
Peptide Toxins in Solitary Wasp Venoms

1
Institute of Natural Medicine, University of Toyama, Toyama 930-0194, Toyama, Japan
2
Faculty of Agriculture, Utsunomiya University, Utsunomiya 321-8505, Tochigi, Japan
*
Author to whom correspondence should be addressed.
Toxins 2016, 8(4), 114; https://doi.org/10.3390/toxins8040114
Submission received: 30 January 2016
/
Revised: 5 April 2016
/
Accepted: 8 April 2016
/
Published: 18 April 2016
(This article belongs to the Special Issue Arthropod Venoms)
Abstract
:Solitary wasps paralyze insects or spiders with stinging venom and feed the paralyzed preys to their larva. Accordingly, the venoms should contain a variety of constituents acting on nervous systems. However, only a few solitary wasp venoms have been chemically studied despite thousands of species inhabiting the planet. We have surveyed bioactive substances in solitary wasp venoms found in Japan and discovered a variety of novel bioactive peptides. Pompilidotoxins (PMTXs), in the venoms of the pompilid wasps Anoplius samariensis and Batozonellus maculifrons, are small peptides consisting of 13 amino acids without a disulfide bond. PMTXs slowed Na+ channel inactivation, in particular against neuronal type Na+ channels, and were rather selective to the Nav1.6 channel. Mastoparan-like cytolytic and antimicrobial peptides are the major components of eumenine wasp venoms. They are rich in hydrophobic and basic amino acids, adopting a α-helical secondary structure, and showing mast cell degranulating, antimicrobial and hemolytic activities. The venom of the spider wasp Cyphononyx fulvognathus contained four bradykinin-related peptides. They are hyperalgesic and, dependent on the structure, differently associated with B1 or B2 receptors. Further survey led to the isolation of leucomyosuppressin-like FMRFamide peptides from the venoms of the digger wasps Sphex argentatus and Isodontia harmandi. These results of peptide toxins in solitary wasp venoms from our studies are summarized.
1. Introduction
The Hymenoptera is one of the largest orders among insects. It is composed of at least 200,000 species of bees, wasps and ants. Most of them have stinging venom, utilized for prey capture and self-defense. There are two distinct lifestyles among the Hymenopteran insects: social and solitary life. Social Hymenoptera includes honeybee, hornets, paper wasps and ants, whereas solitary Hymenoptera includes solitary wasps and parasitic wasps.
Social bees and wasps use their venom for defending themselves and their colonies from attacks by enemies and predators. Stinging with these venoms produces local pain and damage, and occasionally death, in large vertebrates including man. The chemical constituents of these venoms were well documented many years ago: biogenic amines, peptides and proteins act together to produce the toxic and biological effects [1,2]. In contrast, solitary wasps offensively use their venoms for prey capture. They inject their venoms to insects or spiders and paralyze the prey to feed their larvae. Therefore, the solitary wasp venoms should contain neurotoxins acting on nervous systems, ion channels and neuronal receptors in some ways [3]. Until recently, however, only a few studies have been reported for chemical components in solitary wasp venoms. It may be because of the extreme difficulty in collecting the number of specimens required to get a sufficient amount of venom to do bioassay-guided fractionation and chemical analysis due to their solitary lifestyle.
Pioneering studies demonstrated that neurotoxins are indeed contained in solitary wasp venoms. The first neurotoxic component characterized in solitary wasp venom was kinins. In 1987, Piek et al. isolated threonine6-bradykinin (Thr6-BK) and megascoliakinin (Thr6-BK-Lys-Ala) from the venoms of the European scoliid wasps Megascolia flavifrons and Colpa interrupta [4,5]. These kinins irreversibly block the synaptic transmission of the nicotinic acetylcholine receptor (nAChR) in the insect central nervous system [5,6]. Shortly thereafter, philanthotoxins (PhTXs) were found in the venom of the African digger wasp Philanthus triangulum [7,8]. They are acylpolyamine toxins and non-competitive antagonists of the glutamate receptor (GluR) and nAChR [9,10].
We collected solitary wasps found in Japan, and surveyed bioactive compounds in their venom, focusing on small molecules and peptides. We first found the novel peptide neurotoxins pompilidotoxins (PMTXs) in pompilid wasp venoms [11,12], and further survey on other venoms led to the isolation of antimicrobial and cytolytic peptides, a novel type of bradykinin-related peptides and FMRFamide peptides. Thus, our studies indicated that solitary wasp venom might contain a variety of bioactive peptides besides neurotoxins, which could be potentially useful for drug discovery. In this mini-review, we summarize the results from our studies on peptide toxins in solitary wasp venoms.
2. Pompilidotoxins
The spider wasp Anoplius samariensis, hunting only spiders as shown in Figure 1, is one of the most abundant species among the solitary wasps inhabiting in Japan. We isolated a peptide neurotoxin, α-PMTX, from the venom of this wasp by bioassay-guided fractionation using a lobster leg neuromuscular junction. The venom of another spider wasp, Batozonellus maculifrons, also contained α-PMTX together with the closely related β-PMTX, in which the Lys residue at position 12 was replaced by Arg [11]. Both toxins greatly facilitated neurotransmission in the lobster leg neuromuscular junction [13], and β-PMTX is five times more potent than α-PMTX. Structure-activity relationships studies using a number of synthetic analogs of α-PMTX with the lobster neuromuscular synapse showed that the basic amino acids at positions 1, 3 and 12 were important, and the potency was enhanced three to five times when they were replaced by the other, Lys–Arg or Arg–Lys. Furthermore, the C-terminal amide structure and the length of the 13 amino acids are essential since the C-terminal carboxyl analog and any analogs with shorter amino acid lengths were reduced or lost activity [14].
