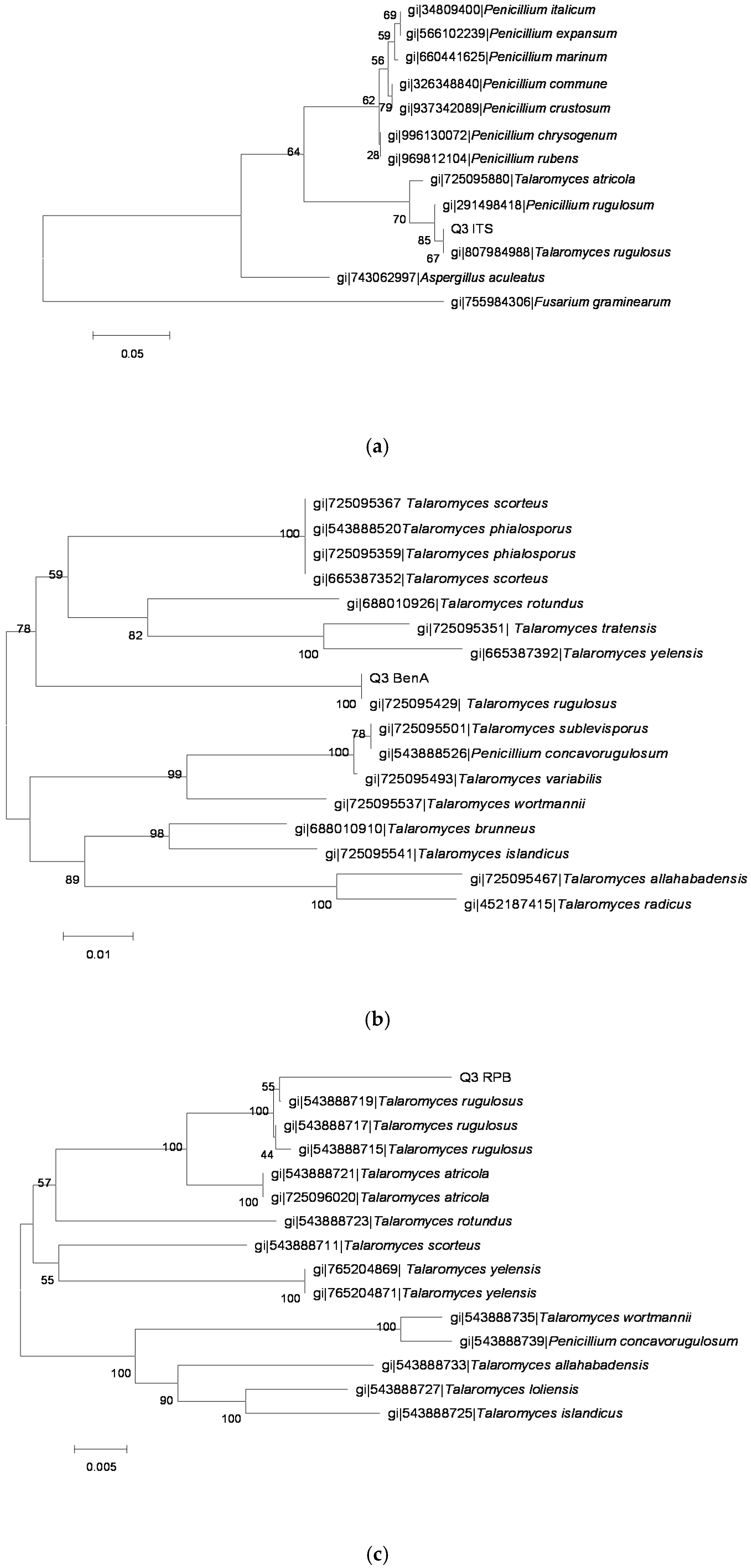
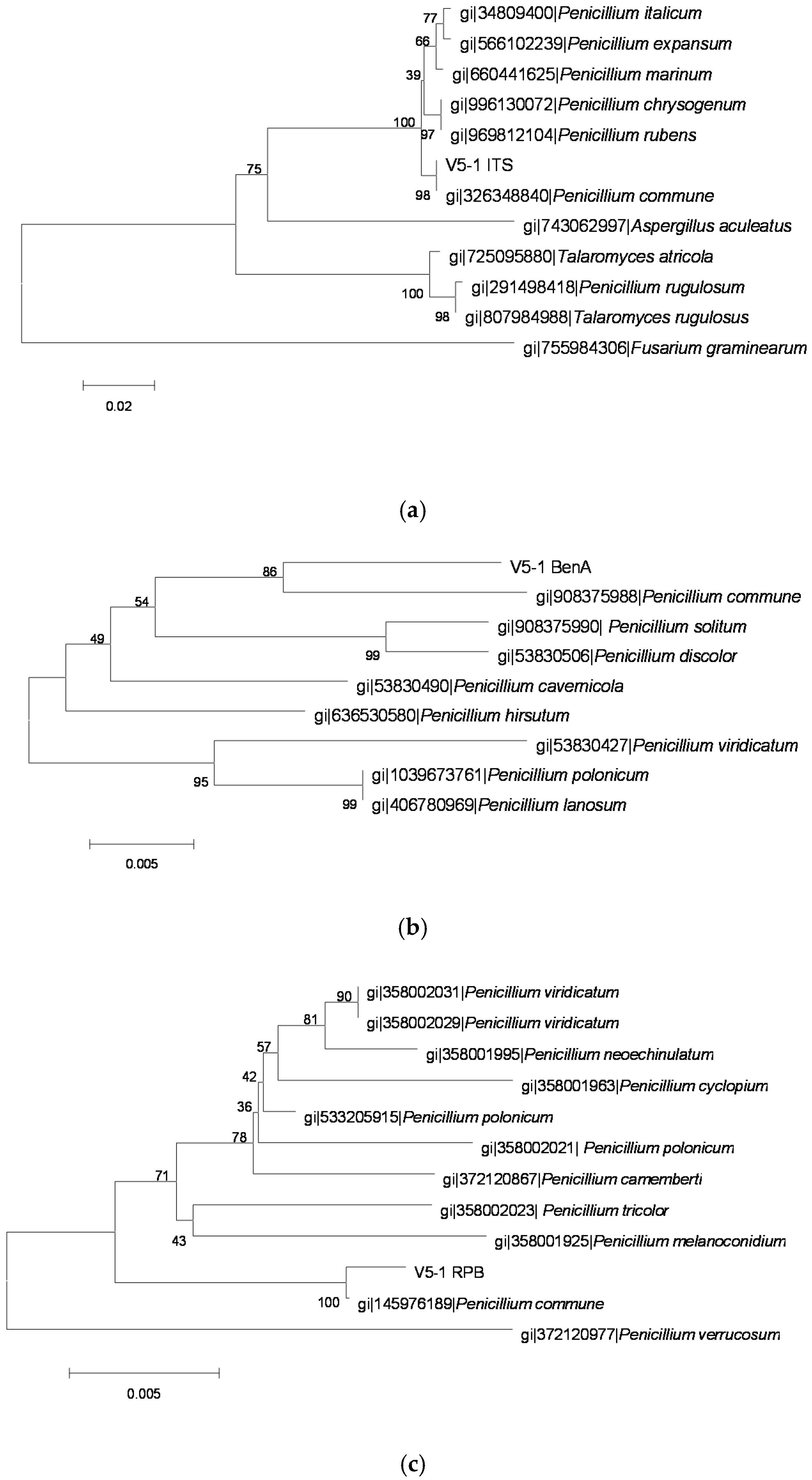
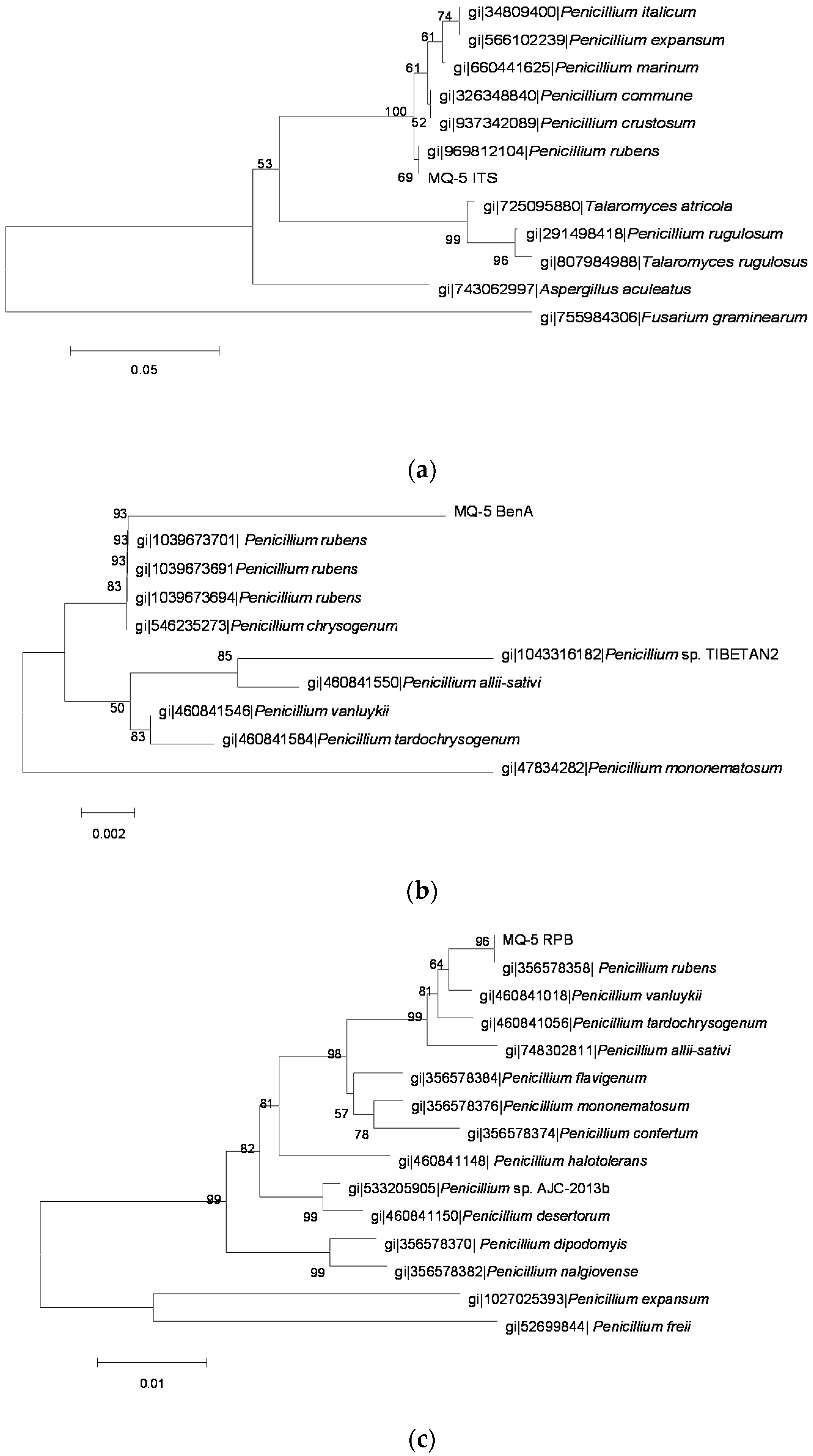
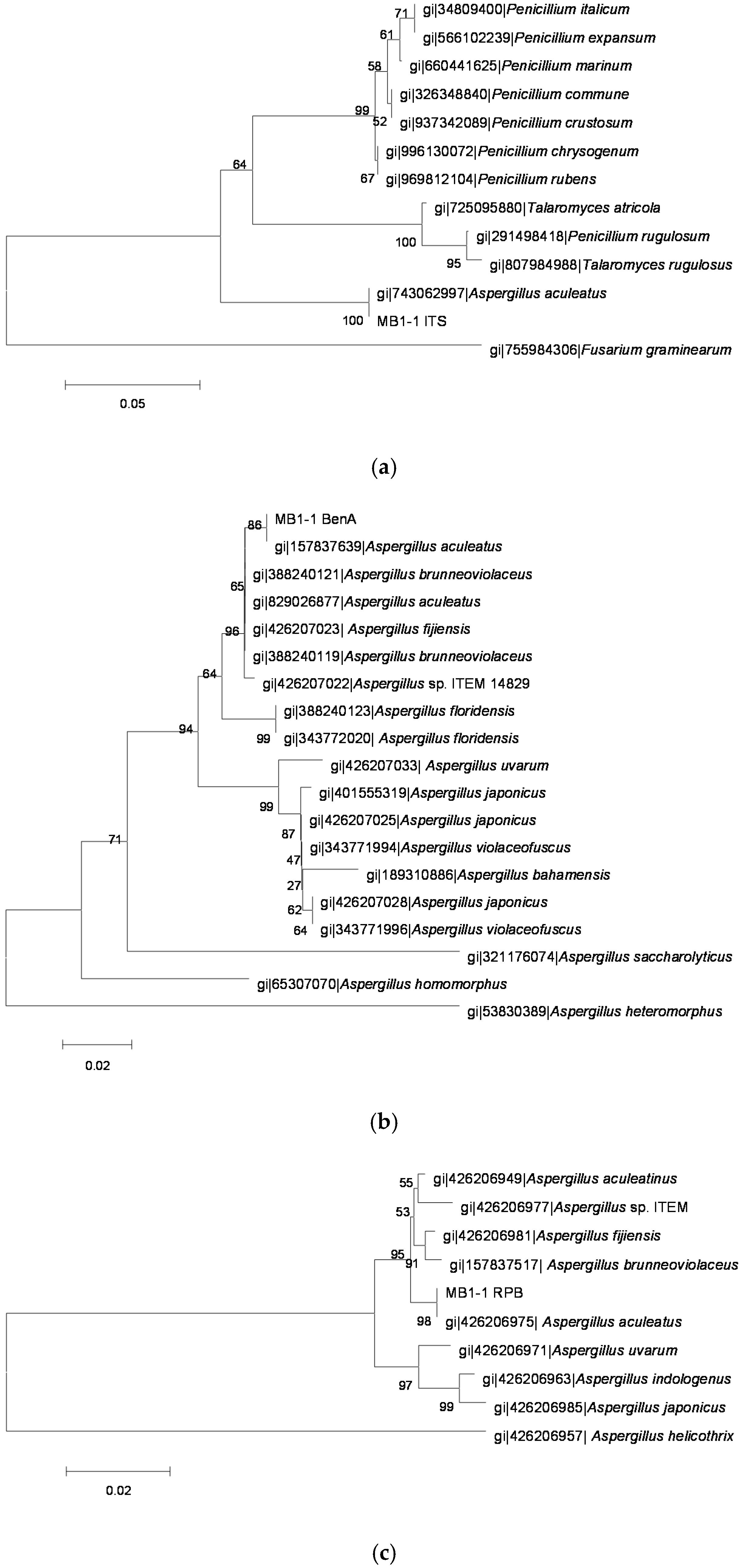
2.3. Identification of OTA-Producing Fungi
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Samples
5.2. Culture Medium
5.3. Isolation of Fungi from Decay Grapes
5.4. Preliminary Screening of Potential OTA-Producing Isolates by Ultraviolet Light
5.5. Confirmation of OTA-Producing Strains by High-Pressure Liquid Chromatography with a Fluorescence Detector (HPLC-FLD)
5.6. Identification of OTA-Producing Fungi
5.6.1. Genomic DNA Extraction
5.6.2. PCR Amplification and Sequencing
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gonzalez, G.; Hinojo, M.J.; Mateo, R.; Medina, A.; Jimenez, M. Occurrence of mycotoxin producing fungi in bee pollen. Int. J. Food Microbiol. 2005, 105, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Van der Merwe, K.J.; Stein, P.S.; Fourie, L. Ochratoxin A, a toxic metabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar] [CrossRef] [PubMed]
- Van Walbeek, W.; Scott, P.M.; Harwig, J.; Lawrence, J.W. Penicillium viridicatum Westling: A new source of ochratoxin A. Can. J. Microbiol. 1969, 15, 1281–1285. [Google Scholar] [CrossRef] [PubMed]
- Covarelli, L.; Beccari, G.; Marini, A.; Tosi, L. A review on the occurrence and control of ochratoxigenic fungal species and ochratoxin A in dehydrated grapes, non-fortified dessert wines and dried vine fruit in the Mediterranean area. Food Control 2012, 26, 347–356. [Google Scholar] [CrossRef]
- Suarez-Quiroz, M.; Gonzalez-Rios, O.; Barel, M.; Guyot, B.; Schorr-Galindo, S.; Guiraud, J.P. Effect of the post-harvest processing procedure on OTA occurrence in artificially contaminated coffee. Int. J. Food Microbiol. 