
Is qPCR a Reliable Indicator of Cyanotoxin Risk in Freshwater?


Abstract
:1. Introduction
1.1. Traditional Methods for Cyanotoxin Detection
1.2. Molecular Tests for Detecting Toxic Cyanobacteria
2. Identification of Gene Clusters Encoding the Enzymes Responsible for Cyanotoxin Synthesis
2.1. Microcystin
2.2. Cylindrospermopsin
2.3. Saxitoxin
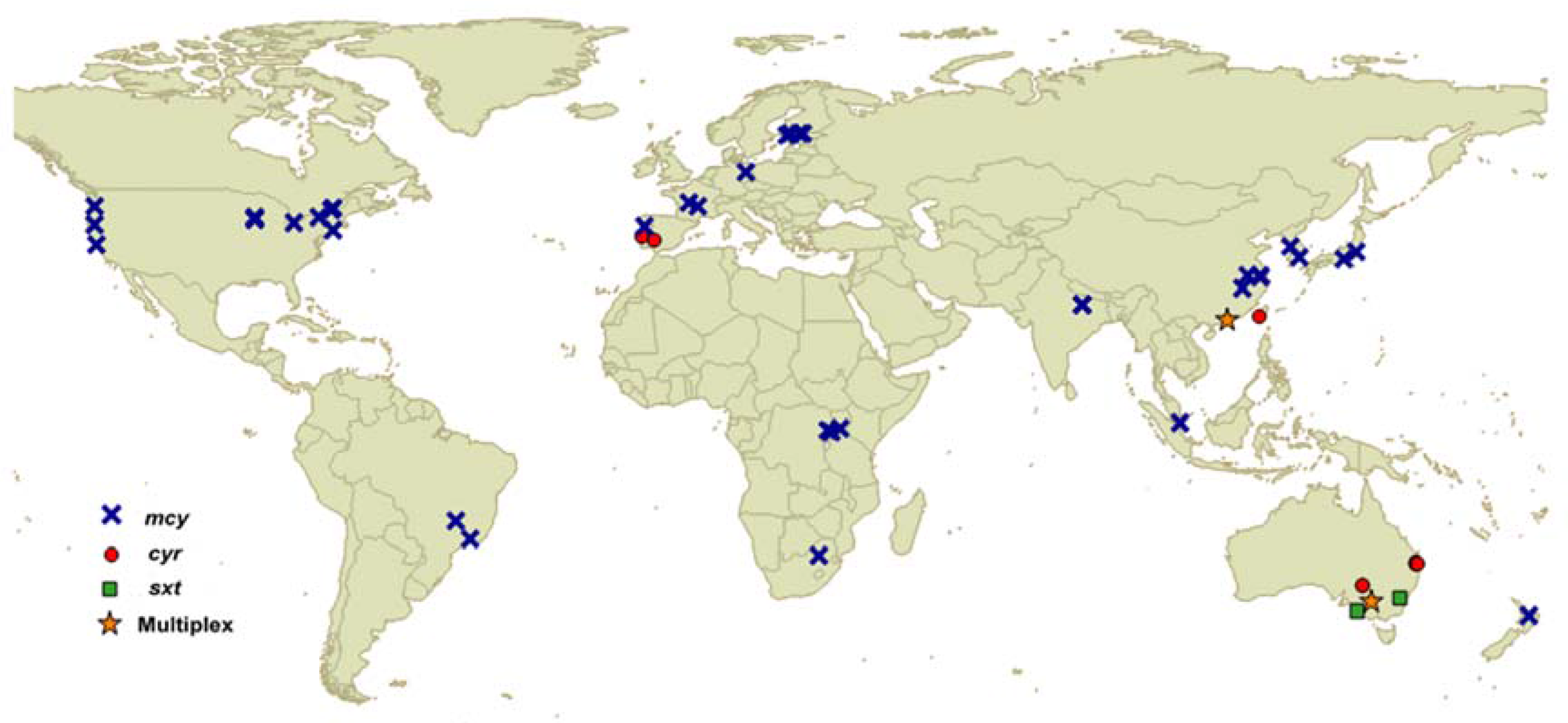
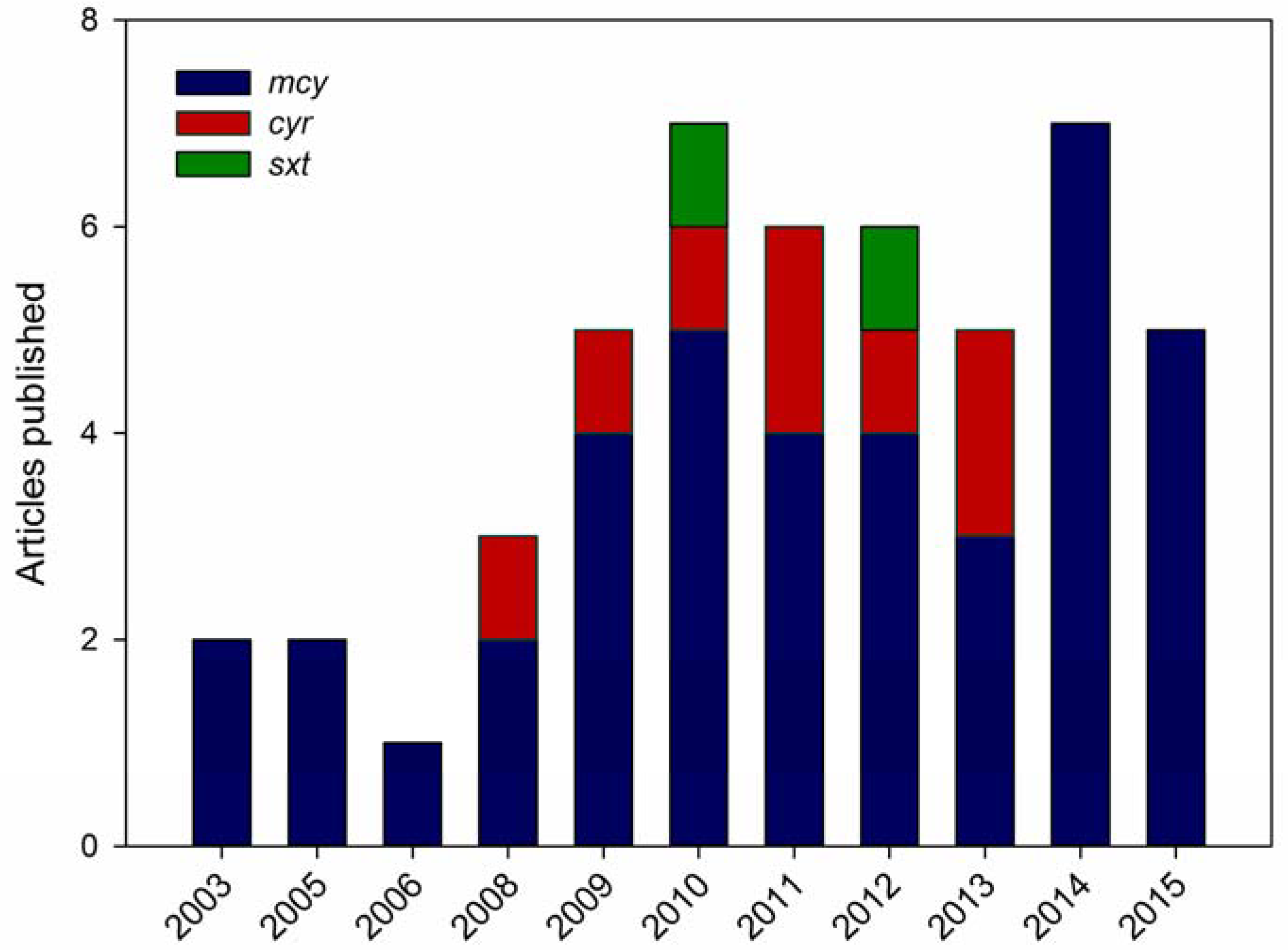
3. qPCR for Cyanotoxins
3.1. qPCR for Detecting Microcystin Genes
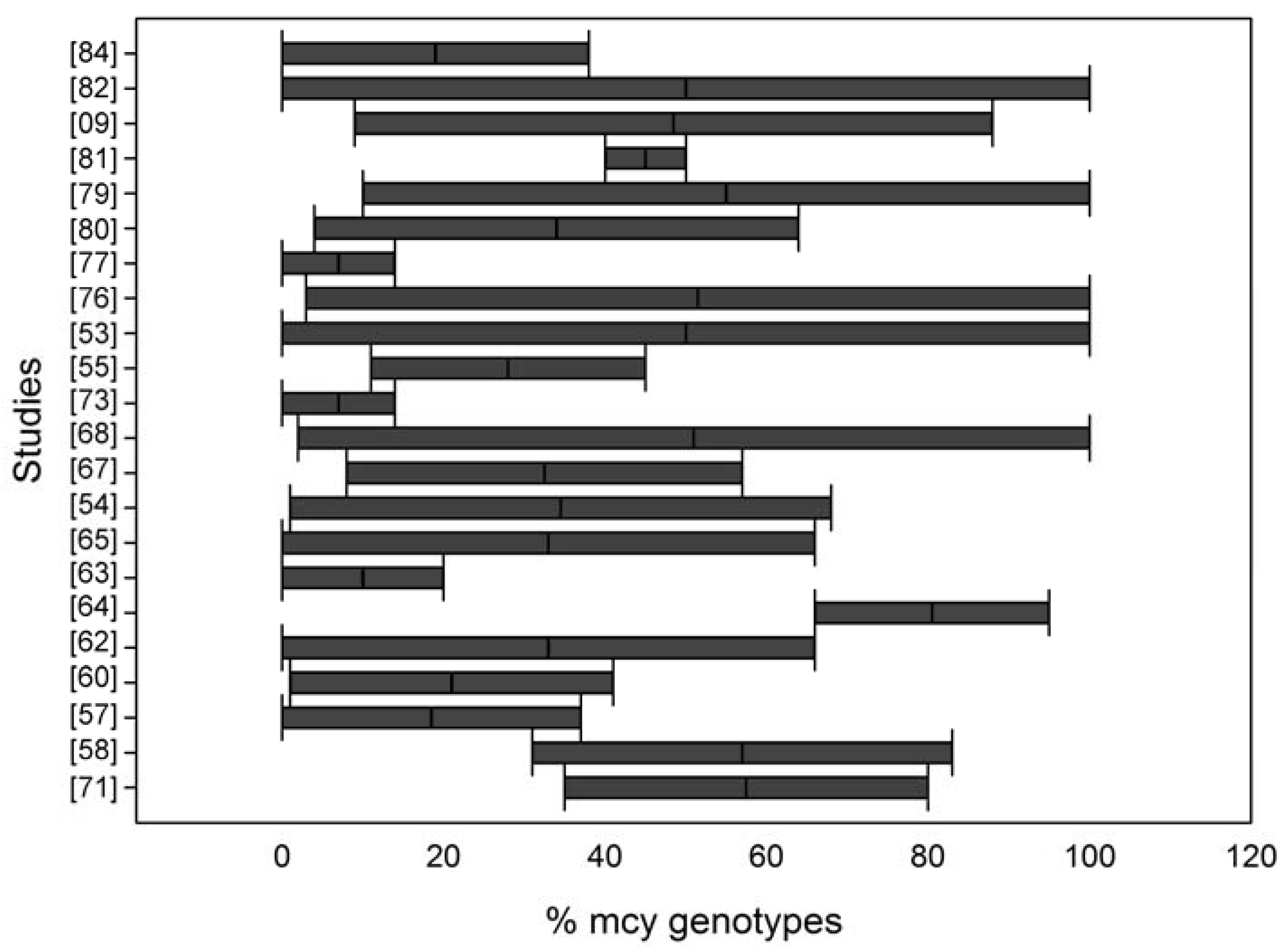
qPCR for Determining the Microcystin Toxic Genotype Proportion
3.2. qPCR for Detecting Cylindrospermopsin Genes
3.3. qPCR for Detecting Saxitoxin Genes
3.4. Multiplex qPCR for Cyanotoxins
3.5. Advantages of qPCR for Estimating Cyanotoxin Concentrations in the Environment
3.6. Limitations of qPCR for Estimating Cyanotoxin Concentrations in the Environment
4. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A guide to their public health consequences, monitoring and management, 1st ed.; Chorus, I., Bartram, J., Eds.; E & FN Spon: London, UK, 1999. [Google Scholar]
- Meissner, S.; Fastner, J.; Dittmann, E. Microcystin production revisited: Conjugate formation makes a major contribution. Environ. Microbiol. 2013, 15, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, R.; Balasubramanian, R. Methods and approaches used for detection of cyanotoxins in environmental samples: A Review. Crit. Rev. Environ. Sci. Technol. 2013, 3389, 1349–1383. [Google Scholar] [CrossRef]
- Srivastava, A.; Singh, S.; Ahn, C.; Oh, H.; Asthana, R.K. Monitoring approaches for a toxic cyanobacterial bloom. Environ. Sci. Technol. 2013, 47, 8999–9013. [Google Scholar] [CrossRef] [PubMed]
- Testai, E.; Buratti, F.M.; Funari, E.; Manganelli, M.; Vichi, S.; Arnich, N.; Biré, R.; Fessard, V.; Sialehaamoa, A. Review and analysis of occurrence, exposure and toxicity of cyanobacteria toxins in food. EFSA Support. Publ. 2016, 998, 309. Available online: http://www.efsa.europa.eu/sites/default/files/scientific_output/files/main_documents/998e.pdf (accessed on 2 June 2016). [Google Scholar]
- Graham, J.L.; Loftin, K.A.; Meyer, M.T.; Ziegler, A.C. Co-occurrence of toxins and taste-and-odor compounds in cyanobacterial blooms from the midwestern United States. Environ. Sci. Technol. 2006, 44, 7361–7368. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, J.; Yang, M.; Zhang, J.; Burch, M.D.; Han, W. Cyanobacterial population and harmful metabolites dynamics during a bloom in Yanghe Reservoir, North China. Harmful Algae 2010, 9, 481–488. [Google Scholar] [CrossRef]
- Al-Tebrineh, J.; Merrick, C.; Ryan, D.; Humpage, A.; Bowling, L.; Neilan, B.A. Community composition, toxigenicity, and environmental conditions during a cyanobacterial bloom occurring along 1,100 kilometers of the Murray River. Appl. Environ. Microbiol. 2012, 78, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Kong, F.; Zhang, M.; Yang, Z.; Shi, X.; Du, M. The dynamics of microcystis genotypes and microcystin production and associations with environmental factors during blooms in lake Chaohu, China. Toxins (Basel). 2014, 6, 3238–3257. [Google Scholar] [CrossRef] [PubMed]
- Otten, T.G.; Crosswell, J.R.; Mackey, S.; Dreher, T.W. Application of molecular tools for microbial source tracking and public health risk assessment of a Microcystis bloom traversing 300km of the Klamath River. Harmful Algae 2015, 46, 71–81. [Google Scholar] [CrossRef]
- Sabart, M.; Crenn, K.; Perrière, F.; Abila, A.; Leremboure, M.; Colombet, J.; Jousse, C.; Latour, D. Co-occurrence of microcystin and anatoxin-a in the freshwater lake Aydat (France): Analytical and molecular approaches during a three-year survey. Harmful Algae 2015, 48, 12–20. [Google Scholar] [CrossRef]