PMTXs act not only on crustacean neuromuscular synapses but also on mammalian nervous systems such as rat cortical neurons and hippocampal neurons [15,16,17]. Electrophysiological experiments revealed in detail that PMTX blocks voltage-gated sodium channels (VGSC) by slowing inactivation [18,19], and it discriminates between neuronal and cardiac VGSC, acting only on neuronal Na channels [20]. Accordingly, we conducted a thorough study of the effects of PMTX on VGSC isoforms, including seven mammalian VGSC isoforms (Nav1.1—Nav1.7) and one insect VGSC (DmNav1), by evaluating the sodium current inactivation [21]. No effects were seen for Nav1.4 and Nav1.5, moderate effects were seen for Nav1.1—Nav1.3 and Nav1.7, and the most potent effect was seen for Nav1.6; in other words, it is rather selective to Nav1.6. The effects for the insect VGSC DmNav1 were far more potent than that for Nav1.6, which is reasonable since the stinging target of solitary wasps is mostly insects.
Toxins targeting sodium channels similar to PMTXs are widely distributed in animal venoms [22], in particular contained as major toxins in sea anemone, scorpion and spider venoms [22,23,24,25,26]. These toxins play an important role in prey capture and self-defense. On the other hand, they have been used as research tools for neuronal functions, such as elucidating action mechanisms of ion channels or characterizing receptor functions, and some of them are of clinical interest for neurological disorders. PMTXs are small peptides consisting of only 13 amino acids without a disulfide bond, which is in marked contrast to other toxins with 40–60 amino acids and three to four disulfide bonds. Therefore, PMTXs may also be useful for research tools and drug discovery [27,28,29,30,31,32].
3. Antimicrobial and Cytolytic Peptides
The Eumenine wasp Anterhynchium flavomarginatum micado may be the most common solitary wasp in Japan, hunting only green caterpillars and building their nests in bamboo twigs with mud. The major component of this venom, eumenine mastoparan-AF (EMP-AF), is a peptide structurally related to mastoparan [33]. Mastoparan was initially isolated from hornet venom, then closely related peptides were found in a variety of social wasp venoms (hornets and paper wasps) [2]. Accordingly, they are collectively called mastoparans or mastoparan-like peptides. The mastoparans are 14 amino acids in length with an amidated C-terminus, and they are rich in hydrophobic and basic amino acids, which makes these peptides have an amphipathic chemical character, adopting an α-helical secondary structure under proper conditions. This chemical feature is essential for their biological activities through the cell membrane, and accordingly, they are also called cytolytic peptides. EMP-AF has all the chemical characteristics of mastoparans, and, in fact, showed high sequence similarity to mastoparan [33] and a high content of α-helical conformation under SDS (sodium dodecyl sulfate) micelle and TFE (trifluoroethanol) conditions by circular dichroism (CD) spectral analysis [34]. NMR analysis supported the CD results, and also showed that the C-terminal amide stabilized the α-helical conformation [35]. In biological evaluation, EMP-AF stimulated degranulation from rat peritoneal mast cells and RBL-2H3 cells to an extent similar to mastoparan, and showed significant hemolytic activity in human erythrocytes. Antimicrobial activity was also investigated because cytolytic peptides are known to show antimicrobial activity. EMP-AF exhibited potent growth inhibition against Gram-positive bacteria as compared to Gram-negative bacteria, for example IC50 was 5 g/mL to Staphylococcus aureus ATCC6538 [34].
Eumenine solitary wasps may have such antimicrobial and cytolytic peptides in common, but their structures and biological activities are somewhat different depending on the species. Those discovered so far are summarized in Table 1. Eumenine mastoparan-OD (EMP-OD) is the second mastoparan to be found in the Eumenine wasp venom of Orancistrocerus drewseni drewseni. This novel peptide has typical chemical features of mastoparan, and exhibits more potent hemolytic activity than that of mastoparan [36,37]. Eumenitin, from Eumenes rubronotatus, is 15 amino acids in length with an extra hydrophilic amino acid at the C-terminus and is not amidated; it still has basically the same chemical features as those of mastoparans. It showed the same biological activities as mastoparans, but its potency tends to be lower [38,39]. Three other eumenine wasp venoms have both mastoparan-type and Eumenitin-type peptides together: EMP-ER and Eumenitin-R from Eumenes rubrofemoratus; EMP-EF and eumenitin-F from Eumenes fraterculus [40]; EpVP2a and EpVP1 from Eumenes pomiformis [41]. Eumenitin-type peptides have only been contained in solitary wasp venoms, and never found in social wasp venoms. Decoralin from Oreumenes decorates is a smaller peptide, being only 11 amino acids in length, but the chemical features can be classified into Eumenitin-type. In fact, it showed typical biological activities of cytolytic peptides [42].
In addition to Eumenine wasps, an antimicrobial and cytolytic peptide has been found in the spider wasp. Anoplin was isolated as a minor component from Anoplius samariensis venom, which contained α-PMTX as a major component [43]. This peptide has only 10 amino acids, but both chemical and biological features were typical for that of cytolytic peptides [44]. Interestingly, the similarities and differences of anoplin vs. decoralin correspond to those of mastoparan vs. eumenitin. Anoplin is the smallest peptide among the antimicrobial and cytolytic peptides found from natural sources, which may make it a lead compound for developing new, potent and useful antimicrobial compounds [45,46,47,48].
Antimicrobial and cytolytic peptides are widely found in many other arthropod venoms such as spider and scorpion venoms [49]. The majority of these peptides are linear, cationic and amphipathic in character, but the amino acid length is variable from 15 to 60 residues. They may play a dual role in acting as antimicrobials and potentiating the venom toxicity by disturbing excitable membranes. It can also be the case for the solitary wasp venoms [50]. Antimicrobials are injected beforehand for protection against microbial infection when larvae consumes the paralyzed prey.