2005, 103, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Ramyaa, P.; Krishnaswamy, R.; Padma, V.V. Quercetin modulates OTA-induced oxidative stress and redox signalling in HepG2 cells-up regulation of Nrf2 expression and down regulation of NF-kappaB and COX-2. Biochim. Biophys. Acta 2014, 1840, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Abrunhosa, L.; Venâncio, A.; Teixeira, J.A. Optimization of process parameters for the production of an OTA-hydrolyzing enzyme from Aspergillus niger under solid-state fermentation. J. Biosci. Bioeng. 2011, 112, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Heenan, C.N.; Shaw, K.J.; Pitt, J.I. Ochratoxin A production by Aspergillus carbonarius and A. niger isolates and detection using coconut cream agar. J. Food Mycol. 1998, 1, 67–72. [Google Scholar]
- Horie, Y. Productivity of ochratoxin A of Aspergillus carbonarius in Aspergillus section Nigri. Nippon Kingakkai Kaiho 1995, 36, 73–76. [Google Scholar]
- Nakajima, M.; Tsubouchi, H.; Miyabe, M.; Ueno, Y. Survey of aflatoxin B1 and ochratoxin A in commercial green coffee beans by high-performance liquid chromatography linked with immunoaffinity chromatography. Food Agric. Immunol. 1997, 9, 77–83. [Google Scholar] [CrossRef]
- Ono, H.; Kataoka, A.; Koakutsu, M.; Tanaka, K.; Kawasugi, S.; Wakazawa, M.; Ueno, Y.; Manabe, M. Ochratoxin A producibility by strains of Aspergillus niger group stored in IFO culture collection. Mycotoxins 1995, 41, 47–51. [Google Scholar] [CrossRef]
- Perrone, G.; Mulè, G.; Susca, A.; Battilani, P.; Pietri, A.; Logrieco, A. Ochratoxin A production and AFLP analysis of Aspergillus carbonarius, Aspergillus tubingensis, and Aspergillus niger strains isolated from grapes in Italy. Appl. Environ. Microbiol. 2006, 72, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Téren, J.; Varga, J.; Hamari, Z.; Rinyu, E.; Kevei, F. Immunochemical detection of ochratoxin A in black Aspergillus strains. Mycopathologia. 