- Neilan, B.A.; Jacobs, D.; Del Dot, T.; Blackall, L.L.; Hawkins, P.R.; Cox, P.T.; Goodman, A.E. rRNA sequences and evolutionary relationships among toxic and nontoxic cyanobacteria of the genus Microcystis. Int. J. Syst. Bacteriol. 1997, 47, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Tillett, D.; Parker, D.L.; Neilan, B.A. Detection of toxigenicity by a probe for the microcystin synthetase a gene (mcyA) of the cyanobacterial genus Microcystis: Comparison of toxicities with 16S rRNA and phycocyanin operon (phycocyanin intergenic spacer) phylogenies. Appl. Environ. Microbiol. 2001, 67, 2810–2818. [Google Scholar] [CrossRef] [PubMed]
- Neilan, B.A. The molecular evolution and DNA profiling of toxic cyanobacteria. Curr. Issues Mol. Biol 2002, 4, 1–11. [Google Scholar] [PubMed]
- Pearson, L.A.; Neilan, B.A. The molecular genetics of cyanobacterial toxicity as a basis for monitoring water quality and public health risk. Curr. Opin. Biotechnol. 2008, 19, 281–8. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, T.; Ueda, A.; Fujii, K.; Harada, K.-I.; Ochi, K.; Shirai, M. Polyketide involved synthase gene in the biosynthesis coupled of the to the cyclic peptide synthetase module heptapeptide microcystin. J. Biochem. 2000, 127, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Tillett, D.; Dittmann, E.; Erhard, M.; von Döhren, H.; Börner, T.; Neilan, B. a Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef]
- Christiansen, G.; Fastner, J.; Erhard, M.; Bo, T.; Dittmann, E. Microcystin biosynthesis in Planktothrix: Genes, evolution, and manipulation. Microbiology 2003, 185, 564–572. [Google Scholar] [CrossRef]
- Rouhiainen, L.; Vakkilainen, T.; Siemer, B.L.; Buikema, W.; Haselkorn, R.; Sivonen, K. Genes coding for hepatotoxic heptapeptides (Microcystins) in the cyanobacterium Anabaena Strain 90. Appl. Environ. Microbiol. 2004, 70, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Izaguirre, G.; Jungblut, A.D.; Neilan, B.A. Benthic cyanobacteria (Oscillatoriaceae) that produce microcystin-LR, isolated from four reservoirs in southern California. Water Res. 2007, 41, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.F.; Genuário, D.B.; da Silva, C.S.P.; Shishido, T.K.; Moraes, L.A.B.; Neto, R.C.; Silva-Stenico, M.E. Microcystin production by a freshwater spring cyanobacterium of the genus Fischerella. Toxicon 2009, 53, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Rantala, A.; Fewer, D.P.; Hisbergues, M.; Rouhiainen, L.; Vaitomaa, J.; Börner, T.; Sivonen, K. Phylogenetic evidence for the early evolution of microcystin synthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R.; Christiansen, G.; Fastner, J.; Börner, T. Abundance of active and inactive microcystin genotypes in populations of the toxic cyanobacterium Planktothrix spp. Environ. Microbiol. 2004, 6, 831–41. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, G.; Molitor, C.; Philmus, B.; Kurmayer, R. Nontoxic strains of cyanobacteria are the result of major gene deletion events induced by a transposable element. Mol. Biol. Evol. 2008, 25, 1695–1704. [Google Scholar] [CrossRef] [PubMed]
- Fewer, D.P.; Halinen, K.; Sipari, H.; Bernardová, K.; Mänttäri, M.; Eronen, E.; Sivonen, K. Non-autonomous transposable elements associated with inactivation of microcystin gene clusters in strains of the genus Anabaena isolated from the Baltic Sea. Environ. Microbiol. Rep. 2011, 3, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, Y.; Kaya, K.; Watanabe, M.M. Evidence for recombination in the microcystin synthetase (mcy) genes of toxic cyanobacteria Microcystis spp. J. Mol. Evol. 2004, 58, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Fewer, D.P.; Rouhiainen, L.; Jokela, J.; Wahlsten, M.; Laakso, K.; Wang, H.; Sivonen, K. Recurrent adenylation domain replacement in the microcystin synthetase gene cluster. BMC Evol. Biol. 2007, 7. [Google Scholar] [CrossRef] [PubMed]