4. Bradykinin-Related Peptides
In 1987, the first structure of a solitary wasp toxin was reported. Two bradykinin-related peptides, threonine6-bradykinin (Thr6-BK) and megascoliakinin (MBK), were isolated from the venoms of the European scoliid wasps Megascolia flavifrons and Colpa interrupta [4,5], as mentioned in the Introduction. The first toxin found in social wasp venoms was also a bradykinin-related peptide found as a pain-producing substance in 1954, and since then, many bradykinin-related peptides, collectively called wasp kinins, have been isolated and chemically characterized from Polistes and Vespid social wasp venoms [2]. Wasp kinins show similar properties to bradykinin (BK) both chemically and pharmacologically, containing a whole BK sequence elongated at the C- or N- terminal, and showing hypotensive and inflammatory activities. Piek et al. reported that Thr6-BK and megascoliakinin irreversibly block the synaptic transmission of the nicotinic acetylcholine receptor (nAChR) in the insect central nervous system, which indicates that these kinins play a major role in the paralyzing activity of the solitary wasp venoms [5,6].
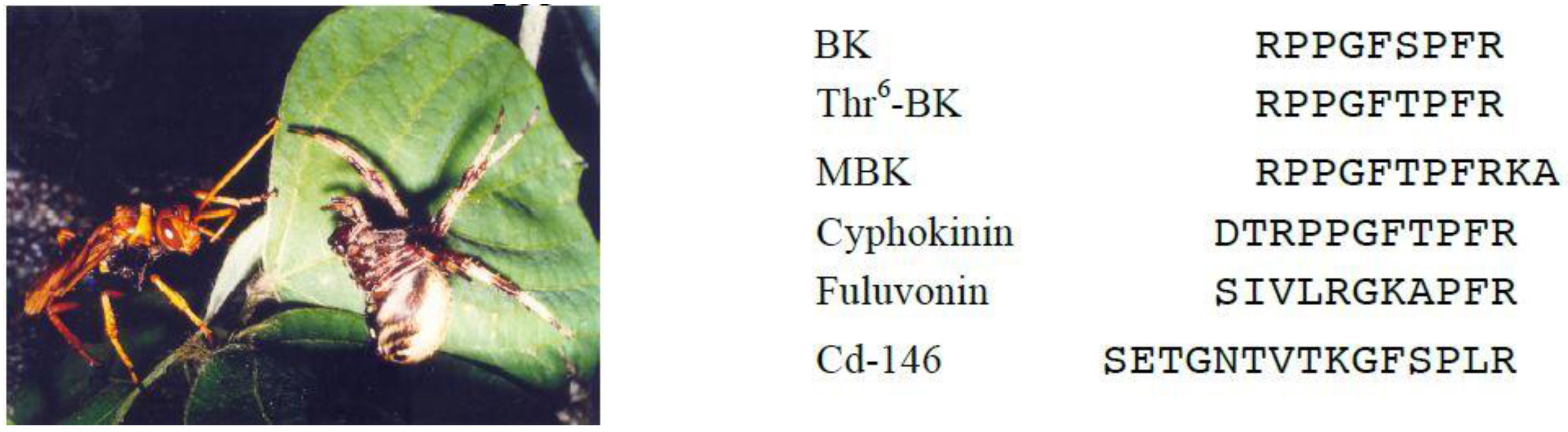
Piek et al. further investigated the pharmacological activities of solitary wasp venoms and found that scoliid wasp venoms commonly showed kinin activity [51]. We demonstrated the presence of Thr6-BK in the venoms of Japanese scoliid wasps [52]. Screening of 23 venom extracts from Japanese solitary wasps by MALDI-TOF MS revealed that scoliid wasp venoms of Megacampsomeris prismatica, Campsomeriella annulata annulata and Carinoscolia melanosoma fascinata contained Thr6-BK as one of the major components. This peptide was also identified in the venom extracts of the spider wasp Cyphononyx fluvognathus (formerly Cyphononyx dorsalis, Figure 2) [53]. This was the first example where a bradykinin-related peptide was found in spider wasp venom. We therefore were interested in this venom and analyzed the minor components carefully, which resulted in finding novel bradykinin-related peptides [54]. Cyphokinin may be classified into wasp kinins because it includes the whole sequence of Thr6-BK with only two extra amino acid residues elongating at its N-terminus. On the other hand, Cd-146 and fulvonin show a lesser similarity to the bradykinin sequence. Cd-146 has a kinin-like sequence at the C-terminal with six residues, while fulvonin has only three residues identical to BK. In pharmacological assays, only cyphokinin, including the whole sequence of BK, promoted the contraction of guinea pig ileum smooth muscle preparations, which was completely blocked by HOE-140, the B2 receptor antagonist, whereas only fulvonin potentiated BK-elicited smooth muscle contraction. In the rat paw pressure test after intraplantar injection, all the peptides showed a hyperalgesic effect with different strengths. This effect is due to the action of BK receptors, since the hyperalgesia induced by Cd-146 and fulvonin was blocked by the B1 receptor antagonist, while the effect of cyphokinin was reversed by the B2 antagonist. Thus, these BK-related peptides are a new type of wasp kinins and show distinct biological activities associated with BK receptors.
The component responsible for the paralytic activity of the C. dorsalis venom was reported to be a protein. Bioassay-guided fractionation led to the identification of an arginine-kinase-like protein with high homology to that of honeybee. Recombinant protein expressed in E. coli exhibited paralytic activity against spiders with the same characteristic symptoms as the crude venom [55]. Arginine-kinase-like protein was also found in the two Eumenine wasp venoms of Eumenes pomiformis and Orancistrocerus drewseni as the most predominant protein in both venoms [56]. The arginine-kinase of these solitary wasps had high sequence identities to those of C. dorsalis (95%–96%), and the honeybee A. mellifera (86%–88%).
