
Isolation of the Binding Protein of Periplocoside E from BBMVs in Midgut of the Oriental Amyworm Mythimna separata Walker (Lepidoptera: Noctuidae) through Affinity Chromatography

Abstract
:
1. Introduction
2. Results and Discussion
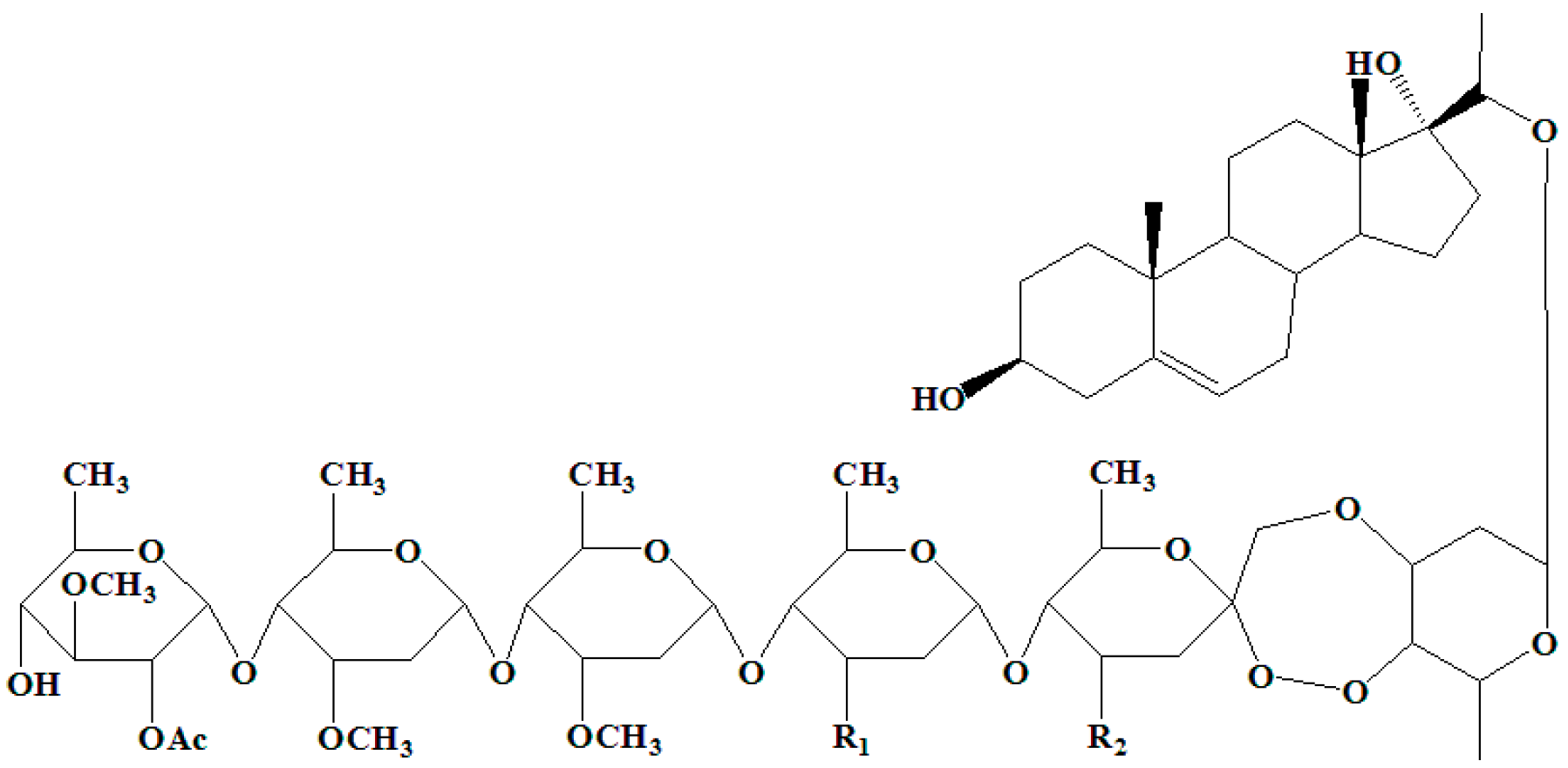
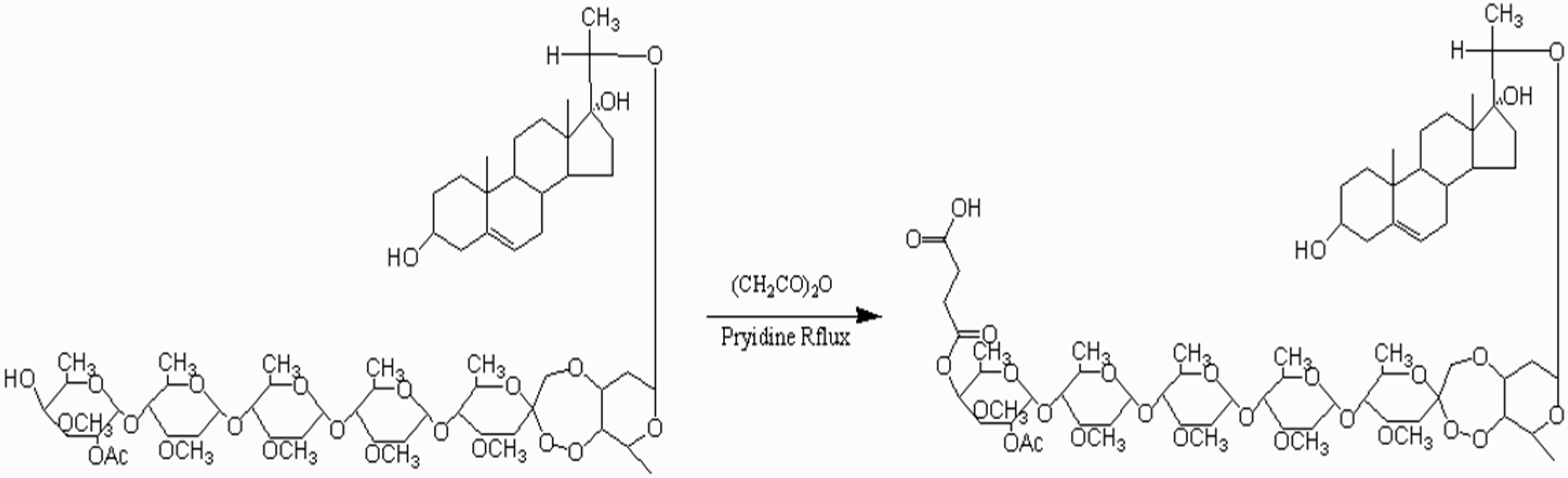
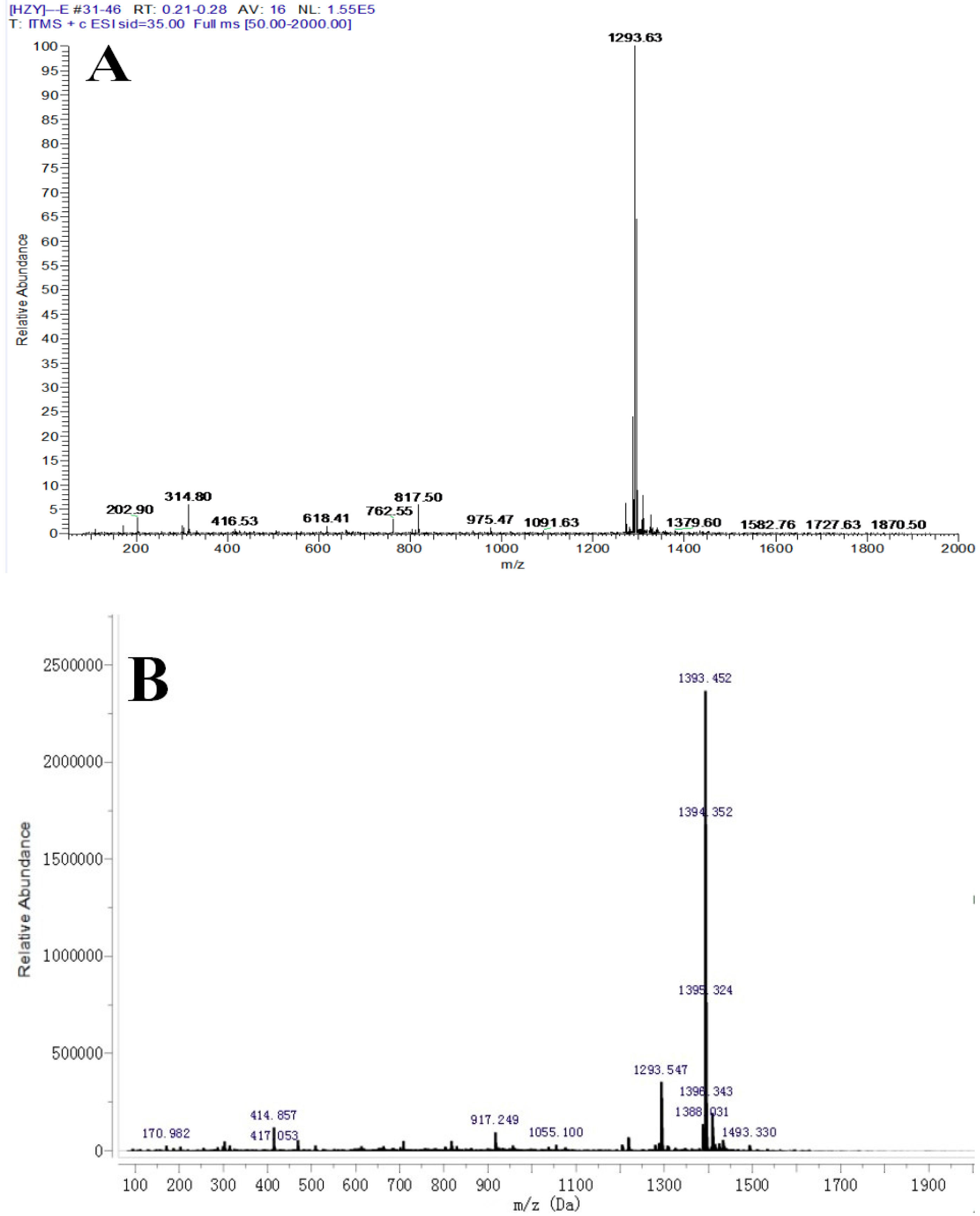
2.1. Identification of the Ligand (Periplocoside E-Semi-Succinic Acid Ester)
2.2. Isolation of Binding Proteins
3. Conclusions
4. Materials and Methods
4.1. Insects
4.2. Chemicals
4.3. Preparation of Brush Border Membrane Vesicles (BBMV)
4.4. Synthesis of Ligand
4.5. Preparation of the Medium and Coupling of the Ligand
4.6. Affinity Chromatography
4.7. SDS-PAGE and LC/Q-TOF-MS Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Senthil-Nathan, S. A review of biopesticides and their mode of action against insect pests. In Environmental Sustainability; Springer: New Delhi, India, 2015; pp. 49–63. [Google Scholar]
- Mouhssen, L. Screening of natural products for drug discovery. Expert Opin. Drug Discov. 2007, 2, 697–705. [Google Scholar]
- Zhang, K.; Pugliese, M.; Pugliese, A.; Passantino, A. Biological active ingredients of traditional Chinese herb astragalus membranaceus on treatment of diabetes: A systematic review. Mini Rev. Med. Chem. 2015, 15, 315–329. [Google Scholar] [PubMed]
- Chen, B.; Butte, A.J. Leveraging big data to transform target selection and drug discovery. Clin. Pharmacol. Ther. 2016, 99, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Li, T.X.; Zhang, C.Y.; Li, G.H.; Li, Q.H.; Zhang, Y.J. Ecological and biological characteristics of Periploca sepium. Drugs Clin. 2013, 28, 730–733. [Google Scholar]
- Tokiwa, T.; Harada, K.; Matsumura, T.; Tukiyama, T. Oriental medicinal herb, Periploca sepium, extract inhibits growth and IL-6 production of human synovial fibroblast-like cells. Biol. Pharm. Bull. 2004, 27, 1691–1693. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.W.; Yang, H.; Li, J.H.; Qian, Y.; Wu, W.J. Studies on insecticidal constituents from Periploca sepium Bunge. Acta Boreali-Occident. Sin. 2006, 15, 620–623. [Google Scholar]