- Tooming-Klunderud, A.; Fewer, D.P.; Rohrlack, T.; Jokela, J.; Rouhiainen, L.; Sivonen, K.; Kristensen, T.; Jakobsen, K.S. Evidence for positive selection acting on microcystin synthetase adenylation domains in three cyanobacterial genera. BMC Evol. Biol. 2008, 8. [Google Scholar] [CrossRef] [PubMed]
- Schembri, M.A.; Neilan, B.A.; Saint, C.P. Identification of genes implicated in toxin production in the cyanobacterium Cylindrospermopsis raciborskii. Environ. Toxicol. 2001, 16, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Shalev-Alon, G.; Sukenik, A.; Livnah, O.; Schwarz, R.; Kaplan, A. A novel gene encoding amidinotransferase in the cylindrospermopsin producing cyanobacterium Aphanizomenon ovalisporum. FEMS Microbiol. Lett. 2002, 209, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl. Environ. Microbiol. 2008, 74, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Moreira, C.; Vasconcelos, V.; Antunes, A. Phylogeny and biogeography of cyanobacteria and their produced toxins. Mar. Drugs 2013, 11, 4350–4369. [Google Scholar] [CrossRef] [PubMed]
- McGregor, G.B.; Sendall, B.C.; Hunt, L.T.; Eaglesham, G.K. Report of the cyanotoxins cylindrospermopsin and deoxy-cylindrospermopsin from Raphidiopsis mediterranea Skuja (Cyanobacteria/Nostocales). Harmful Algae 2011, 10, 402–410. [Google Scholar] [CrossRef]
- Stuken, A.; Jakobsen, K.S. The cylindrospermopsin gene cluster of Aphanizomenon sp. strain 10E6: Organization and recombination. Microbiology 2010, 156, 2438–2451. [Google Scholar] [CrossRef] [PubMed]
- Mazmouz, R.; Chapuis-Hugon, F.; Mann, S.; Pichon, V.; Mejean, A.; Ploux, O. Biosynthesis of cylindrospermopsin and 7-epicylindrospermopsin in Oscillatoria sp. strain PCC 6506: Identification of the cyr gene cluster and toxin analysis. Appl. Environ. Microbiol. 2010, 76, 4943–4949. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Xiao, P.; Yu, G.; Sano, T.; Pan, Q.; Li, R. Molecular basis and phylogenetic implications of deoxycylindrospermopsin biosynthesis in the cyanobacterium Raphidiopsis curvata. Appl. Environ. Microbiol. 2012, 78, 2256–2263. [Google Scholar] [CrossRef] [PubMed]
- Kellmann, R.; Mihali, T.K.; Jeon, Y.J.; Pickford, R.; Pomati, F.; Neilan, B.A. Biosynthetic intermediate analysis and functional homology reveal a saxitoxin gene cluster in cyanobacteria. 2008, 74, 4044–4053. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.A.; Mihali, T.K.; Neilan, B.A. Extraordinary conservation, gene loss, and positive selection in the evolution of an ancient neurotoxin. Mol. Biol. Evol. 2011, 28, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.M.J.; Wood, S.A.; van Ginkel, R.; Broady, P.A.; Gaw, S. First report of saxitoxin production by a species of the freshwater benthic cyanobacterium, Scytonema Agardh. Toxicon 2011, 57, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Borges, H.L.F.; Branco, L.H.Z.; Martins, M.D.; Lima, C.S.; Barbosa, P.T.; Lira, G.A.S.T.; Bittencourt-Oliveira, M.C.; Molica, R.J.R. Cyanotoxin production and phylogeny of benthic cyanobacterial strains isolated from the northeast of Brazil. Harmful Algae 2015, 43, 46–57. [Google Scholar] [CrossRef]
- Mihali, T.K.; Kellmann, R.; Neilan, B.A. Characterisation of the paralytic shellfish toxin biosynthesis gene clusters in Anabaena circinalis AWQC131C and Aphanizomenon sp. NH-5. BMC Biochem. 2009, 10. [Google Scholar] [CrossRef] [PubMed]
- Stucken, K.; John, U.; Cembella, A.; Murillo, A.A.; Soto-liebe, K.; Juan, J. The smallest known genomes of multicellular and toxic cyanobacteria: Comparison, minimal gene sets for linked traits and the evolutionary implications. PLoS ONE 2010, 5, e9235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihali, T.K.; Carmichael, W.W.; Neilan, B.A. A putative gene cluster from a Lyngbya wollei bloom that encodes paralytic shellfish toxin biosynthesis. PLoS ONE 2011, 6, e14657. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A.; Loram, J.E.; Hackett, J.D.; Anderson, D.M.; Plumley, F.G.; Bhattacharya, D. Origin of saxitoxin biosynthetic genes in cyanobacteria. PLoS ONE 2009, 4, e5758. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.; Vasconcelos, V. Use of qPCR for the study of hepatotoxic cyanobacteria population dynamics. Arch. Microbiol. 2011, 193, 615–27. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lim, J.; Lee, C. Quantitative real-time PCR approaches for microbial community studies in wastewater treatment systems: Applications and considerations. Biotechnol. Adv. 2013, 31, 1358–1373. [Google Scholar] [CrossRef] [PubMed]