5. FMRFamide-Related Neuropeptides
Digger wasps are a large group of solitary wasps, and they build a nest by digging a burrow in the ground. They have a large variety of hunting prey from spiders and crickets to flies and bees. Philanthus triangulum, inhabiting the Sahara desert in Africa, is the only species among digger wasps that have been chemically studied, and their major toxin is philanthotoxins-433 (PhTX-433) [7,8], as mentioned in the Introduction. This wasp toxin has been utilized for extensive studies on glutamatergic neurotransmission systems, resulting in the discovery of highly useful glutamate receptor antagonists [57].
Many digger wasps inhabit Japan, but they have never been studied until quite recently. We have chemically studied the venoms of two dominant species in Japan, Sphex argentatus argentatus and Isodontia harmandi, and found FMRFamide-related neuropeptides for the first time in solitary wasp venom [58]. Sphex argentatus argentatus is frequently found and is the largest in size among the Japanese solitary wasps. They sometimes make a colony, nesting in the same place, and hunt only grasshoppers as shown in Figure 3. From the venom extracts of this wasp, we isolated two closely related peptides, Sa-112 and Sa-12,both of which have the same 10-amino-acid sequence and the only difference is whether the C-terminal is amidated or not. Sa-112 was also found in the venom of the close relative Isodontia harmandi, from which Sh-5b, an N-terminal-missing analog of Sa-112, was obtained together. All these peptides should be classified into FMRFamide-related peptides because they bear the FMLF sequence at the C-terminal. The FMRFamide peptides are called due to their characteristic C-terminal sequence, and they are a group of neuropeptides, distributed widely in arthropods and in insects, involved in the regulation of physiological processes by acting as neurotransmitters, neurohormones and/or neuromodulation. These processes generally include activities such as digestion, circulation, and reproduction [59]. Sa-112 is highly homologous to leucomyosupressin [60] and SchistoFLRFamide [61], the FMRFamide-related neuropeptides from the cockroach and locust, respectively. Only the N-terminal amino acid was replaced to E (glutamic acid) from P (proline) or pQ (pyroglutamic acid), and accordingly, it is expected to show similar activities to those of the known neuropeptides. Indeed, Sa-112 inhibited contraction of the locust oviduct, a typical activity of FMRFamide peptides, at 50 nM to the same extent as SchistoFLRFamide, while Sa-12, a non-amidated peptide, showed no activity in the locust oviduct contraction. These results demonstrated again the essential role of the C-terminal amide structure to their biological activity. This is the first example of an FMRF-related neuropeptide to be found in solitary wasp venom. Another peptide, Sh-5b, was already synthesized for structure-activity relationship studies and was shown to have the same activity as that of SchistoFLRFamide.
It is interesting that these digger wasps inject FMRFamide neuropeptides into their prey, grasshoppers, because the grasshoppers have similar neuropeptides in their body and they are functioning in some way. Then, what is the role and effect of injected neuropeptides from wasp venom? They may not be directly involved in the paralyzing effect, but might play a supporting role in disturbing physiological regulation with the neuropeptide in the grasshoppers. In any case, it should be studied further.
6. Other Peptide Toxins
We have found other novel peptides in spider wasp venom. They are Cd-125 from Cyphononyx dorsalis [53], As-126 from Anoplius samariensis [62] and Bm-10 from Batozonellus maculifrons [62] as summarized in Table 2. The sequences were determined mainly by using mass spectrometric methods. In particular, Cd-125 is the major component of this wasp venom with a length of only 8 amino acids, but its biological activity remains to be studied. This is also the case for As-126 and Bm-10, the minor components of these venoms.
Orancis-Protonectin was isolated from the Eumenine wasp venom of Orancistrocerus drewseni drewseni together with EMP-OD [36,37]. The sequence is highly homologous to those of protonectins, hemolytic peptides isolated from social wasp venoms. In fact, Oricis-Protonectin showed more potent hemolytic activity than mastoparan. This is the first example of a protonectin-type peptide that was found in solitary wasp venom.
The presence of a higher molecular weight peptide, As-fr-19, in the Anoplius samariensis venom has been reported [63]. It was found in a fraction responsible for spider paralysis. This peptide is a novel multiple-cysteine peptide with high sequence similarity to known peptides such as dendrotoxins (K+ channel blocker) and Kuniz-type protease inhibitor. However, its biological properties are not characterized yet because it is still a component of a fraction (a mixture of several peptides). A dendrotoxin-like sequence was also found in the transcripts from the Eumenine wasp venoms [41].
7. Concluding Remarks
The major role of solitary wasp venom is to paralyze the prey and serve it to the wasps’ larva as food. Therefore, the solitary wasp venoms should contain neurotoxins acting on a variety of nervous systems. Our studies indicated that solitary wasp venoms might contain a variety of bioactive peptides besides neurotoxins, such as antimicrobial and cytolytic peptides, bradykinin-related peptides and neuropeptides. As discussed by Lee and co-workers [64], there are similarities and differences between solitary and social wasp toxins. Antimicrobial and cytolytic peptides and bradykinin-related peptides are common, but pompilidotoxins and FMRFamide-related neuropeptides are distinct to solitary wasp venoms. The prey of solitary wasps is specific to the class of wasps, for example pompilido wasps prey only on spiders, whereas eumenine wasps only on caterpillars. The reasons for this specificity are not known yet, but it may indicate that distinct components are included in the solitary wasp venoms depending on the species and/or class of wasps. Therefore, more diverse bioactive substances would be found if more solitary wasp venoms were studied. On the other hand, those solitary wasp toxins would be potentially useful not only for research tools but also for drug discovery, along with other venom toxins from snakes, scorpions, spiders and cone snails. It is noteworthy that the structure of these peptide toxins is relatively simple: only 8–16 amino acids in length without a disulfide bond, which is advantageous for practical use and development. Among more than 20,000 solitary wasps inhabiting in the planet, only about 10 species have been studied thus far. It may be because of the extreme difficulty in collecting a sufficient amount of the venom; in other words, a large number of wasps is required for bioassay-guided fractionation and chemical analysis due to their solitary lifestyle. However, recent developments of chemical analysis, especially by mass spectrometry, have made it possible to analyze quite minute amounts of venom components [53,62,65]. Quite recently, we established that, with using LC-MS, only 10% of the amount of crude venom from a single wasp specimen is sufficient to analyze most of the peptide toxins (peptidomic analysis). LC-MS/MS analysis allows de novo sequencing of most of the peptides, sometimes with more than 100 components, contained in the crude venom [66]. Biological activities would be evaluated by using synthetic specimens of deduced sequences. This is in marked contrast to bioassay-guided fractionation, which gives only major and a few minor components. With this method in hand, we are further surveying solitary wasp venoms, which would lead to finding a variety of useful bioactive substances.