1996, 134, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Kocsubé, S.; Tóth, B.; Frisvad, J.C.; Perrone, G.; Susca, A.; Merger, M.; Samson, R.A. Aspergillus brasiliensis sp. nov. a biseriate black Aspergillus species with worldwide distribution. Int. J. Syst. Evol. Microbiol. 2007, 57, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C. The connection between the Penicillia and Aspergilli and mycotoxins with special emphasis on misidentified isolates. Arch. Environ. Contam. Toxicol. 1989, 18, 452–467. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Filtenborg, O. Terverticillate Penicillia: Chemotaxonomy and mycotoxin production. Mycologia 1989, 81, 837–861. [Google Scholar] [CrossRef]
- Pitt, J.I. Penicillium viridicatum, Penicillium verrucosum and production of ochratoxin A. Appl. Environ. Microbiol. 1987, 53, 535–539. [Google Scholar]
- Ciegler, A.; Fennell, D.J.; Mintzlaff, H.J.; Leistner, L. Ochratoxin synthesis by Penicillium species. Sci. Nat. 1972, 59, 365–366. [Google Scholar] [CrossRef]
- Krogh, P.; Hald, B.; Pedersen, E.J. Occurrence of ochratoxin A and citrinin incereals associated with mycotoxic porcine nephropathy. Acta. Pathol. Microbiol. Scand. 1973, 81, 689–695. [Google Scholar]
- Larsen, T.O.; Svendsen, A.; Smedsgaard, J. Biochemical characerization of ochratoxin A-producing strains of the genus Penicillium. Appl. Environ. Microbiol. 2001, 67, 3630–3635. [Google Scholar] [CrossRef] [PubMed]
- Bellí, N.; Pardo, E.; Marín, S.; Farré, G.; Ramos, A.J.; Sanchis, V. Occurrence of ochratoxin A and toxigenic potential of fungal isolates from Spanish grapes. Appl. Environ. Microbiol. 2004, 84, 541–546. [Google Scholar] [CrossRef]
- Duarte, S.C.; Pena, A.; Lino, C.M. A review on ochratoxin A occurrence and effects of processing of cereal and cereal derived food products. Food Control 2010, 27, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Kuiper-Goodman, T.; Scott, P.M. Risk assessment of the mycotoxin ochratoxin A. Biomed. Environ. Sci. 1989, 2, 179–248. [Google Scholar] [PubMed]