- Foulds, I.V.; Granacki, A.; Xiao, C.; Krull, U.J.; Castle, A.; Horgen, P.A. Quantification of microcystin-producing cyanobacteria and E. coli in water by 5’ -nuclease PCR. J. Appl. Microbiol. 2002, 93, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R.; Christiansen, G.; Chorus, I. The abundance of microcystin-producing genotypes correlates positively with colony size in Microcystis sp. and determines its microcystin net production in Lake Wannsee. Appl. Environ. Microbiol. 2003, 69, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R.; Kutzenberger, T. Application of real-time PCR for quantification of microcystin genotypes in a population of the toxic cyanobacterium Microcystis sp.pdf. Appl. Environ. Microbiol. 2003, 69, 6723–6730. [Google Scholar] [CrossRef] [PubMed]
- Vaitomaa, J.; Rantala, A.; Halinen, K.; Rouhiainen, L.; Tallberg, P.; Mokelke, L.; Sivonen, K. Quantitative real-time PCR for determination of microcystin synthetase E copy numbers for Microcystis and Anabaena in Lakes. Appl. Environ. Microbiol. 2003, 69, 7289–7297. [Google Scholar] [CrossRef] [PubMed]
- Rinta-Kanto, J.M.; Ouellette, A.J.A.; Boyer, G.L.; Twiss, M.R.; Bridgeman, T.B.; Wilhelm, S.W. Quantification of toxic Microcystis spp. during the 2003 and 2004 blooms in western Lake Erie using quantitative real-time PCR. Environ. Sci. Technol. 2005, 39, 4198–4205. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Noda, N.; Tsuneda, S.; Saito, T.; Itayama, T.; Inamori, Y. Highly sensitive real-time PCR assay for quantification of toxic cyanobacteria based on microcystin synthetase A gene. J. Biosci. Bioeng. 2006, 102, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-G.; Joung, S.-H.; Ahn, C.-Y.; Ko, S.-R.; Boo, S.M.; Oh, H.-M. Annual variation of Microcystis genotypes and their potential toxicity in water and sediment from a eutrophic reservoir. FEMS Microbiol. Ecol. 2010, 74, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Conradie, K.R.; Barnard, S. The dynamics of toxic Microcystis strains and microcystin production in two hypertrofic South African reservoirs. Harmful Algae 2012, 20, 1–10. [Google Scholar] [CrossRef]
- Zhang, W.; Lou, I.; Ung, W.K.; Kong, Y.; Mok, K.M. Analysis of cylindrospermopsin- and microcystin-producing genotypes and cyanotoxin concentrations in the Macau storage reservoir. Hydrobiologia 2014, 741, 51–68. [Google Scholar] [CrossRef]
- Hotto, A.M.; Satchwell, M.F.; Berry, D.L.; Gobler, C.J.; Boyer, G.L. Spatial and temporal diversity of microcystins and microcystin-producing genotypes in Oneida Lake, NY. Harmful Algae 2008, 7, 671–681. [Google Scholar] [CrossRef]
- Briand, E.; Gugger, M.; François, J.-C.; Bernard, C.; Humbert, J.-F.; Quiblier, C. Temporal variations in the dynamics of potentially microcystin-producing strains in a bloom-forming Planktothrix agardhii (Cyanobacterium) population. Appl. Environ. Microbiol. 2008, 74, 3839–3848. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Ha, J.H.; Hidaka, T.; Tsuno, H. Quantification of toxic Microcystis and evaluation of its dominance ratio in blooms using real-time PCR. Environ. Sci. Technol. 2009, 43, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Liu, X.; Tan, J.; Li, D.; Yang, H. Diversity and dynamics of microcystin—Producing cyanobacteria in China’s third largest lake, Lake Taihu. Harmful Algae 2009, 8, 637–644. [Google Scholar] [CrossRef]