- Li, Y.; Zeng, X.N.; Wang, W.Z.; Luo, C.H.; Yan, Q.; Tian, M. Chemical constituents from the roots of Periploca sepium with insecticidal activity. J. Asian Nat. Prod. Res. 2012, 14, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.X.; Zhao, J.; Zhang, J.W.; Hu, Z.N.; Wen, W.J. Fluorescence localization and comparative ultrastructural study of periplocoside NW from Periploca sepium Bunge in the midgut of the Oriental Amyworm, Mythimna separata Walker (Lepidoptera: Noctuidae). Toxins 2014, 6, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.N.; Sun, W.C.; Feng, M.X.; Hu, Z.N.; Wen, W.J. Immuno-electron microscopic localization research of periplocoside T from Periploca sepium Bunge on midgut of larvae in the Oriental Amyworm, Mythimna separata Walker (Lepidoptera: Noctuidae). J. Northwest A F Univ. 2015, 44, 118–122. [Google Scholar]
- Feng, M.X.; Shi, B.J.; Zhao, Y.C.; Hu, Z.N.; Wen, W.J. Histopathological effects and immunolocalization of periplocoside NW from Periploca sepium Bunge on the midgut epithelium of Mythimna separata Walker larvae. Pestic. Biochem. Physiol. 2014, 115, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Kool, J.; Jonker, N.; Irth, H.; Niessen, W.M.A. Studying protein-protein affinity and immobilized ligand-protein affinity interactions using MS-based methods. Anal. Bioanal. Chem. 2011, 401, 1109–1125. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.T. Separation and Preparation of Polyclonal Antibody of Perplocoside T; Northwest A & F University: Yangling, China, 2012. [Google Scholar]
- Dirk, D.; Fereshte, A.S. Botulinum toxin: Mechanisms of action. Eur. Neurol. 2005, 63, 3–9. [Google Scholar]
- Yang, R.Y.; Liu, H.X.; Hu, Z.N.; Zhang, L.S.; Tian, X.M. Study on the mechanism of action of celangulin V. J. Northwest Agric. Univ. 2000, 28, 18–21. [Google Scholar]
- Knight, P.J.K.; Crickmore, N.; Ellar, D.J. The receptor for Bacillus thuringiensis CrylA(c) delta-endotoxin in the brush border membrane of the lepidopteran Manduca sexta is aminopeptidase N. Mol. Microbiol. 1994, 11, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Francis, B.R.; Bulla, L.A. Further characterization of BT-R1, the cadherin-like receptor for Cry1Ab toxin in tobacco hornworm (Manduca sexta) Midguts. Insect Biochem. Mol. Biol. 1997, 27, 541–550. [Google Scholar] [CrossRef]