- Jφrgensen, K. Survey of pork, poultry, coffee, beer and pulses for ochratoxin A. Food Addit. Contam. 1998, 15, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Ueno, Y. Residue and risk of ochratoxin A in human plasma and beverages in Japan. Mycotoxins 1998, 47, 25–32. [Google Scholar] [CrossRef]
- Skaug, M.A. Analysis of Norwegian milk and infant formulas for ochratoxin A. Food Addit. Contam. 1999, 16, 75–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdaspal, P.A.; Legarda, T.M. Ocratoxina A in wines and grape products originated from Spain and other European countries. Alimentaria 1999, 36, 107–112. [Google Scholar]
- Covarelli, L.; Tosi, L.; Beccari, G. Risks related to the presence of fungal species and mycotoxins in grapes, wines and other derived products in the mediterranean area. In The Mediterranean Diet: An. Evidence-Based Approach; Preddy, V.R., Watson, R.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 563–575. [Google Scholar]
- Pardo, E.; Marín, S.; Sanchis, V.; Ramos, A.J. Impact of relative humidity and temperature on visible fungal growth and OTA production of ochratoxigenic Aspergillus ochraceus isolates on grapes. Food Microbiol. 2005, 22, 383–389. [Google Scholar] [CrossRef]
- Zimmerli, B.; Dick, R. Ochratoxin A in table wines and grape-juice: Occurrence and risk assessment. Food Addit. Contam. 1996, 13, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Romani, S.; Sachetti, G.; López, C.C.; Pinnavia, G.G.; Rosa, M.D. Screening on the occurrence of chratoxin A in green coffee beans of different origins types. J. Agric. Food Chem. 2000, 48, 3616–3619. [Google Scholar] [CrossRef] [PubMed]
- Taniwaki, M.H.; Pitt, J.I.; Teixeira, A.A.; Iamanaka, B.T. The source of ochratoxin A in Brazilian coffee and its formation in relation to process methods. Int. J. Food Microbiol. 2003, 82, 173–179. [Google Scholar] [CrossRef]
- Stander, M.A.; Steyn, P.S. Survey of ochratoxin A in South African wines. S. Afr. Jenol. Vitic. 2002, 23, 9–13. [Google Scholar]