Acknowledgments
We are grateful to Prof. Sôichi Yamane (Ibaraki University), Prof. Akira Endo (Ritsumeikan University) and Mr. Manabu Kato (Yamada Apiculture Center, Inc.) for collection and identification of wasps and their invaluable discussions. A part of this work was finacially supported by the JSPS KAKENHI Grant Number 15K07805, Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) of Japan.
Author Contributions
Katsuhiro Konno, Kohei Kazuma and Ken-ichi Nihei conceived the contents and analyzed the data. Katsuhiro Konnowrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Banks, B.E.C.; Shipolini, R.A. Chemistry and pharmacology of honey-bee venom. In Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Piek, T., Ed.; Academic Press: London, UK, 1986; pp. 330–416. [Google Scholar]
- Nakajima, T. Pharmacological biochemistry of vespid venoms. In Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Piek, T., Ed.; Academic Press: London, UK, 1986; pp. 309–327. [Google Scholar]
- Piek, T.; Spanjer, W. Chemistry and pharmacology of solitary wasp venoms. In Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Piek, T., Ed.; Academic Press: London, UK, 1986; pp. 161–307. [Google Scholar]
- Yasuhara, T.; Mantel, P.; Nakajima, T.; Piek, T. Two kinins isolated from an extract of the venom reservoirs of the solitary wasp Megascolia flavifrons. Toxicon 1987, 25, 527–535. [Google Scholar] [CrossRef]
- Piek, T.; Hue, S.; Mantel, P.; Nakajima, T.; Pelhate, M.; Yasuhara, T. Threonine6-bradykinin in the venom of the wasp Colpa interrupta (F.) presynaptically blocks nicotinic synaptic transmission in the insect CNS. Comp. Biochem. Physiol. 1990, 96C, 157–162. [Google Scholar] [CrossRef]
- Piek, T.; Hue, B.; Mony, L.; Nakajima, T.; Pelhate, M.; Yasuhara, T. Block of synaptic transmission in insect CNS by toxins from the venom of the wasp Megascolia flavifrons (FAB.). Comp. Biochem. Physiol. 1987, 87C, 287–295. [Google Scholar] [CrossRef]
- Eldefrawi, A.T.; Eldefrawi, M.E.; Konno, K.; Mansour, N.A.; Nakanishi, K.; Oltz, E.; Usherwood, P.N.R. Structure and synthesis of a potent glutamate receptor antagonist in wasp venom. Proc. Natl. Acad. Sci. USA 1988, 85, 4910–4913. [Google Scholar] [CrossRef] [PubMed]
- Piek, T.; Fokkens, R.H.; Karst, H.; Kruk, C.; Lind, A.; van Marle, J.; Nakajima, T.; Nibbering, N.M.M.; Shinozaki, H.; Spanjer, W.; et al. Polyamine like toxins—A new class of pesticides? In Neurotox ′88: Molecular Basis of Drug and Pesticide Action; Lunt, G.G., Ed.; Elsevier: Amsterdam, The Netherlands, 1988; pp. 61–76. [Google Scholar]
- Nakanishi, K.; Goodnow, R.; Konno, K.; Niwa, M.; Bukownik, R.; Kalimopoulos, T.A.; Usherwood, P.N.R.; Eldefrawi, A.T.; Eldefrawi, M.E. Philanthotoxin-433 (PhTX-433), a non-competitive glutamate receptor inhibitor. Pure Appl. Chem. 1990, 62, 1223–1230. [Google Scholar] [CrossRef]
- Piek, T.; Hue, B. Philanthotoxins, a new class of neuroactive polyamines, block nicotinic transmission in the insect CNS. Comp. Biochem. Physiol. 1989, 93c, 403–406. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Itagaki, Y.; Naoki, H.; Kawai, N.; Miwa, A.; Yasuhara, T.; Takayama, H. Isolation and structure of pompilidotoxins (PMTXs), novel neurotoxins in solitary wasp venoms. Biochem. Biophys. Res. Commun. 1998, 250, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Kawai, N. Pompilidotoxins: Novel peptide neurotoxins blocking sodium channel inactivation from solitary wasp venom. Curr. Med. Chem. Central Nerv. Syst. Agents 2004, 4, 139–146. [Google Scholar] [CrossRef]
- Konno, K.; Miwa, A.; Takayama, H.; Hisada, M.; Itagaki, Y.; Naoki, H.; Yasuhara, T.; Kawai, N. α-Pompilidotoxin (α-PMTX), a novel neurotoxin from the venom of a solitary wasp, facilitates transmission in the crustacean neuromuscular synapse. Neurosci. Lett. 1997, 238, 99–102. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Yasuhara, T.; Nakata, Y.; Miwa, A.; Kawai, N. Molecular determinants of binding of a wasp toxin (PMTXs) and its analogs in the Na+ channels proteins. Neurosci. Lett. 2000, 285, 29–32. [Google Scholar] [CrossRef]
- Harsch, A.; Konno, K.; Takayama, H.; Kawai, N.; Robinson, H. Effects of α-pompilidotoxin on synchronized firing in networks of rat cortical neurons. Neurosci. Lett. 1998, 252, 49–52. [Google Scholar] [CrossRef]
- Miyawaki, T.; Tsubokawa, H.; Yokota, H.; Ogura, K.; Konno, K.; Masuzawa, T.; Kawai, N. Differential effects of novel wasp toxin on rat hippocampal interneurons. Neurosci. Lett. 2002, 328, 25–28. [Google Scholar] [CrossRef]