- Chen, W.B.; Liu, C.X.; Xiao, Y.T.; Zhang, D.D.; Zhang, Y.D.; Li, X.C.; Tabashnik, B.E.; Wu, K.M. A toxin-binding alkaline phosphatase fragment synergizes Bt toxin Cry1Ac against susceptible and resistant Helicoverpa armigera. PLoS ONE 2015, 10, e0126288. [Google Scholar] [CrossRef] [PubMed]
- Vega-Cabrera, A.; Cancino-Rodezno, A.; Porta, H.; Pardo-Lopez, L. Aedes aegypti Mos20 cells internalizes cry toxins by endocytosis, and actin has a role in the defense against Cry11Aa toxin. Toxins 2014, 6, 464–487. [Google Scholar] [CrossRef]
- Lu, L.N.; Qi, Z.J.; Zhang, J.W.; Wu, W.J. Separation of binding protein of Celangulin V from the midgut of Mythimna separata Walker by affinity chromatography. Toxins 2015, 7, 1738–1748. [Google Scholar] [CrossRef] [PubMed]
- Mounier, N.; Sparrw, J.C. Muscle actin genes in insects. Comp. Biochem. Physiol. Part B 1993, 105, 231–238. [Google Scholar] [CrossRef]
- Doherty, G.J.; McMahon, H.T. Mediation, modulation, and consequences of membrane-cytoskeleton interactions. Annu. Rev. Biophys. 2008, 37, 65–95. [Google Scholar] [CrossRef] [PubMed]
- Pollard, T.D.; Cooper, J.A. Actin, a central player in cell shape and movement. Science 2009, 326, 1208–1212. [Google Scholar] [CrossRef] [PubMed]
- Rescher, U.; Ruhe, D.; Ludwig, C.; Zobiack, N.; Gerke, V. Annexin 2 is a phosphatidylinositol (4,5)-bisphosphate binding protein recruited to actin assembly sites at cellular membranes. J. Cell Sci. 2004, 117, 3473–3480. [Google Scholar] [CrossRef] [PubMed]
- Hafkenscheid, J.C.M. Aminopeptidases and amino acid arylamidases. Methods Enzym. Anal. 1984, 5, 2–34. [Google Scholar]
- Wolfersberger, M.G. Enzymology of plasma membranes of insect intestinal cells. Am. Zool. 1984, 24, 187–197. [Google Scholar] [CrossRef]
- Taylor, A. Aminopeptidases: Structure and function. FASEB J. 1993, 7, 290–298. [Google Scholar] [PubMed]