- Rinta-Kanto, J.M.; Konopko, E.A.; DeBruyn, J.M.; Bourbonniere, R.A.; Boyer, G.L.; Wilhelm, S.W. Lake Erie Microcystis: Relationship between microcystin production, dynamics of genotypes and environmental parameters in a large lake. Harmful Algae 2009, 8, 665–673. [Google Scholar] [CrossRef]
- Baxa, D.V.; Kurobe, T.; Ger, K.A.; Lehman, P.W.; Teh, S.J. Estimating the abundance of toxic Microcystis in the San Francisco Estuary using quantitative real-time PCR. Harmful Algae 2010, 9, 342–349. [Google Scholar] [CrossRef]
- Sabart, M.; Pobel, D.; Briand, E.; Combourieu, B.; Salençon, M.J.; Humbert, J.F.; Latour, D. Spatiotemporal variations in microcystin concentrations and in the proportions of microcystin-producing cells in several Microcystis aeruginosa populations. Appl. Environ. Microbiol. 2010, 76, 4750–4759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okello, W.; Ostermaier, V.; Portmann, C.; Gademann, K.; Kurmayer, R. Spatial isolation favours the divergence in microcystin net production by Microcystis in Ugandan freshwater lakes. Water Res. 2010, 44, 2803–2814. [Google Scholar] [CrossRef] [PubMed]
- Fortin, N.; Aranda-Rodriguez, R.; Jing, H.; Pick, F.; Bird, D.; Greer, C.W. Detection of microcystin-producing cyanobacteria in missisquoi bay, Quebec, Canada, using quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 5105–5112. [Google Scholar] [CrossRef] [PubMed]
- Joung, S.-H.; Oh, H.-M.; Ko, S.-R.; Ahn, C.-Y. Correlations between environmental factors and toxic and non-toxic Microcystis dynamics during bloom in Daechung Reservoir, Korea. Harmful Algae 2011, 10, 188–193. [Google Scholar] [CrossRef]
- Martins, A.; Moreira, C.; Vale, M.; Freitas, M.; Regueiras, A.; Antunes, A.; Vasconcelos, V. Seasonal dynamics of Microcystis spp. and their toxigenicity as assessed by qPCR in a temperate reservoir. Mar. Drugs 2011, 9, 1715–1730. [Google Scholar] [CrossRef] [PubMed]
- Te, S.H.; Gin, K.Y.-H. The dynamics of cyanobacteria and microcystin production in a tropical reservoir of Singapore. Harmful Algae 2011, 10, 319–329. [Google Scholar] [CrossRef]
- Wood, S.A.; Rueckert, A.; Hamilton, D.P.; Cary, S.C.; Dietrich, D.R. Switching toxin production on and off: intermittent microcystin synthesis in a Microcystis bloom. Environ. Microbiol. Rep. 2011, 3, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Pobel, D.; Godon, J.-J.; Humbert, J.-F.; Robin, J. High-frequency monitoring of the genetic diversity and the potential toxicity of a Microcystis aeruginosa bloom in a French shallow lake. FEMS Microbiol. Ecol. 2012, 79, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Otten, T.G.; Xu, H.; Qin, B.; Zhu, G.; Paerl, H.W. Spatiotemporal patterns and ecophysiology of toxigenic microcystis blooms in Lake Taihu, China: Implications for water quality management. Environ. Sci. Technol. 2012, 46, 3480–3488. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Choi, G.-G.; Ahn, C.-Y.; Oh, H.-M.; Ravi, A.K.; Asthana, R.K. Dynamics of microcystin production and quantification of potentially toxigenic Microcystis sp. using real-time PCR. Water Res. 2012, 46, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.H.; Jeong, D.H.; Cho, Y.C. Quantification of toxigenic Microcystis spp. in freshwaters by quantitative real-time PCR based on the microcystin synthetase A gene. J. Microbiol. 2013, 51, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Hautala, H.; Lamminmäki, U.; Spoof, L.; Nybom, S.; Meriluoto, J.; Vehniäinen, M. Quantitative PCR detection and improved sample preparation of microcystin-producing Anabaena, Microcystis and Planktothrix. Ecotoxicol. Environ. Saf. 2013, 87, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, J.S.M.; Giani, A. Estimating toxic cyanobacteria in a Brazilian reservoir by quantitative real-time PCR, based on the microcystin synthetase D gene. J. Appl. Phycol. 2013, 25, 1545–1554. [Google Scholar] [CrossRef]