- Lund, F.; Filtenborg, O.; Frisvad, J.C. Associated mycoflora of cheese. Food Microbiol. 1995, 12, 173–180. [Google Scholar] [CrossRef]
- Ciegler, A. Bioproduction of ochratoxin A and penicillic acid by membersof the Aspergillus ochraceus group. Can. J. Microbiol. 1972, 18, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.I.; Cruickshank, R.H.; Leistner, L. Penicillium Commune, P. camembertii, the origin of white cheese moulds, and the production of cyclopiazonic acid. Food Microbiol. 1986, 3, 363–371. [Google Scholar] [CrossRef]
- Houbraken, J.; Frisvad, J.C.; Samson, R.A. Fleming’s penicillin producing strain is not Penicillium Chrysogenum but P. Rubens. IMA Fungus 2011, 2, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, S.W.; Jurjević, Ž. Talaromyces columbinus sp. nov., and Genealogical Concordance Analysis in Talaromyces Clade 2a. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Tatsuno, T.; Kobayashi, N.; Okubo, K.; Tsunoda, H. Recherches Toxicologiques sur les Substances Toxiques de Penicillium tardum I. Isolement et Identification des Substances. Chem. Pharm. Bull. 1975, 23, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Iizuka. Aspergillus aculeatus. J. Agric. Food Chem. Soc. 1953, 27, 807. [Google Scholar]
- Andersen, R.; Buchi, G.; Kobbe, B.; Demain, A.L. Secalonic acids D and F are toxic metabolites of Aspergillus aculeatus. J. Org. Chem. 1977, 42, 352–353. [Google Scholar]
- Perrone, G.; Varga, J.; Susca, A. Aspergillus uvarum sp. nov., an uniseriate black Aspergillus species isolated from grapes in Europe. Int. J. Syst. Evol. Microbiol. 2008, 58, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Bau, M.; Castellá, G.; Bragulat, M.R.; Cabañes, F.J. DNA-based characterization of ochratoxin-A-producing and non-producing Aspergillus carbonarius strains from grapes. Res. Microbiol. 