- Knight, P.J.K.; Carroll, J.; Ellar, D.J. Analysis of glycan structures on the 120 kDa aminopeptidase N of Manduca sexta and their interactions with Bacillus thuringiensis Cry1Ac toxin. Insect Biochem. Mol. Biol. 2004, 34, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Cowles, E.A.; Francis, V. Identification, isolation, and cloning of a Bacillus thuringiensis CryIAc toxin-binding protein from the midgut of the lepidoptera insect Heliothis virescens. J. Biol. Chem. 1995, 270, 27277–27282. [Google Scholar] [CrossRef] [PubMed]
- Valaitis, A.P.; Mi, K.L.; Rajamohan, F.; Dean, D.H. Brush border membrane aminopeptidase-N in the midgut of the gypsy moth serves as the receptor for the CryIA (c) δ-endotoxin of Bacillus thuringiensis. Insect Biochem. Mol. Biol. 1995, 25, 1143–1151. [Google Scholar] [CrossRef]
- Lee, M.K.; You, T.H.; Young, B.A.; Cotrill, J.A.; Valaitis, A.P.; Dean, D.H. Aminopeptidase N purified from gypsy moth brush border membrane vesicles is a specific receptor for Bacillus thuringiensis CryIAc toxin. Appl. Environ. Microbiol. 1996, 62, 2845–2849. [Google Scholar] [PubMed]
- Gill, M.; Ellar, D. Transgenic Drosophila reveals a functional in vivo receptor for the Bacillus thuringiensis toxin Cry1Ac1. Insect Mol. Biol. 2002, 11, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G. Anchoring and biosynthesis of stalked brush border membrane proteins: Glycosidases and peptidases of enterocytes and renal tubuli. Annu. Rev. Cell Biol. 1986, 2, 255–307. [Google Scholar] [CrossRef] [PubMed]
- Gierasch, L.M. Signal sequences. Biochemistry 1989, 28, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Delmas, B.; Gelfi, J.; L’Haridon, R.; Vogel, L.K.; Sjöström, H.; Norén, O.; Laude, H. Aminopeptidase N is a major receptor for the entero-pathogenic coronavirus TGEV. Nature 1992, 357, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Yeager, C.L.; Ashmun, R.A.; Williams, R.K.; Cardellichio, C.B.; Shapiro, L.H.; Look, A.T.; Holmes, K.V. Human aminopeptidase N is a receptor for human coronavirus 229E. Nature 1992, 357, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.J. Studies on the Insecticidal Constituent of the Root Bark of Periploca sepium and Tripterygium hypoglaucum; Northwest A & F University: Yangling, China, 2009. [Google Scholar]