- Ngwa, F.F.; Madramootoo, C.A.; Jabaji, S. Comparison of cyanobacterial microcystin synthetase (mcy) E gene transcript levels, mcy E gene copies, and biomass as indicators of microcystin risk under laboratory and field conditions. Microbiologyopen 2014, 3, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Savela, H.; Vehniainen, M.; Spoof, L.; Nybom, S.; Meriluoto, J.; Lamminmaki, U. Rapid quantification of mcyB copy numbers on dry chemistry PCR chips and predictability of microcystin concentrations in freshwater environments. Harmful Algae 2014, 39, 280–286. [Google Scholar] [CrossRef]
- Guedes, I.A.; da Costa Leite, D.M.; Manhaes, L.A.; Bisch, P.M.; Azevedo, S.M.F.O.; Pacheco, A.B.F. Fluctuations in microcystin concentrations, potentially toxic Microcystis and genotype diversity in a cyanobacterial community from a tropical reservoir. Harmful Algae 2014, 39, 303–309. [Google Scholar] [CrossRef]
- Li, D.; Gu, A.Z.; He, M. Quantification and genetic diversity of total and microcystin-producing Microcystis during blooming season in Tai and Yang-cheng lakes, China. J. Appl. Microbiol. 2014, 116, 1482–1494. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yu, Y.; Yang, Z.; Kong, F.; Zhang, T.; Tang, S. The dynamics of toxic and nontoxic Microcystis during bloom in the large shallow lake, Lake Taihu, China. Environ. Monit. Assess. 2014, 186, 3053–3062. [Google Scholar] [CrossRef] [PubMed]
- Beversdorf, L.J.; Chaston, S.D.; Miller, T.R.; McMahon, K.D. Microcystin mcyA and mcyE gene abundances are not appropriate indicators of microcystin concentrations in lakes. PLoS ONE 2015, 10, e0125353. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.A.; Rollwagen-Bollens, G.; Bollens, S.M.; Faber-Hammond, J.J. Environmental influence on cyanobacteria abundance and microcystin toxin production in a shallow temperate lake. Ecotoxicol. Environ. Saf. 2015, 114, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Rai, P.K.; Chau, R.; Ravi, A.K.; Neilan, B.A.; Asthana, R.K. Temporal variations in microcystin-producing cells and microcystin concentrations in two fresh water ponds. Water Res. 2015, 69, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Christoffersen, K.; Lyck, S.; Winding, A. Microbial activity and bacterial community structure during degradation of microcystins. Aquat. Microb. Ecol. 2002, 27, 125–136. [Google Scholar] [CrossRef]
- Rasmussen, J.P.; Giglio, S.; Monis, P.T.; Campbell, R.J.; Saint, C.P. Development and field testing of a real-time PCR assay for cylindrospermopsin-producing cyanobacteria. J. Appl. Microbiol. 2008, 104, 1503–1515. [Google Scholar] [CrossRef] [PubMed]
- Orr, P.T.; Rasmussen, J.P.; Burford, M.A.; Eaglesham, G.K.; Lennox, S.M. Evaluation of quantitative real-time PCR to characterise spatial and temporal variations in cyanobacteria, Cylindrospermopsis raciborskii (Woloszynska) Seenaya et Subba Raju and cylindrospermopsin concentrations in three subtropical Australian reservoirs. Harmful Algae 2010, 9, 243–254. [Google Scholar] [CrossRef]
- Moreira, C.; Martins, A.; Azevedo, J.; Freitas, M.; Regueiras, A.; Vale, M.; Antunes, A.; Vasconcelos, V. Application of real-time PCR in the assessment of the toxic cyanobacterium Cylindrospermopsis raciborskii abundance and toxicological potential. Appl. Microbiol. Biotechnol. 2011, 92, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Marbun, Y.; Yen, H.; Lin, T.; Lin, H.; Michinaka, A. Rapid on-site monitoring of cylindrospermopsin-producers in reservoirs using quantitative PCR. Sustain. Environ. Res. 2012, 22, 143–151. [Google Scholar]
- Campo, E.; Lezcano, M.-Á.; Agha, R.; Cirés, S.; Quesada, A.; El-Shehawy, R. First TaqMan assay to identify and quantify the cylindrospermopsin-producing cyanobacterium Aphanizomenon ovalisporum in water. Adv. Microbiol. 2013, 3, 430–437. [Google Scholar] [CrossRef]