- Yokota, H.; Tsubokawa, H.; Miyawaki, T.; Konno, K.; Nakayama, H.; Masuzawa, T.; Kawai, N. Modulation of synaptic transmissions in hippocampal CA1 neurons by a novel neurotoxin (β-PMTX) derived from wasp venom. Neurosci. Res. 2001, 41, 365–371. [Google Scholar] [CrossRef]
- Sahara, Y.; Gotoh, M.; Konno, K.; Miwa, A.; Tsubokawa, H.; Robinson, H.P.C.; Kawai, N. A new class of neurotoxin from wasp venom slows inactivation of sodium current. Eur. J. Neurosci. 2000, 12, 1961–1970. [Google Scholar] [CrossRef] [PubMed]
- Kawai, N.; Konno, K. Molecular determinants of two neurotoxins that regulate sodium current inactivation in rat hippocampus. Neurosci. Lett. 2004, 361, 44–46. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, E.; Maejima, H.; Yamaoka, K.; Konno, K.; Kawai, N.; Shimizu, E.; Yokote, S.; Nakayama, H.; Seyama, I. Novel wasp toxin discriminates between neuronal and cardiac sodium channels. Mol. Pharmacol. 2001, 59, 1457–1463. [Google Scholar]
- Schiavon, E.; Stevens, M.; Zaharenko, A.J.; Konno, K.; Tytgat, J.; Wanke, E. Voltage-gated sodium channels isoform-specific effects of pompilidotoxins. FEBS J. 2010, 277, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, J.; Olivera, B.M.; Bosmans, F. Animal toxins influence voltage-gated sodium channel function. Handb. Exp. Pharmacol. 2014, 221, 203–229. [Google Scholar] [PubMed]
- Oliveira, J.S.; Redaelli, E.R.; Zaharenko, A.; Cassulini, R.R.; Konno, K.; Pimenta, D.C.; de Freitas, J.C.; Clare, J.J.; Wanke, E. Nav 1.1–1.6 sodium channels binding specificity to sea anemone toxins: Unexpected contributions from changes in the IV/S3-S4 outer loop. J. Biol. Chem. 2004, 279, 33323–33335. [Google Scholar] [CrossRef] [PubMed]
- Honma, T.; Shiomi, K. Peptide toxins in sea anemones: Structural and functional aspects. Marine Biotech. 2006, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Cerni, F.A.; Pinheiro, E.L., Jr.; Bordon, K.C.F.; Amorim, F.G.; Cordeiro, F.A.; Longhim, H.T.; Cremonez, C.M.; Oliveira, G.H.; Arantes, E.C. Tityus serrulatus venom—A lethal cocktail. Toxicon 2015, 108, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Bosmans, F.; Swartz, K.J. Targeting voltage sensors in sodium channels with spider toxins. Trends Pharmacol. Sci. 2010, 31, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Grieco, T.M.; Raman, I.M. Production of resurgent current in NaV1.6-null Purkinje neurons by slowing sodium channel inactivation with β-pompilidotoxin. J. Neurosci. 2004, 24, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Brain, K.L.; Wilson, R.H.; Grem, L.L.; Vincent, A. Oxaliplatin induces hyperexcitability at motor and autonomic neuromuscular junctions through effects on voltage-gated sodium channels. Br. J. Pharmacol. 2005, 146, 1027–1039. [Google Scholar] [CrossRef] [PubMed]
- Khaliq, Z.M.; Raman, I.M. Relative contributions of axonal and somatic Na channels to action potential initiation in cerebellar Purkinje neurons. J. Neurosci. 2006, 26, 1935–1944. [Google Scholar] [CrossRef] [PubMed]
- Yokote, S.; Setoguchi, R.; Shimizu, E.; Mishima, N.; Kawahara, K.; Kuniyasu, A.; Shirasaki, T.; Takahama, K.; Konno, K.; Kawai, N.; et al. A synthetic approach to develop peptide inhibitors selective for brain-type sodium channels on the basis of pompilidotoxin structure. Heterocycles 2009, 79, 925–933. [Google Scholar]
- Magloire, V.; Czrnecki, A.; Anwander, H.; Streit, J. β-Pompilidotoxin modulated spontaneous activity and persistent sodium currents in spinal networks. Neuroscience 2011, 172, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, N.; Zhu, H.-L.; Yotsu-Yamashita, M.; Inai, T.; Cunnane, T.C. Resurgent-like currents in mouse vas deferens myocytes are mediated by NaV1.6 voltage-gated sodium channels. Pflugers Arch. 2012, 464, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Kawai, N.; Miwa, A.; Yasuhara, T.; Morimoto, Y.; Nakata, Y. Structure and biological activities of eumenine mastoparan-AF (EMP-AF), a novel mast cell degranulating peptide in the venom of the solitary wasp (Anterhynchium flavomarginatum micado). Toxicon 2000, 38, 1505–1515. [Google Scholar] [CrossRef]