2005, 156, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Bau, M.; Bragulata, M.R.; Abarca, M.L.; Minguezb, S.; Cabañes, F.J. Ochratoxigenic species from Spanish wine grapes. Int. J. Food Microbiol. 2005, 98, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.Y.; Sh, J.L.; Jiang, C.M.; Liu, Y.L. Inhibition of the growth and ochratoxin A production by Aspergillus carbonarius and Aspergillus ochraceus in vitro and in vivo through antagonistic yeasts. Food Control 2015, 50, 125–132. [Google Scholar] [CrossRef]
- Dyer, S.K.; McCammon, S. Detection of aflatoxigenic isolates of Aspergillus flavusand related species on coconut cream agar. J. Appl. Bacteriol. 1994, 76, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Kizis, D.; Natskoulis, P.; Nychas, G.J.E.; Panagou, E.Z. Biodiversity and ITS-RFLP characterization of Aspergillus. Section Nigri isolates in grapes from four traditional grape-producing areas in Greece. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Cabañes, F.J.; Bragulat, M.R.; Castella, G. Characterization of nonochratoxigenic strains of Aspergillus carbonarius from grapes. Food Microbiol. 2013, 36, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Abarca, M.L.; Bragulat, M.R.; Cabañes, F.J. A new in vitro method to detect growth and ochratoxin A-producing ability of multiple fungal species commonly found in food commodities. Food Microbiol. 2014, 44, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Chiotta, M.L.; Ponsone, M.L.; Sosa, D.M.; Combina, M.; Chulze, S.N. Biodiversity of Aspergillus section Nigri populations in Argentinian vineyards and ochratoxin A contamination. Food Microbiol. 2013, 36, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Tannous, J.; Atoui, A.; El Khoury, A.; Kantar, S.; Chdid, N.; Oswald, I.P.; Puel, O.; Lteif, R. Development of a real-time PCR assay for Penicillium expansum quantification and patulin estimation in apples. Food Microbiol. 