- Burford, M.A.; Davis, T.W.; Orr, P.T.; Sinha, R.; Willis, A.; Neilan, B.A. Nutrient-related changes in the toxicity of field blooms of the cyanobacterium, Cylindrospermopsis raciborskii. FEMS Microbiol. Ecol. 2014, 89, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Al-Tebrineh, J.; Gehringer, M.M.; Akcaalan, R.; Neilan, B.A. A new quantitative PCR assay for the detection of hepatotoxigenic cyanobacteria. Toxicon 2011, 57, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Rucker, J.; Stuken, A.; Nixdorf, B.; Fastner, J.; Chorus, I.; Wiedner, C. Concentrations of particulate and dissolved cylindrospermopsin in 21 Aphanizomenon-dominated temperate lakes. Toxicon 2007, 50, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Cirés, S.; Wörmer, L.; Timón, J.; Wiedner, C.; Quesada, A. Cylindrospermopsin production and release by the potentially invasive cyanobacterium Aphanizomenon ovalisporum under temperature and light gradients. Harmful Algae 2011, 10, 668–675. [Google Scholar] [CrossRef]
- Al-Tebrineh, J.; Mihali, T.K.; Pomati, F.; Neilan, B.A. Detection of saxitoxin-producing cyanobacteria and Anabaena circinalis in environmental water blooms by quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 7836–7842. [Google Scholar] [CrossRef] [PubMed]
- Al-Tebrineh, J.; Pearson, L.A.; Yasar, S.A.; Neilan, B.A. A multiplex qPCR targeting hepato- and neurotoxigenic cyanobacteria of global significance. Harmful Algae 2012, 15, 19–25. [Google Scholar] [CrossRef]
- Te, S.H.; Chen, E.Y.; Gin, K.Y.-H. Comparison of quantitative PCR and droplet digital PCR multiplex assays for two genera of bloom-forming cyanobacteria, Cylindrospermopsis and Microcystis. Appl. Environ. Microbiol. 2015, 81, 5203–5211. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Target Genes | MC Extraction | MC Analysis | [MC] range µg·L−1 | Correlation between mcy genes and [MC]. | Correlation between Chla or No. Cells and [MC] | Study Site | Ref. | Year |
|---|---|---|---|---|---|---|---|---|
| mcyE | Total | HPLC | 0–9 | Yes | n.d | Lakes Tuusulanjarvi and Hiidenvesi, FI | [51] | 2003 |
| mcyB | Particulate | HPLC | n.a | n.d | Yes | Lake Wannsee, DE | [49] | 2003 |
| mcyD, 16S rRNA | Particulate | PPIA | 0–15.4 | n.d | n.d | Lake Erie, U.S. | [52] | 2005 |
| mcyA | Total | HPLC | 1.3–22.7 | n.d | n.d | Lakes Kasumigaura and Kitaura, JP | [53] | 2006 |
| mcyD, 16S rRNA | Particulate | PPIA | 0–2.6 | No | Yes | Lake Oneida, NY, U.S. | [57] | 2008 |
| mcyA, PG-IGS | Particulate | PPIA | 0–7.4 | No | n.d | BNV artificial lake, FR | [58] | 2008 |
| mcyD, 16S rRNA | Particulate | PPIA | 0.1–78.8 | Yes | No | Lakes Champlain, Agawam Ronkonkoma and Mill Pond, U.S. | [59] | 2009 |
| mcyA, 16S rRNA | Total | ELISA | 8–8000 | Yes | Yes | Hirosawa-No-ike fish pond, JP | [60] | 2009 |
| mcyA | Particulate | PPIA | 0–9 | No | n.d | Lake Taihu, CN | [61] | 2009 |
| mcyD, 16S rRNA | Particulate | PPIA | 0-21.7 | Yes | Yes | Lake Erie, US | [62] | 2009 |
| mcyD, 16S rRNA | Particulate | PPIA | 0–3.6 | No | n.d | San Francisco Bay, U.S. | [63] | 2010 |
| mcyJ, PG-IGS | Dissolved | HPLC | 0–0.6 | n.d | Yes | Daechung Reservoir, KR | [54] | 2010 |
| mcyB, cpcBA | Particulate | PPIA, LC-MS/MS | 1–18 | No | Yes | Loire river, FR | [64] | 2010 |
| mcyB, PC-IGS | Particulate | HPLC | 0.02–10 | n.d | Yes | Lakes Saka, George, Edward, Mburo, Murchison Bay, UG | [65] | 2010 |
| mcyD | Particulate | ELISA, HPLC | 0–4 | Yes | n.d | Lake Champlain, CA | [66] | 2010 |
| mcyJ, cpcBA | Particulate | HPLC | 0–0.6 | Yes | Yes | Daechung Reservoir, KR | [67] | 2011 |
| mcyB,A, 16S rRNA | Particulate | ELISA | 2.5–7.0 | Yes | No | Tâmega River, PT | [68] | 2011 |
| mcyE, 16S rRNA | Total | ELISA | 0-586 | Yes | Yes | Kranji Reservoir, SG | [69] | |
| mcyE | Particulate and dissolved | ELISA | 0–25 | Yes | Yes | Lake Rotorua, NZ | [70] | 2011 |