- Cabrera, M.P.S.; Souza, B.M.; Fontana, R.; Konno, K.; Palma, M.S.; de Azevedo, W.F., Jr.; Ruggiero Neto, J. Conformation and lytic activity of eumenine mastoparan: A new antimicrobial peptide from wasp venom. J. Peptide Res. 2004, 64, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Sforça, M.L.; Oyama, S., Jr.; Canduri, F.; Lorenzi, C.C.B.; Pertinez, T.A.; Konno, K.; Souza, B.M.; Palma, M.S.; Ruggiero Neto, J.; de Azevedo, W.F., Jr.; et al. How C-terminal carboxyamidation alters the mast cell degranulating activity of peptides from the venom of the eumenine solitary wasp. Biochemistry 2004, 43, 5608–5617. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Shinada, T.; Ohfune, Y.; Hisada, M.; Yasuda, A.; Naoki, H.; Nakajima, T. Novel mastoparan and protonectin analogs isolated from a solitary wasp, Orancistrocerus drewseni drewseni. Amino Acids 2009, 37, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Lee, S.H. Isolation and molecular cloning of venom peptides from Orancistrocerus drewseni (Hymenoptera: Eumenidae). Toxicon 2010, 55, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Fontana, R.; Rangel, M.; Oliveira, J.S.; Cabrera, M.P.; Ruggiero Neto, J.; Hide, I.; et al. Eumenitin, a novel antimicrobial peptide from the venom of the solitary eumenine wasp Eumenes rubronotatus. Peptides 2006, 27, 2624–2631. [Google Scholar] [CrossRef] [PubMed]
- Arcisio-Miranda, M.; Cabrera, M.P.S.; Konno, K.; Rangel, M.; Procopio, J. Effects of the cationic antimicrobial peptide Eumenitin from the venom of solitary wasp Eumenes rubronotatus in planar lipid bilayers: Surface charge and pore formation activity. Toxicon 2008, 51, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Rangel, M.; Cabrera, M.P.S.; Kazuma, K.; Ando, K.; Wang, X.; Kato, M.; Nihei, K.; Hirata, I.Y.; Cross, T.; Garcia, A.N.; et al. Chemical and biological characterization of four new antimicrobial and α-helical peptides from the venoms of two solitary eumenine wasps. Toxicon 2011, 57, 1081–1092. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Lee, S.H. Differential gene expression profiles in the venom gland/sac of Eumenes pomiformis (Hymenoptera: Eumenidae). Toxicon 2010, 55, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Rangel, M.; Oliveira, J.S.; Cabrera, M.P.S.; Fontana, R.; Hirata, I.Y.; Nakata, Y.; Mori, K.; Kawano, M.; Fuchino, H.; et al. Decoralin, a novel linear cationic α-helical peptide from the venom of the solitary eumenine wasp Oreumenes decoratus. Peptides 2007, 28, 2320–2327. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hisada, M.; Fontana, R.; Lorenzi, C.C.B.; Naoki, H.; Itagaki, Y.; Miwa, A.; Kawai, N.; Nakata, Y.; Yasuhara, T.; et al. Anoplin, a novel antimicrobial peptide from the venom of the solitary wasp Anoplius samariensis. Biochim. Biophys. Acta 2001, 1550, 70–80. [Google Scholar] [CrossRef]
- Cabrera, M.P.S.; Arcisio-Miranda, M.; Costa, S.T.B.; Konno, K.; Ruggiero, J.R.; Procopio, J.; Ruggiero Neto, J. Study of the mechanism of action of Anoplin, a helical antimicrobial decapeptide with ion channel-like activity, and the role of the amidated C-terminus. J. Peptide Sci. 2008, 14, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Munk, J.K.; Uggerhøj, L.E.; Poulsen, T.J.; Frimodt-Møller, N.; Wimmer, R.; Nyberg, N.T.; Hansen, P.R. Synthetic analogs of anoplin show improved antimicrobial activities. J. Pept. Sci. 2013, 19, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Jindrichova, B.; Burketova, L.; Novotna, Z. Novel properties of antimicrobial peptide anoplin. Biochem. Biophys. Res. Commun. 2014, 444, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, J.; Zheng, X.; Yang, X.; Ma, P.; Cai, Y.; Zhang, B.; Chen, Y. Design of novel analogues of short antimicrobial peptide anoplin with improved antimicrobial activity. J. Pept. Sci. 2014, 20, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Daben, M.; Libardo, J.; Nagella, S.; Lugo, A.; Pierce, S.; Angeles-Boza, A.M. Copper-binding tripeptide motif increases potency of the antimicrobial peptide Anoplin via Reactive Oxygen Species generation. Biochem. Biophys. Res. Commun. 2015, 456, 446–451. [Google Scholar]
- Kuhn-Nentwig, L. Antimicrobial and cytolytic peptides of venomous arthropods. CMLS Cell. Mol. Life Sci. 2003, 60, 2651–2668. [Google Scholar] [CrossRef] [PubMed]
- Moreau, S.J.M. “It stings a bit but it cleans well”: Venoms of Hymenoptera and their antimicrobial potential. J. Insect Physiol. 2013, 59, 186–204. [Google Scholar] [CrossRef] [PubMed]
- Piek, T. Neurotoxic kinins from wasp and ant venoms. Toxicon 1991, 29, 139–149. [Google Scholar] [CrossRef]
- Konno, K.; Palma, M.S.; Hirata, I.Y.; Juliano, M.A.; Juliano, L.; Yasuhara, T. Identification of bradykinins in solitary wasp venoms. Toxicon 2002, 40, 309–312. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Yasuhara, T.; Juliano, M.A.; Juliano, L.; Palma, M.S.; Nakajima, T. Isolation and sequence determination of peptides in the venom of the spider wasp Cyphononyx dorsalis guided by matrix-assisted laser desorption/ionization time of flight (MALDI-TOF) mass spectrometry. Toxicon 2001, 39, 1257–1260. [Google Scholar] [CrossRef]