2015, 50, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguch, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultan, Y.; Magan, N. Mycotoxigenic fungi in peanuts from different geographic regions of Egypt. Mycotox. Res. 2010, 26, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noonim, P.; Mahakarnchanakul, W.; Varga, J.; Frisvad, J.C.; Samson, R.A. Two novel species of Aspergillus section Nigri from Thai coffee beans. Int. J. Syst. Evol. Microbiol. 2008, 58, 1727–1734. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | OTA Content (ng/g) |
|---|---|
| O1 | 97.5 ± 2.8 |
| Q3 | 50.11 ± 2.1 |
| V5-1 | 43.94 ± 1.8 |
| MQ-5 | 96.86 ± 3.2 |
| MB1-1 | 25.9 ± 1.9 |
| Gene | Profile Type | Initial Denaturing | Cycles | Denaturing Annealing Elongation | Final Elongation | Rest Period |
|---|---|---|---|---|---|---|
| ITS, BenA | standard | 94 °C, 10 min | 35 | 94 °C, 45 s; 55 °C, 45 s; 72 °C, 60 s | 72 °C, 10 min | 4 °C, ∞ |
| RPB | touch-up | 94 °C, 5 min | 5 | 94 °C, 45 s; 50 °C, 45 s; 72 °C, 60 s | - | - |
| - | - | - | 5 | 94 °C, 45 s; 52 °C, 45 s; 72 °C, 60 s | - | - |
| - | - | - | 30 | 94 °C, 45 s; 55 °C, 45 s; 72 °C, 60 s | 72 °C, 10 min | 4 °C, ∞ |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Li, Y.; Wang, H.; Gu, X.; Zheng, X.; Wang, Y.; Diao, J.; Peng, Y.; Zhang, H. Screening and Identification of Novel Ochratoxin A-Producing Fungi from Grapes. Toxins 2016, 8, 333. https://doi.org/10.3390/toxins8110333
Zhang X, Li Y, Wang H, Gu X, Zheng X, Wang Y, Diao J, Peng Y, Zhang H. Screening and Identification of Novel Ochratoxin A-Producing Fungi from Grapes. Toxins. 2016; 8(11):333. https://doi.org/10.3390/toxins8110333
Chicago/Turabian StyleZhang, Xiaoyun, Yulin Li, Haiying Wang, Xiangyu Gu, Xiangfeng Zheng, Yun Wang, Junwei Diao, Yaping Peng, and Hongyin Zhang. 2016. "Screening and Identification of Novel Ochratoxin A-Producing Fungi from Grapes" Toxins 8, no. 11: 333. https://doi.org/10.3390/toxins8110333