| mcyB, mcyE, 16S rRNA | Total | ELISA | 0–217 | Yes | n.d | Roodeplaat reservoir, ZA | [55] | 2012 |
| mcyB, cpcBA | Particulate | HPLC | 10–100 | No | Yes | Shallow lake, FR | [71] | 2012 |
| mcyE, cpcBA | Total | ELISA | 0.4–28.7 | Yes | Yes | Lake Taihu, CN | [72] | 2012 |
| mcyA, 16S rRNA | Particulate | LC-MS/MS | 0-528 | Yes | Yes | Durgakund Pond, Varanasi, IN | [73] | 2012 |
| mcyA, 16S rRNA | Particulate | HPLC | 1.3–48.6 | No | n.d | Daechung, Yongdam, Chungju, Soyang, Euam reservoir, KR | [74] | 2013 |
| mcyB | Particulate | HPLC, ELISA, LC-MS/MS | 0–145 * | Yes | n.d | Hauninen reservoir, FI | [75] | 2013 |
| mcyD | Particulate | ELISA | 0–0.5 | Yes | n.d | Furnas reservoir, BR | [76] | 2013 |
| mcyE, 16S rRNA | Particulate | ELISA | 0.01–24 | Yes | Yes | Missisquoi Bay, CA | [77] | 2014 |
| mcyB | Particulate | ELISA, LC-MS/MS | 0–30.4 | Yes | n.d | Aland Islands, FI | [78] | 2014 |
| mcyB, 16S rRNA | Total | LC-MS/MS | 0.02–0.5 | No | Yes | Funil reservoir, BR | [79] | 2014 |
| mcyA-E, G, J 16S rRNA | Total | LC-MS/MS | 0–0.05 | No | No | Macau storage reservoir, CN | [56] | 2014 |
| mcyA, 16S rRNA | Total | ELISA | n.a | Yes | Yes | Lakes Tai and Yang-cheng, CN | [80] | 2014 |
| mcyD, cpcBA | Particulate | HPLC | 0.2–4.2 | No | Yes | Lake Taihu, CN | [81] | 2014 |
| mcyD, 16S rRNA | Particulate and dissolved | HPLC | 1–17.6 | Yes | Yes | Lake Chaohu, CN | [9] | 2014 |
| mcyE, mcyA, 16S rRNA | Total | LC-MS/MS | 0–66 | No | n.d | Lakes Mendota, Monona, Wingra and Kegonsa, U.S. | [82] | 2015 |
| mcyA, mcyE | Particulate and dissolved | ELISA | 0–15 | Yes | Yes | Vancouver Lake, U.S. | [83] | 2015 |
| mcyB, PC-IGS, | Total | ELISA | 0.3–165 | Yes | Yes | Klamath river, U.S. | [10] | 2015 |
| mcyA | Particulate | ELISA | 0–77 | Yes | No | Lake Aydat, FR | [11] | 2015 |
| mcyA, mcyB, 16S rRNA | Particulate and dissolved | LC-MS/MS | 2.2–38.6 | Yes | Yes | Lakshmikund and Sankuldhara, IN | [84] | 2015 |
| Target Genes | CYL Extraction | CYL Analysis | [CYL] range µg·L−1 | Correlation between cyr genes and [CYL] | Study Site | Ref. | Year |
|---|---|---|---|---|---|---|---|
| cyrC, rpoC1 | Particulate | LC-MS or MALDI-TOF/MS | n.a | No | Lakes South Australia and Imperial, AU | [86] | 2008 |
| cyrC, rpoC1, 16S rRNA | Particulate and dissolved | LC–MS/MS | n.a | Yes | Lakes Samsonvale, Somerset and Wivenhoe, AU | [87] | 2010 |
| cyrC, 16S rRNA, rpoC1 | Particulate and dissolved | HPLC | 0–0.3 | n.d | Lake Vela, PT | [88] | 2011 |
| cyrC, rpoC1 | Total | ELISA | 0–0.7 | n.d | Lake Cheng Kung, TW | [89] | 2012 |
| cyrA, 16S rRNA | Total | ELISA | 0.2–0.6 | Yes | Murray river, AU | [8] | 2012 |
| cyrJ, rpoC1 | Dissolved | LC-MS/MS | 0.1–0.7 | Yes | Alange reservoir, ES | [90] | 2013 |
| cyrA, 16S rRNA | Particulate | LC-MS/MS | n.a | - | North Pine reservoir, AU | [91] | 2014 |
| cyrC, rpcC1 | - | LC-MS/MS | 0–1.3 | Yes | Macau storage reservoir, MO | [56] | 2014 |
| Target Genes | STX Extraction | STX Analysis | [STX] range µg·L−1 | Correlation between sxt genes and [STX] | Study Site | Ref. | Year |
|---|---|---|---|---|---|---|---|
| sxtA, 16S rRNA | Particulate | HPLC | 0.08–14.5 | Yes | Australian water bodies | [95] | 2010 |
| sxtA, 16S rRNA | Total | ELISA | 0.015–0.023 | n.d | Murray River, AU | [8] | 2012 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacheco, A.B.F.; Guedes, I.A.; Azevedo, S.M.F.O. Is qPCR a Reliable Indicator of Cyanotoxin Risk in Freshwater? Toxins 2016, 8, 172. https://doi.org/10.3390/toxins8060172
Pacheco ABF, Guedes IA, Azevedo SMFO. Is qPCR a Reliable Indicator of Cyanotoxin Risk in Freshwater? Toxins. 2016; 8(6):172. https://doi.org/10.3390/toxins8060172
Chicago/Turabian StylePacheco, Ana Beatriz F., Iame A. Guedes, and Sandra M.F.O. Azevedo. 2016. "Is qPCR a Reliable Indicator of Cyanotoxin Risk in Freshwater?" Toxins 8, no. 6: 172. https://doi.org/10.3390/toxins8060172