- Picolo, G.; Hisada, M.; Sciani, J.M.; Conceição, I.M.; Machado, M.; Oliveira, V.; de Melo, R.L.; Cury, Y.; Konno, K.; Hayashi, M.A.F. Bradykinin-related peptides from the venom of the solitary wasp Cyphononyx fulvognathus. Biochem. Pharmacol. 2010, 79, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Arimoto, H.; Kinumi, T.; Oba, Y.; Uemura, D. Identification of proteins from venom of the paralytic spider wasp, Cyphononyx dorsalis. Insect Biochem. Mol. Biol. 2007, 37, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Lee, S.H. Identification and characterization of venom proteins of two solitary wasps, Eumenes pomiformis and Orancistrocerus drewseni. Toxicon 2010, 56, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Frølund, S.; Bella, A.; Kristensen, A.S.; Ziegler, H.L.; Witt, M.; Olsen, C.A.; Strømgaard, K.; Henrik Franzyk, H.; Jaroszewski, J.W. Assessment of structurally diverse philanthotoxin analogues for inhibitory activity on ionotropic Glutamate receptor subtypes: Discovery of nanomolar, nonselective, and use-dependent antagonists. J. Med. Chem. 2010, 53, 7441–7451. [Google Scholar] [CrossRef] [PubMed]
- Nihei, K.; Kazuma, K.; Ando, K.; Konno, K. Chemical and biological characterization of a novel neuropeptide in the venom of solitary digger wasp. Toxicon 2012, 60, 144. [Google Scholar] [CrossRef]
- Orchard, I.; Lange, A.B.; Bendena, W.G. FMRFamide-related peptides: A multifunctional family of structurally related neuropeptides in insects. Adv. Insect Physiol. 2001, 28, 267–329. [Google Scholar]
- Peeff, N.M.; Orchard, I.; Lange, A.B. Isolation, sequence, and bioactivity of PDVDHVFLRFamide and ADVGHVFLRFamide peptides from the locust central nervous system. Peptides 1994, 15, 387–392. [Google Scholar] [CrossRef]
- Holman, G.M.; Cook, B.J.; Nachman, R.J. Isolation, primary structure and synthesis of Leucomyosuppressin, an insect neuropeptide that inhibits spontaneous contractions of the cockroach hindgut. Comp. Biochem. Physiol. 1986, 85C, 329–333. [Google Scholar]
- Hisada, M.; Konno, K.; Itagaki, Y.; Naoki, H.; Nakajima, T. Sequencing wasp venom peptides by endopeptidase digestion and nested collision-induced dissociation/post-source decay method. Rapid Commun. Mass Spectr. 2002, 16, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Hisada, M.; Satake, H.; Masuda, K.; Aoyama, M.; Murata, K.; Shinada, T.; Iwashita, T.; Ohfune, Y.; Nakajima, T. Molecular components and toxicity of the venom of the solitary wasp, Anoplius samariensis. Biochem. Biophys. Res. Commun. 2005, 330, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Baek, J.H.; Yoon, K.A. Differential properties of venom peptides and proteins in solitary vs. social hunting wasps. Toxins 2016, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Hisada, M.; Konno, K.; Itagaki, Y.; Naoki, H.; Nakajima, T. Advantages of using nested collision induced dissociation/post-source decay with matrix-assisted laser desorption/ionization time of flight mass spectrometry: Sequencing of novel peptides from wasp venom. Rapid Commun. Mass Spectr. 2000, 14, 1828–1834. [Google Scholar] [CrossRef]
- Konno, K. Bioactive peptides in solitary wasp venoms. In Program & Abstracts, Proceedings of the 10th IST Asia Pacific Conference on Animal, Plants and Microbial Toxins, Changsha, China, June 2014; p. 18.
Figure 1.
The spider wasp Anoplius samariensis, taking the paralyzed prey to her nest.

Figure 2.
The spider paralyzed by the stinging venom of Cyphononyx fulvognathus.

Figure 3.
The digger wasp Sphex argentatus argentatus is stinging the grasshopper prey.


{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Name | Sequence |
|---|---|---|---|
| Mastoparan | INLKALAALAKKIL-NH2 | EMP-OD | GRILSFIKGLAEHL-NH2 |
| EMP-AF | INLLKIAKGIIKSL-NH2 | Eumenitin | LNLKGIFKKVKSLLT |
| EMP-ER | FDIMGLIKKVAGAL-NH2 | Eumenitin-R | LNLKGLIKKVASLLN |
| EMP-EF | FDVMGIIKKIASAL-NH2 | Eumenitin-F | LNLKGLFKKVASLLT |
| EpVP2a | FDLLGLVKKVASAL-NH2 | EpVP1 | INLKGLIKKVASLLT |
| Anoplin | GLLKRIKTLL-NH2 | Decoralin | SLLSLIRKLIT |
| Name | Sequence |
|---|---|
| Cd-125 | DTARLKWH |
| As-126 | pQDPPVVKMK-NH2 |
| Bm-10 | pQTAPVPKAISK-NH2 |
| Orancis-Protonectin | ILGIITSLLKSL-NH2 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Konno, K.; Kazuma, K.; Nihei, K.-i. Peptide Toxins in Solitary Wasp Venoms. Toxins 2016, 8, 114. https://doi.org/10.3390/toxins8040114
AMA Style
Konno K, Kazuma K, Nihei K-i. Peptide Toxins in Solitary Wasp Venoms. Toxins. 2016; 8(4):114. https://doi.org/10.3390/toxins8040114
Chicago/Turabian StyleKonno, Katsuhiro, Kohei Kazuma, and Ken-ichi Nihei. 2016. "Peptide Toxins in Solitary Wasp Venoms" Toxins 8, no. 4: 114. https://doi.org/10.3390/toxins8040114
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.