- Wolfersberger, M.; Luethy, P.; Maurer, A.; Parenti, P.; Sacchi, F.V.; Giordana, B.; Hanozet, G.M. Preparation and partial characterization of amino acid transporting brush border membrane vesicles from the larval midgut of the cabbage butterfly (Pieris brassicae). Comp. Biochem. Physiol. Part A 1987, 86, 301–308. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Protein ID | Protein Mass | Protein |
|---|---|---|---|
| 1 | gi|512918093 | 63095 | Luciferin 4-monooxygenase |
| 2 | gi|15212557 | 114719 | Aminopeptidase N |
| 3 | gi|21327773 | 107153 | Aminopeptidase N3 |
| 4 | gi|253807662 | 66764 | Nicotinamide adenine dinucleotide health (NADH) dehydrogenase subunit 5 |
| 5 | gi|520836887 | 78123 | Phosphatidylinositol 3-phosphate 3-phosphatase myotubularin |
| 6 | gi|323435326 | 31100 | Actin |
| 7 | gi|512901127 | 73062 | Uncharacterized family 31 glucosidase KIAA1161 |
| 8 | gi|399942986 | 86025 | 2OG-Fe(2) oxygenase superfamily protein |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, M.; He, Z.; Wang, Y.; Yan, X.; Zhang, J.; Hu, Z.; Wu, W. Isolation of the Binding Protein of Periplocoside E from BBMVs in Midgut of the Oriental Amyworm Mythimna separata Walker (Lepidoptera: Noctuidae) through Affinity Chromatography. Toxins 2016, 8, 139. https://doi.org/10.3390/toxins8050139
Feng M, He Z, Wang Y, Yan X, Zhang J, Hu Z, Wu W. Isolation of the Binding Protein of Periplocoside E from BBMVs in Midgut of the Oriental Amyworm Mythimna separata Walker (Lepidoptera: Noctuidae) through Affinity Chromatography. Toxins. 2016; 8(5):139. https://doi.org/10.3390/toxins8050139
Chicago/Turabian StyleFeng, Mingxing, Zhenyu He, Yuanyuan Wang, Xiufang Yan, Jiwen Zhang, Zhaonong Hu, and Wenjun Wu. 2016. "Isolation of the Binding Protein of Periplocoside E from BBMVs in Midgut of the Oriental Amyworm Mythimna separata Walker (Lepidoptera: Noctuidae) through Affinity Chromatography" Toxins 8, no. 5: 139. https://doi.org/10.3390/toxins8050139